壮药龙盘止咳方治疗小鼠甲型流感病毒H1N1肺炎的作用及机制*
2022-03-28邹敏谢彩英甘娜农志飞李崇进韦杏
邹敏, 谢彩英, 甘娜, 农志飞, 李崇进, 韦杏△
壮药龙盘止咳方治疗小鼠甲型流感病毒H1N1肺炎的作用及机制*
邹敏1, 谢彩英2, 甘娜1, 农志飞3, 李崇进4, 韦杏2△
(1广西国际壮医医院儿科,广西 南宁 530001;2广西中医药大学壮医药学院,广西 南宁 530200;3广西医科大学第一附属医院儿科,广西 南宁 530021;4茂名市中医院儿科,广东 茂名 525099)
探究壮药龙盘止咳方的抗流感病毒性肺炎作用及其相关机制。BALB/c小鼠随机分为对照组、模型组、高剂量(10.4 g/kg)龙盘止咳方组、低剂量(5.2 g/kg)龙盘止咳方组和龙盘止咳方(10.4 g/kg)+Toll样受体3(TLR3)激动剂聚肌胞苷酸[Poly(I:C),20 mg/kg]组,每组20只。采用甲型流感病毒(IAV)H1N1滴鼻法建立病毒性肺炎模型,观察小鼠一般状态,并每组取10只小鼠,记录15 d的存活率和存活时间;计算肺、脾和胸腺指数;HE染色观察肺组织病理变化;实时荧光定量PCR检测肺病毒载量;ELISA法检测肺匀浆中白细胞介素6(IL-6)、IL-4、干扰素α(IFN-α)、IFN-β和IFN-γ水平;Western blot检测肺组织TLR3/视黄酸诱导基因I(RIG-I)/核因子κB(NF-κB)通路相关蛋白的表达。对照组小鼠精神状态良好,在实验期间未出现死亡;与对照组相比,模型组小鼠在感染后出现典型的流感症状,小鼠存活率、存活时间、脾指数和胸腺指数显著降低(<0.05),肺指数,肺组织病理学评分,病毒载量,肺匀浆中IL-6、IL-4、IFN-γ、IFN-α和IFN-β水平,肺匀浆中IFN-γ/IL-4比值,以及肺组织TLR3、胞核NF-κB p65、RIG-I和干扰素β启动子刺激蛋白1(IPS-1)表达均显著增加(<0.05);与模型组相比,高、低剂量龙盘止咳方组小鼠症状明显减轻,小鼠存活率、存活时间、脾指数、胸腺指数、肺匀浆中IFN-α和IFN-β水平及肺组织RIG-I和IPS-1表达均显著增加(<0.05),肺指数,肺组织病理学评分,病毒载量,肺匀浆中IL-6、IL-4和IFN-γ水平,肺匀浆中IFN-γ/IL-4比值,以及TLR3和胞核NF-κB p65表达均显著降低(<0.05);而Poly(I:C)可明显减弱龙盘止咳方对IAV H1N1感染小鼠的肺脏保护作用。壮药龙盘止咳方有减轻IAV H1N1感染小鼠肺损伤的作用,其机制可能与调节TLR3/RIG-I/NF-κB信号通路,抑制过度的先天炎症反应有关。
壮药龙盘止咳方;甲型流感病毒H1N1;肺炎;Toll样受体3;视黄酸诱导基因I;核因子κB
流感是由流感病毒引起并在人与人之间传播的急性病毒性呼吸道感染。甲型流感病毒(influenza A virus, IAV)感染是其主要致病因素,可扩散到肺部并在肺部大量复制,引起病毒性肺炎,严重威胁人类健康[1-2]。疫苗提供的免疫效果有限,且需要每年重新接种。目前除接种疫苗预防外,确诊后治疗以抗病毒药物为主,但长期大量使用抗病毒药可增加病毒的抗药性,具有较大毒副作用[3]。因此,对高效、低毒药物的开发一直是国内外学者关注的重点。
Toll样受体(Toll-like receptor, TLR)是识别细菌、病毒和真菌中病原体相关分子模式的主要受体,其中TLR3识别病毒复制过程中产生的双链RNA(double-stranded RNA, dsRNA),并且会诱导核因子κB(nuclear factor-κB, NF-κB)的激活,触发炎症细胞因子[白细胞介素6(interleukin, IL-6)、肿瘤坏死因子α(tumor necrosis factor-α, TNF-α)和干扰素γ(interferon-γ, IFN-γ)]产生[4-5]。视黄酸诱导基因I(retinoic acid-inducible gene I, RIG-I)是病毒诱导的抗病毒干扰素(interferon, IFN)和参与清除病毒感染的促炎细胞因子的重要调节剂,是抗病毒免疫的关键介质[6],可被病毒感染细胞中的病毒dsRNA激活,与线粒体抗病毒信号蛋白干扰素β启动子刺激蛋白1(interferon-β promoter stimulator 1, IPS-1)相互作用,诱导下游信号传导,并激活I型和III型IFN的产生[7]。通过RIG-I途径感知病毒存在对于宿主成功防御RNA病毒感染至关重要。据报道,RIG-I过表达增加了IAV感染小鼠的存活率[8],表明TLR3/RIG-I/NF-κB通路为IAV感染治疗的潜在靶点。
近年来,中医药在流感病毒性肺炎的防治上具有其独特的优势和发展前景,尤其体现在免疫功能的调节上[9-10]。壮药龙盘止咳方(Longpan-Zhike formula, LPZKF)具有清宣肺气、化痰止咳、补虚扶正的功效,现代研究发现其具有抗炎和免疫调节作用,临床上治疗小儿咳嗽(风热犯肺证)疗效显著,且安全性好[11-12]。然而,LPZKF的抗流感作用却尚未被报道。因此,本研究以IAV亚型H1N1肺炎模型小鼠为对象,旨在探究LPZKF的抗流感病毒性肺炎作用及其相关机制。
材料和方法
1 材料
