生长时间和栽培模式对柴胡根际微生物群落结构的影响
2022-03-15冯海洋高德民
刘 丽,范 娅,冯海洋,杜 衎,高德民
(山东中医药大学药学院,济南 250355)
【研究意义】柴胡(BupleurumchinenseDC.)是伞形科柴胡属多年生药用植物,以干燥根入药。柴胡是我国常用的传统药材,已有数千年的用药历史,具有疏风解表、疏肝解郁、升举阳气等功效[1]。柴胡属大宗药材,市场需求量很大,但野生柴胡资源有限,因此栽培柴胡是市场的主要来源[2]。柴胡对环境因子,尤其是土壤温度、湿度、海拔等非常敏感,因此适宜的产地有限,导致其栽培地柴胡重茬连作现象十分普遍。柴胡种植二年后即可采收,而长期连作造成柴胡根腐病加剧,生长势减弱,死苗、烂根现象突出,种质退化严重,药材质量和产量下降,产生连作障碍。因此,研究药材连作障碍的发生机理机制对于提出合理有效的消减措施,保障中药材生产的可持续发展和药材质量的安全性具有重要的理论和现实意义。【前人研究进展】连作障碍发生的机理非常复杂,既包括根际土壤理化环境失衡、土壤微生物区系改变等土壤因素,也包括植物本身的化感自毒作用、虫害叠加累积效应等[3-4]。近年来,从植物-土壤-微生物组成的根际微生态系统探讨连作障碍的成因和消减措施已成为一个重要的研究方向,并在西洋参、黄芪[5-6]等药材生长中取得了重要进展,其连作障碍的根际微生物机制也得到了初步的阐明,并且通过轮作、间作等栽培模式来效缓解连作障碍造成的影响。【本研究切入点】以柴胡不同生长时间和不同栽培模式的根际微生物为切入点,探讨其变化机制。【拟解决的关键问题】采用高通量测序技术,比较柴胡不同生长时间、不同栽培方式根际微生物的区系变化,从根际微生物角度探讨柴胡连作障碍的发生机制,为缓解生产难题,加快柴胡优质道地药材基地建设,促进柴胡种植业健康发展提供依据。
1 材料与方法
1.1 样品采集
山东省章丘区年平均日照2647.6 h,日照率60%。年平均气温12.8 ℃,年平均降水量600.8 mm,土壤类型为棕壤。山东省长清区年平均日照时数为2447.6 h,平均气温14.4 ℃,平均降水量661.7 mm,土壤类型为棕壤。甘肃省宕昌县年平均日照2100 h,年均气温9.3 ℃,平均降水量584 mm,土壤类型为黄棕壤。
本研究采集的柴胡包括:柴胡不同生长期(一年生(B_1),二年生(B_2)和三年生(B_3))、不同栽培模式(二年生柴胡与玉米间作(B_4)和三年生柴胡与玉米轮作(B_5))、不同产地两年生柴胡(B_6和B_7)。间作柴胡的密度为80 000 株/667 m2,玉米的密度为4000 株/667 m2。玉米的种植时间为5月,柴胡的种植时间为7月。采样时间为2020年8月。采集样品的详细信息见表1。
1.2 根际土壤采集
采用五点采样法采集根际土壤样品。每个样品随机选择3个面积为2 m×2 m的样方,将样方中柴胡整株取出,采用抖根法收集根际土壤,同一样方内的土壤作为一个混合样[7]。所有土壤样品入无菌袋中密封,立即带回实验室,过2 mm筛,于-20 ℃保存备用,用于微生物分析。
1.3 DNA 提取、PCR 扩增
用土壤DNA 试剂盒(Mo Bio Laboratories,Carlsbad,CA,USA)提取样品DNA。 PCR扩增土壤细菌16S rDNA V4~V5区和土壤真菌ITS1(94 ℃预变性2 min,接着30个循环,94 ℃变性30 s;55 ℃退火30 s;72 ℃延伸45 s;72 ℃延伸10 min,最后保温10 min)。引物为515F 5′-GTGCCAGCMGCCGCGGTA A-3′,926R 5′-CCGTCAATTCMTTTGAGTTT-3′,ITS 1F5′-CTTGGTCATTTAGAGGAAGTAA-3′和 ITS1R 5′-GCTGCGTTCTTCATCGATGC-3′。PCR结束后对扩增产物进行琼脂糖电泳检测并回收。
1.4 建库和测序
使用TruSeq DNA PCR-Free Sample Prep Kit建库,构建好的文库经过定量和文库检测合格后,在上海微基生物科技有限公司通过 Illumina MiSeq平台上进行模式测序。
1.5 数据分析
采用Mothur(version 1.33.3)软件对序列质量进行质控和过滤,将模糊碱基、单碱基高重复区、过长和过短的序列以及PCR过程中产生的一些嵌合体去除,得到优化序列。采用UPARSE 软件(Usearch Version V8.1.1756,http://drive5.com/uparse/)进行OTU (Operational taxonomic unit) 聚类。基于分类学信息,在门和属水平上对柴胡根际微生物群落结构进行统计分析。使用Mothur(version 1.33.3)软件进行微生物群落多样性分析。在上述分析的基础上,通过R软件(version 3.6.0)计算并可视化稀释曲线、Venn图、Heatmap、PCoA和群落结构直方图。