1.1实验动物、病毒和细胞SPF级BALB/c小鼠100只,6~8周龄,雌雄各半,体重(20±2) g,购自北京维通利华实验动物技术有限公司,许可证为SCXK(京)2019-0009。小鼠适应流感病毒A/FM1/47(H1N1, FM1)来自中国疾病预防控制中心,首先将该病毒在Madin-Darby犬肾(Madin-Darby canine kidney, MDCK)细胞(武汉普诺赛生命科技有限公司,CL-0154)中进行噬斑纯化,并在9日龄的鸡胚胎中进行复制。在研究之前,在小鼠中预先滴定病毒库以确定合适的攻击剂量。两次常规鸡胚复苏后,血凝滴度为1∶40。通过双重稀释法测定了不同浓度的病毒感染后14 d小鼠的死亡率。使用引起20%小鼠死亡的病毒浓度(血凝滴度1∶640),并给予每只小鼠50 μL病毒溶液进行实验。
1.2药品及试剂LPZKF含有龙脷叶10 g、鱼腥草10 g、柿叶5 g、不出林5 g、盘龙参5 g和甘草5 g。将上述药材浸入8倍量的水中充分浸泡30 min后,进行煎煮40 min,然后过滤,药渣再加入6倍量水继续煎煮40 min后,再次过滤,合并两次过滤药液,煎煮浓缩至100 mL,1 mL相当于生药0.4 g。TLR3激动剂聚肌胞苷酸[polyinosinic:polycytidylic acid, Poly(I:C)]购自上海懋康生物科技有限公司;HE染色试剂、RIPA裂解液和BCA试剂盒购自碧云天生物科技公司;小鼠IL-4、IL-6、IFN-γ、IFN-α和IFN-β ELISA检测试剂盒购自上海酶联生物技术有限公司;TRIzol、PrimeScript™ RT试剂盒和SYBR®Premix Ex Taq™ II试剂盒购自Takara;实时荧光定量PCR引物购自上海GenePharma公司;兔抗小鼠TLR3、NF-κB p65、GAPDH和histone H3抗体及HRP标记的山羊抗兔IgG Ⅱ抗购自Abcam;抗RIG-I和IPS-1抗体购自Santa Cruz。
1.3仪器iMark™多功能酶标仪(Bio-Rad);Prism®7300型荧光定量PCR系统(Applied Biosystems);BX61电动显微镜(Olympus)。
2 主要方法
2.1分组、造模及干预将小鼠随机分为5组:对照(control)组、模型(model)组、高剂量(10.4 g/kg)LPZKF(high-dose LPZKF, LPZKF-H)组、低剂量(5.2 g/kg)LPZKF(low-dose LPZKF, LPZKF-L)组和LPZKF+Poly(I:C)[10.4 g/kg LPZKF+20 mg/kg Poly(I:C)]组,每组20只。适应1周后,除control组外,其余各组小鼠均用乙醚轻度麻醉,用50 μL IAV H1N1悬浮液(稀释1∶640)经鼻攻击建立流感病毒性肺炎模型[13];control组在用乙醚轻度麻醉后,鼻腔滴入50 μL 0.9% NaCl无菌溶液。受感染的小鼠均未死亡。根据人与小鼠的体重差异计算药物剂量,小鼠剂量相当于人类(按平均体质量70 kg体表面积换算)临床剂量的9.1倍,LPZKF成人每日给药剂量相当于生药40 g,则小鼠每日给药剂量为5.2 g/kg,因此设置低、高剂量药物剂量为5.2和10.4 g/kg。感染后24 h,给予感染小鼠上述剂量的LPZKF进行灌胃治疗,治疗组每天灌胃一次,共5 d;LPZKF+Poly(I:C)组小鼠在给予10.4 g/kg LPZKF灌胃的同时腹腔注射20 mg/kg Poly(I:C)[14]。control组和model组在相同时间点给予相同体积的生理盐水,持续给药直至实验结束。
2.2小鼠一般状态、存活率、肺和免疫相关器官指数的变化每天观察小鼠的症状、饮水和食物摄入量、毛发颜色、活动情况;每组随机选取10只小鼠,记录连续15 d的存活率和存活时间。每组剩余10只小鼠,于感染后5 d末次给药2 h后处死以收集肺、脾和胸腺组织,PBS洗涤后,用滤纸干燥并称重。肺、脾或胸腺指数=肺、脾或胸腺的重量(mg)/体重(g)。
2.3肺组织病理变化观察取部分新鲜肺组织经4%多聚甲醛固定,脱水,石蜡包埋,5 µm连续切片。采用苏木精-伊红(HE)染色,光镜下观察肺组织病理变化。通过使用先前描述的评分系统[15]对H1N1引起的肺组织病理学变化进行评分来评估肺损伤的严重程度。0到4分分别代表正常、轻度、重度和非常重度肺损伤。具体而言,正常肺为0分,轻度间质性肺炎(低于25%)为1分,中度间质性肺炎(25~50%)为2分,严重间质性肺炎(50~75%)为3分,非常严重的间质性肺炎(肺受累高于75%)为4分。
2.4实时荧光定量PCR检测肺组织病毒载量取部分肺组织,研磨后在4 ℃下以4 000×离心15 min取上清液为肺匀浆。使用TRIzol提取其中的总RNA,并按照制造商的说明将其逆转录成cDNA,然后进行PCR扩增(参数:95 ℃ 2 min;95 ℃ 15 s,62 ℃ 30 s,72 ℃ 30 s,40个循环)。每个样品的分析一式三份进行。PCR产物的IAV量以GAPDH作为内参照,采用2-ΔΔCt法计算。IAV M基因的正向引物序列为5'-AATGGTGCAGGCGATGAGAG-3',反向引物序列为5'-TACTTGCGGCAACAACGAGAG-3';GAPDH的正向引物序列为5'-CCTCGTCCCGTAGACAAAATG-3',反向引物序列为5'-TGAGGTCAATGAAGGGGTCG-3'。
2.5ELISA检测肺匀浆中细胞因子水平取2.4中制备的肺匀浆上清液,根据制造商的说明使用ELISA试剂盒检测小鼠肺匀浆中IL-6、IL-4、IFN-α、IFN-β和IFN-γ水平。
2.6Western blot检测肺组织TLR3/RIG-I/NF-κB通路相关蛋白表达取小鼠部分肺组织加入RIPA裂解液研磨后,离心,取上清液为总蛋白溶液,并采用细胞核蛋白提取试剂盒提取细胞核中的蛋白质。用BCA法测量蛋白浓度后取等量(25 μg)蛋白质通过10% SDS-PAGE分离并转移到PVDF膜上,在室温下用5%脱脂牛奶封闭1 h。然后将膜与Ⅰ抗(RIG-I和TLR3,1∶1 000;IPS-1和NF-κB p65,1∶500;GAPDH和histone H3,1∶2 000)在4°C下孵育过夜。在室温下与HRP标记的羊抗兔IgG Ⅱ抗(1∶5 000)孵育2 h后,增强型化学发光试剂盒进行显色,使用Image J软件测量每个条带的灰度值。以GAPDH或Histone H3为内参蛋白,计算各目的蛋白的相对表达量。
3 统计学分析
采用SPSS 22.0和GraphPad Prism 8.0软件进行统计分析。数据均符合正态分布,以均数±标准差(mean±SD)表示。多组间比较采用单因素方差分析(One-way ANOVA),组间两两比较采用SNK-检验。以<0.05为差异有统计学意义。
结果
1 LPZKF对小鼠一般状态、存活率的影响
control组小鼠精神状态良好,毛色有光泽,行动敏捷,呼吸顺畅,体重自然增长;model组小鼠在病毒感染后2 d出现典型的流感症状,包括毛皮竖起、身体蜷曲、弓背、食欲不振、饮水减少、抽搐和呼吸急促,同时体重逐渐下降;与model组相比,LPZKF-H和LPZKF-L组小鼠头发耸立、呼吸急促、抽搐等症状明显减轻,体重增加;LPZKF+Poly(I:C)组小鼠上述症状仅表现出轻微改善,见图1A。
control组小鼠在实验期间未出现死亡;病毒感染小鼠在感染后5 d开始死亡,8 d达到高峰;model组病毒感染小鼠均在15 d内死亡,平均存活时间为7 d。与control组相比,model组小鼠存活率和存活时间均显著降低(<0.05);与model组相比,LPZKF-H和LPZKF-L组小鼠存活率和存活时间均显著增加(<0.05);与LPZKF-H组相比,LPZKF+Poly(I:C)组小鼠存活率和存活时间均显著降低(<0.05),见图1B、C。

Figure 1.Changes of body weight, and survival rate and survival time within 15 d of the mice in each group. A: body weight (n=20); B: survival rate (n=10); C: survival time (n=10). Mean±SD. *P<0.05 vs control group; #P<0.05 vs model group; △P<0.05 vs LPZKF-H group.
2 LPZKF对小鼠肺和免疫相关器官指数的影响
与control组相比,model组小鼠肺指数显著增加,脾指数和胸腺指数均显著降低(<0.05);与model组相比,LPZKF-H和LPZKF-L组小鼠肺指数显著降低,脾指数和胸腺指数均显著增加(<0.05);与LPZKF-H组相比,LPZKF+Poly(I:C)组小鼠肺指数显著增加,脾指数和胸腺指数均显著降低(<0.05),见图2。

Figure 2.Comparison of lung, spleen and thymus indexes of the mice in each group. Mean±SD. n=10. *P<0.05 vs control group; #P<0.05 vs model group; △P<0.05 vs LPZKF-H group.
3 LPZKF对小鼠肺组织病理学变化的影响
HE染色结果显示,control组小鼠肺组织肺泡结构清晰完整,肺泡壁薄,肺泡腔内无炎性分泌物,无炎性细胞浸润;与control组相比,model组小鼠肺组织出现炎性损伤,包括肺泡腔内大量炎性细胞浸润、肺泡间隔增厚及严重的间质水肿、血管充血、炎性渗出导致的支气管阻塞,肺组织病理学评分显著增加(<0.05);与model组相比,LPZKF-H和LPZKF-L组小鼠肺组织病理变化得到了改善,肺泡壁薄,毛细血管扩张,肺泡壁充血少,肺泡间隔无明显增厚,仅有少量单核细胞和淋巴细胞,细支气管无炎性渗出,肺组织病理学评分显著降低(<0.05);与LPZKF-H组相比,LPZKF+Poly(I:C)组小鼠肺组织病理学评分显著增加(<0.05),见图3。

Figure 3.Pathological changes of mouse lung tissues (HE staining, scale bar=100 μm) and the lung histopathological score. Mean±SD. n=10. *P<0.05 vs control group; *P<0.05 vs model group; △P<0.05 vs LPZKF-H group.
4 LPZKF对小鼠肺组织病毒载量的影响
与control组相比,model组小鼠肺组织中病毒载量显著增加(<0.05);与model组相比,LPZKF-H和LPZKF-L组小鼠肺组织病毒载量显著降低(<0.05);与LPZKF-H组相比,LPZKF+Poly(I:C)组小鼠肺组织病毒载量显著增加(<0.05),见图4。
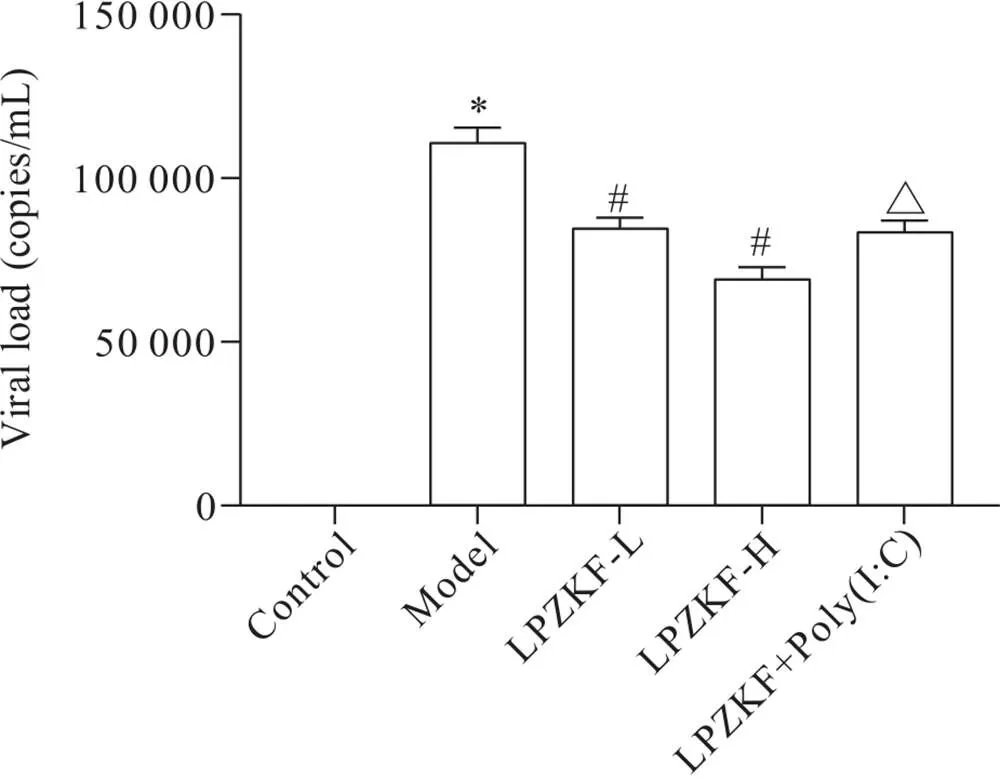
Figure 4.Comparison of viral load in lung tissues of the mice in each group. Mean±SD. n=10. *P<0.05 vs control group; #P<0.05 vs model group; △P<0.05 vs LPZKF-H group.
5 LPZKF对小鼠肺组织炎性因子的影响
与control组相比,model组小鼠肺组织中IL-6、IL-4、IFN-γ、IFN-α和IFN-β水平,以及IFN-γ/IL-4比值均显著升高(<0.05);与model组相比,LPZKF-H和LPZKF-L组小鼠肺组织IL-6、IL-4和IFN-γ水平及IFN-γ/IL-4比值均显著降低,而IFN-α和IFN-β水平均显著升高(<0.05);与LPZKF-H组相比,LPZKF+Poly(I:C)组小鼠肺组织IL-6、IL-4和IFN-γ水平及IFN-γ/IL-4比值均显著升高,而IFN-α和IFN-β水平均显著降低(<0.05),见图5。

Figure 5.Comparison of the levels of IL-6, IL-4, IFN-γ, IFN-α and IFN-β in the lung tissues of the mice in each group. Mean±SD. n=10. *P<0.05 vs control group; #P<0.05 vs model group; △P<0.05 vs LPZKF-H group.
6 LPZKF对小鼠肺组织TLR3/RIG-I/NF-κB通路相关蛋白表达的影响
与control组相比,model组小鼠肺组织中TLR3、胞核NF-κB p65、RIG-I和IPS-1表达均显著增加(<0.05);与model组相比,LPZKF-H和LPZKF-L组小鼠肺组织TLR3和胞核NF-κB p65表达均显著减少,RIG-I和IPS-1表达均显著增加(<0.05);与LPZKF-H组相比,LPZKF+Poly(I:C)组小鼠肺组织TLR3和胞核NF-κB p65表达均显著增加,RIG-I和IPS-1表达均显著减少(<0.05),见图6。
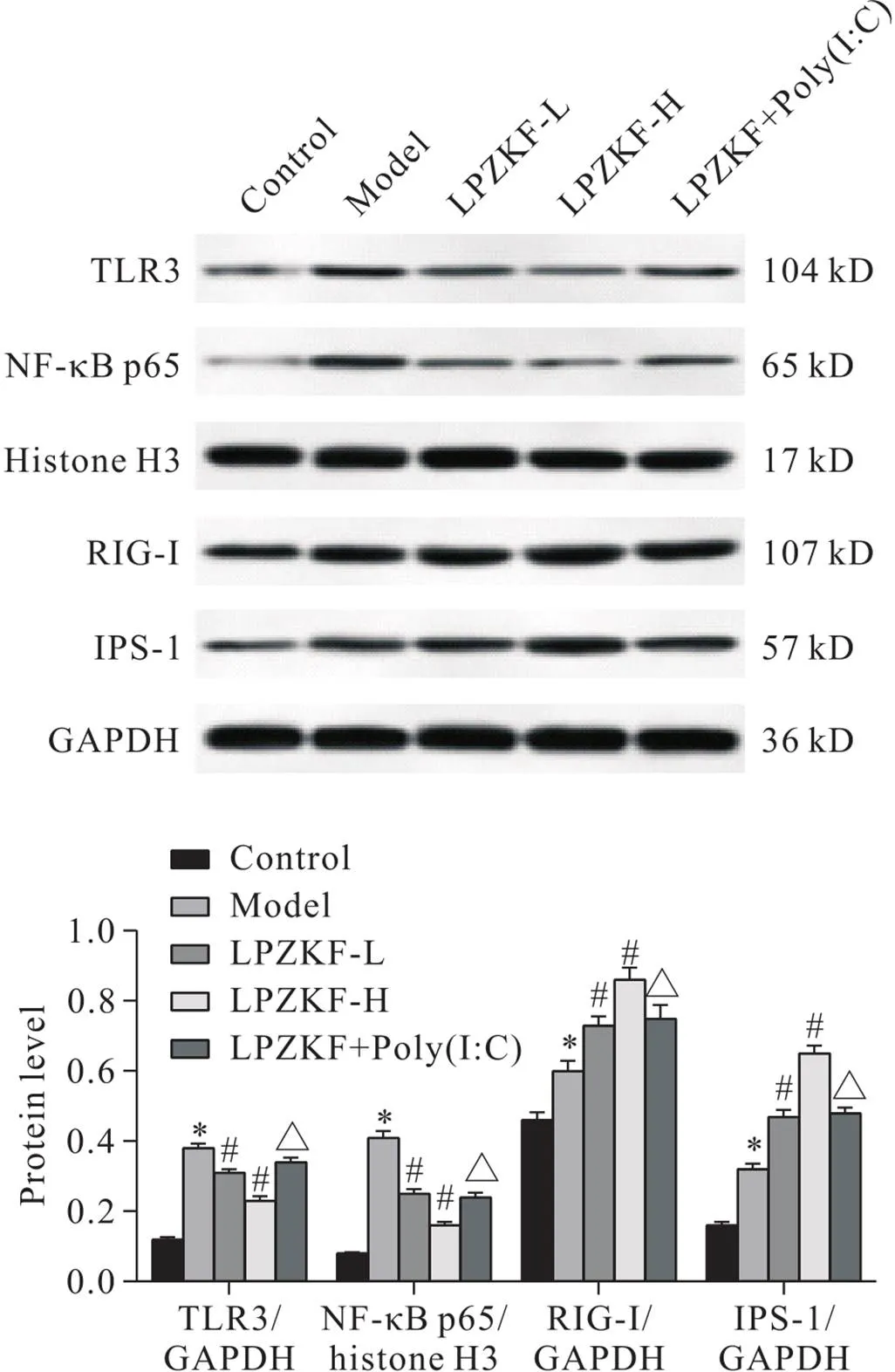
Figure 6.The expression of TLR3/RIG-I/NF-κB signaling pathway-related proteins in mouse lung tissues. Mean±SD. n=10. *P<0.05 vs control group; #P<0.05 vs model group; △P<0.05 vs LPZKF-H group.
讨论
病毒性肺炎是由具有不同传染性的病毒引起的一种急性呼吸道感染。1918年大流感、2003年严重急性呼吸系统综合征(SARS)冠状病毒、2009年甲型H1N1流感病毒及2019年的严重急性呼吸综合征冠状病毒2(SARS-CoV-2)等暴发和(或)大流行对人类生活、社会行为和经济发展产生重大影响,且目前尚未开发出针对这些病毒的特定药物。与疫苗和化学药物不同,中医药在治疗病毒性肺炎方面有着悠久的历史,具有副作用少、治疗方法多样、药源丰富、耐药性低等诸多优点[10, 16]。
中医虽无“病毒性肺炎”之称,但主要归咎于“外感病”或“外证”。中医通常根据其临床表现将其归类为“咳嗽”或“肺胀”。此外,传染性强、病死率高的病毒性肺炎在中医中通常被归类为“疫病”。中医治疗“疫病”在中国历史悠久,临床经验丰富,疗效显著。中药在治疗病毒性肺炎方面具有多种药理作用。除了直接或间接的抗病毒作用外,中医最大的优点是调节免疫功能和低副作用[17]。LPZKF中龙脷叶,有润肺止咳的功效,并有显著的抗炎活性,可减轻哮喘模型大鼠肺组织与支气管炎性病变[18];鱼腥草,为肺病圣药,具有广谱抗流感病毒和促进免疫功能的作用,治疗小儿呼吸道合胞病毒肺炎能够显著改善患儿临床症状、调节患儿免疫应答[19],与龙脷叶同为主药;柿叶,肃肺止咳,且其有效成分(如黄酮类、多糖类、萜类和有机酸类)可调节免疫[20];不出林具理气化痰止咳之功,盘龙参强于润肺、止咳、滋阴补虚;甘草,补中益气、润肺止咳,调和诸药,且甘草及甘草酸类成分有抗冠状病毒、流感病毒、呼吸道合胞病毒和人巨细胞病毒等呼吸道病毒的作用,可调节免疫,改善肺功能[21-22]。由此我们猜想LPZKF可能对IAV感染损伤的肺脏具有保护作用。本研究结果也显示,LPZKF可显著缓解感染小鼠的临床症状,减少体重减轻程度,延长存活时间,减轻肺部病变。这些结果表明LPZKF可以有效地保护小鼠免受IAV感染。
流感病毒诱导炎症细胞浸润肺组织并释放促炎细胞因子(如IL-6、TNF-α和IFN-γ),导致严重的继发性肺炎,诱导死亡。肺指数可反映肺部感染的严重程度;脾脏和胸腺是免疫相关器官,脾脏和胸腺指数反映了小鼠的免疫功能。在本研究中,LPZKF处理的小鼠肺指数和肺组织病理学评分较低,表明LPZKF可能降低肺水肿的发生率,缓解IAV引起的肺部炎症。此外,本研究数据显示IAV会显著降低小鼠胸腺和脾脏指数,提高肺组织中的IL-6、IL-4和IFN-γ水平及IFN-γ/IL-4比值。IFN-γ主要由Th1细胞分泌,IL-4主要由Th2细胞分泌,机体正常时,Th1和Th2细胞功能处于动态平衡状态,当机体受到病毒等微生物感染时,机体Th1和Th2细胞平衡失调[23]。流感病毒是一种严格的胞内病原微生物,感染宿主后主要刺激Th细胞向Th1细胞分化并释放其标志性细胞因子(IFN-γ、TNF-α、IL-2等)[24]。而LPZKF可以抑制促炎细胞因子的产生,抑制组织和细胞中的炎症反应,并且LPZKF可以增加胸腺和脾脏指数,使IAV感染小鼠中增高的IFN-γ水平及IFN-γ/IL-4比值降低,逆转Thl/Th2的失衡;提示,LPZKF可抑制过度的先天炎症反应,减少机体的免疫炎症损伤,从而保护感染IAV的小鼠免受进一步损害。
本研究还发现,LPZKF可显著增加IAV感染小鼠肺组织中IFN-α和IFN-β水平;IFN-α和IFN-β是主要的I型IFN,可抑制H1N1病毒的复制[25]。I型IFN的快速产生和最近发现的III型IFN构成了宿主抵御呼吸道病毒感染的主要防御机制,是抗病毒反应的核心和重要组成部分。TLR3和RIG-I均属于固有免疫的模式识别受体,可识别病毒增殖过程中产生的dsRNA从而激活TLR3信号通路和RIG-I信号通路,最终促进细胞因子产生,从而发挥抗病毒效应。TLR3是TLR家族中识别新型H1N1病毒启动先天免疫反应和适应性免疫反应的主要成员,介导I型IFN、促炎细胞因子和趋化因子的转录诱导;据报道,2009年H1N1流感感染患者中TLR3表达升高[26];IAV感染后TLR3信号通路优先被激活,诱导NF-κB的核转录,以释放一系列促炎细胞因子,引发严重的炎症反应[27]。RIG-I是一种高度诱导型细胞质RNA解旋酶,可感知病毒感染并与抗病毒信号蛋白IPS-1相互作用,激活对IAV的抗病毒反应,诱导I型和III型IFN产生,这可能会限制病毒复制并增加对感染的抵抗力[28];并且IFN-α和IFN-β水平可以间接反映RIG-I的活化情况。本研究结果显示,模型组肺组织病毒载量和RIG-I、胞核NF-κB p65、TLR3、IPS-1蛋白表达均显著升高,说明TLR3/RIG-I/NF-κB信号通路被激活;model组小鼠肺组织RIG-I、IPS-1表达增加,推测其原因可能是机体在面对感染时所作出的一种代偿性的增加,但这种代偿性的增加不足以抵抗病毒感染。小鼠口服LPZKF后,病毒复制和胞核NF-κB p65、TLR3蛋白过表达受到抑制,RIG-I、IPS-1蛋白表达增加;且在LPZKF干预的基础上,使用TLR3激动剂Poly(I:C)上调TLR3的表达后,发现LPZKF对IAV感染小鼠肺损伤的保护作用被明显减弱,且RIG-I、IPS-1表达降低。推测原因可能是Poly(I:C)干预后,TLR3被激活,诱导NF-κB活化并易位至细胞核,促进一系列促炎细胞因子释放,加重炎症反应,这可能使机体对感染的抵抗力减弱,进而导致RIG-I、IPS-1表达降低。这些结果表明LPZKF可能通过调节TLR3/RIG-I/NF-κB信号通路来抑制过度的炎症反应,进而减轻IAV感染小鼠肺损伤。
综上所述,壮药LPZKF可能通过调节TLR3/RIG-I/NF-κB信号通路,抑制过度的先天炎症反应,从而减轻IAV感染小鼠的肺损伤。本研究仅初步探讨了LPZKF对IAV感染小鼠肺损伤的保护作用,在未来的工作中,我们将进一步研究其他潜在的活性成分是否具有直接的抗病毒或抗炎作用;此外,在后续的研究中将考虑进行基因敲除或沉默的细胞实验,深入分析其作用机制是否与TLR3/RIG-I/NF-κB通路有关。
[1] Roychoudhury S, Das A, Sengupta P, et al. Viral pandemics of the last four decades: pathophysiology, health impacts and perspectives[J]. Int J Environ Res Public Health, 2020, 17(24):9411-9449.
[2]石云锋, 师小函, 朱军, 等. NLRP3炎症小体在小鼠甲型流感病毒感染继发MRSA肺炎中的表达变化[J]. 中国病理生理杂志, 2020, 36(9):1673-1679.
Shi YF, Shi XH, Zhu J, et al. Expression changes of NLRP3 inflammasome in MRSA pneumonia secondary to influenza A virus infection in mice[J]. Chin J Pathophysiol, 2020, 36(9):1673-1679.
[3] Ye Y, Wang H, Liu J, et al. Polygalasaponin F treats mice with pneumonia induced by influenza virus[J]. Inflammopharmacology, 2020, 28(1):299-310.
[4] Alexopoulou L, Holt AC, Medzhitov R, et al. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3[J]. Nature, 2001, 413(6857):732-738.
[5] Zhang L, Zhang B, Wang L, et al. Huanglian-Houpo drug combination ameliorates H1N1-induced mouse pneumonia via cytokines, antioxidant factors and TLR/MyD88/NF-κB signaling pathways[J]. Exp Ther Med, 2021, 21(5):428.
[6] Rehwinkel J, Tan CP, Goubau D, et al. RIG-I detects viral genomic RNA during negative-strand RNA virus infection[J]. Cell, 2010, 140(3):397-408.
[7] Onomoto K, Onoguchi K, Yoneyama M. Regulation of RIG-I-like receptor-mediated signaling: interaction between host and viral factors[J]. Cell Mol Immunol, 2021, 18(3):539-555.
[8] Wang X, Wu W, Zhang W, et al. RIG-I overexpression decreases mortality of cigarette smoke exposed mice during influenza A virus infection[J]. Respir Res, 2017, 18(1):166-178.
[9]周鹏, 刘洋, 吕伟, 等. 化湿解毒方对重症流感病毒性肺炎“炎症风暴”的免疫调节作用[J]. 中国中医急症, 2021, 30(4):685-688.
Zhou P, Liu Y, Lv W, et al. Immunomodulatory effect of Huashi Jiedu Recipe on "inflammatory storm" of severe influenza virus pneumonia[J]. J Emerg Syndr Tradit Chin Med, 2021, 30(4):685-688.
[10] 李玲, 吴佳敏, 欧阳建军, 等. 抗流感病毒性肺炎的有效中药复方筛选及机制研究[J]. 中国免疫学杂志, 2018, 34(8):1168-1173.
Li L, Wu JM, Ouyang JJ, et al. Screening and mechanism of effective traditional Chinese medicine compounds against influenza virus pneumonia[J]. Chin J Immunol, 2018, 34(8):1168-1173.
[11] 韦杏, 冯智琼, 邹敏, 等. 龙盘止咳方治疗小儿咳嗽证效研究[J]. 吉林中医药, 2021, 41(7):895-898.
Wei X, Feng ZQ, Zou M, et al. Study on the syndrome effect of Longpan Zhike recipe in the treatment of children's cough[J]. J Tradit Chin Med Chin Mater Med Jilin, 2021, 41(7):895-898.
[12] 韦杏, 邹敏, 冯志琼, 等. 壮医龙盘止咳方治疗小儿急性支气管炎风热犯肺证30例临床观察[J]. 湖南中医杂志, 2020, 36(3):1-3.
Wei X, Zou M, Feng ZQ, et al. Clinical observation on treatment of 30 cases of acute bronchitis with wind-heat invasion of lung syndrome by zhuang medicine longpan cough decoction[J]. Hunan J Tradit Chin Med, 2020, 36(3):1-3.
[13] Fu YJ, Yan YQ, Qin HQ, et al. Effects of different principles of Traditional Chinese Medicine treatment on TLR7/NF-κB signaling pathway in influenza virus infected mice[J]. Chin Med, 2018, 13:42.
[14] Garcia-Valtanen P, Guzman-Genuino RM, Hayball JD, et al. Polyinosinic: polycytidylic acid and murine cytomegalovirus modulate expression of murine IL-10 and IL-21 in white adipose tissue[J]. Viruses, 2020, 12(5):569-584.
[15] Liu W, Ren X, Wang Q, et al. Pharmacological inhibition of poly (ADP-ribose) polymerase by olaparib ameliorates influenza-virus-induced pneumonia in mice[J]. Eur J Clin Microbiol Infect Dis, 2021, 40(1):159-167.
[16] Xi S, Li Y, Yue L, et al. Role of traditional Chinese medicine in the management of viral pneumonia[J]. Front Pharmacol, 2020, 11:582322.
[17] Ma HD, Deng YR, Tian Z, et al. Traditional chinese medicine and immune regulation[J]. Clin Rev Allergy Immunol, 2013, 44(3):229-241.
[18] 陈素雯, 魏嘉宝, 邱思娃, 等. 龙脷叶水提取物对哮喘大鼠肺组织病理学及炎症因子的影响[J]. 广东药科大学学报, 2021, 37(1):55-60.
Chen SW, Wei JB, Qiu SW, et al. The effect of water extract of Longyiye on lung histopathology and inflammatory factors in asthmatic rats[J]. J Guangdong Pharm Univ, 2021, 37(1):55-60.
[19] 郭艳辉, 张琴, 王洪声. 鱼腥草雾化辅助治疗小儿呼吸道合胞病毒肺炎的效果研究[J]. 包头医学院学报, 2020, 36(12):15-18.
Guo YH, Zhang Q, Wang HS. Effect of atomization of Houttuynia cordata on respiratory syncytial virus pneumonia in children [J]. J Baotou Med Coll, 2020, 36(12):15-18
[20]诸葛芳, 韦金丽, 余晖, 等. 柿叶有效成分抗肿瘤作用及其作用机制的研究进展[J]. 中华中医药学刊, 2021, 39(6):125-128.
Zhu GF, Wei JL, Yu H, et al. Research progress on antitumor effect and mechanism of active components of persimmon leaves [J]. Chin J Tradit Chin Med, 2021, 39(6):125-128
[21] 王钧楠, 周永峰, 崔园园, 等. 甘草增强免疫的物质基础及其作用机制研究进展[J]. 中草药, 2021, 52(6):1844-1850.
Wang JN, Zhou YF, Cui YY, et al. Research progress on the material basis and mechanism of immune enhancement of licorice[J]. Chin Tradit Herb Drugs, 2021, 52(6):1844-1850.
[22] 张明发, 沈雅琴. 甘草及甘草酸类成分抗病毒性肺炎的药理作用研究进展[J]. 药物评价研究, 2020, 43(7):1452-1468.
Zhang MF, Shen YQ. Research progress of pharmacological effects of licorice and glycyrrhizic acid on viral pneumonia[J]. Drug Eval Res, 2020, 43(7):1452-1468.
[23] 丁伊, 王孟清, 罗银河, 等. 五虎汤抑制STAT3通路并调节哮喘小鼠Th2/Th17免疫平衡[J]. 中国病理生理杂志, 2021, 37(5):857-863.
Ding Y, Wang MQ, Luo YH, et al. Wuhu Decoction inhibits STAT3 pathway and regulates Th2/Th17 immune balance in asthmatic mice[J]. Chin J Pathophysiol, 2021, 37(5):857-863.
[24] Lai Y, Zhang Q, Long H, et al. Ganghuo Kanggan decoction in influenza: integrating network pharmacology andpharmacological evaluation[J]. Front Pharmacol, 2020, 11:607027.
[25] Osterlund P, Pirhonen J, Ikonen N, et al. Pandemic H1N1 2009 influenza A virus induces weak cytokine responses in human macrophages and dendritic cells and is highly sensitive to the antiviral actions of interferons[J]. J Virol, 2010, 84(3):1414-1422.
[26] Liu Y, Chen H, Sun Y, et al. Antiviral role of Toll-like receptors and cytokines against the new 2009 H1N1 virus infection[J]. Mol Biol Rep, 2012, 39(2):1163-1172.
[27] Huo C, Jin Y, Zou S, et al. Lethal influenza A virus preferentially activates TLR3 and triggers a severe inflammatory response[J]. Virus Res, 2018, 257(1):102-112.
[28] Weber M, Sediri H, Felgenhauer U, et al. Influenza virus adaptation PB2-627K modulates nucleocapsid inhibition by the pathogen sensor RIG-I[J]. Cell Host Microbe, 2015, 17(3):309-319.
Effect of Zhuang medicine Longpan-Zhike formula on influenza A virus H1N1 pneumonia in mice and its mechanism
ZOU Min1, XIE Cai-ying2, GAN Na1, NONG Zhi-fei3, LI Chong-jin4, WEI Xing2△
(1,,530001,;2,,530200,;3,,530021,;4,,525099,)
To explore the effect of Zhuang medicine Longpan-Zhike formula (LPZKF) on anti-influenza virus pneumonia and its related mechanism.BALB/c mice were randomly divided into control group, model group, high-dose (10.4 g/kg) LPZKF group, low-dose (5.2 g/kg) LPZKF group, and LPZKF (10.4 g/kg)+Toll-like receptor 3 (TLR3) agonist polyinosinic:polycytidylic acid [Poly(I:C), 20 mg/kg] group, with 20 rats in each group. Influenza A virus (IAV) H1N1 intranasal drip method was used to establish a viral pneumonia model. The general state of the mice was observed, and 10 mice were taken from each group to record the 15-day survival rate and survival time. The lung, spleen and thymus indexes were calculated, and HE staining was used to observe the pathological changes of lung tissue. Real-time fluorescence quantitative PCR was used to detect lung viral load, and ELISA was used to measure the levels of interleukin (IL)-6, IL-4, interferon (IFN)-α, IFN-β and IFN-γ in lung homogenates. Western blot was used to detect the expression of TLR3/retinoic acid-inducible gene-I (RIG-I)/nuclear factor-κB (NF-κB) pathway related proteins in lung tissues.All control mice were shown in good mental state and did not die during the experiment. Compared with control group, the mice in model group showed typical flu symptoms after infection, the survival rate, survival time, spleen index and thymus index of the mice were reduced significantly (<0.05), while the lung index, lung histopathological score, viral load, IL-6, IL-4, IFN-γ, IFN-α and IFN-β levels in lung homogenates, IFN-γ/IL-4 ratio in lung homogenates, and TLR3, nuclear NF-κB p65, RIG-I and interferon-β promoter stimulator 1 (IPS-1) expression in lung tissues were increased significantly (<0.05). Compared with model group, the symptoms of the mice in high- and low-dose LPZKF groups were attenuated significantly, the survival rate, survival time, spleen index, thymus index, IFN-α and IFN-β levels in lung homogenates, and RIG-I and IPS-1 expression in lung tissues were increased significantly (<0.05), while the lung index, lung histopathological score, viral load, IL-6, IL-4 and IFN-γ levels in lung homogenates, IFN-γ/IL-4 ratio in lung homogenates, and TLR3 and nuclear NF-κB p65 expression in lung tissues were reduced significantly (<0.05). Moreover, Poly(I:C) significantly blocked the protective effect of LPZKF on the lung of IAV H1N1-infected mice.Zhuang medicine Longpan-Zhike formula reduces lung injury in mice infected with IAV H1N1, and the mechanism may be related to TLR3/RIG-I/NF-κB signaling pathway and inhibiting excessive innate inflammatory response.
Zhuang medicine Longpan-Zhike formula; Influenza A virus H1N1; Pneumonia; Toll-like receptor 3; Retinoic acid-inducible gene I; Nuclear factor-κB
R285.5; R363
A
10.3969/j.issn.1000-4718.2022.03.018
1000-4718(2022)03-0526-09
2021-10-26
2022-01-04
[基金项目]广西高校中青年教师基础能力提升项目(No. 2018KY0302);2021年广西国际壮医医院院级课题(No. GZ2021012)
Tel: 13877196656; E-mail: 614486902@qq.com
(责任编辑:卢萍,罗森)
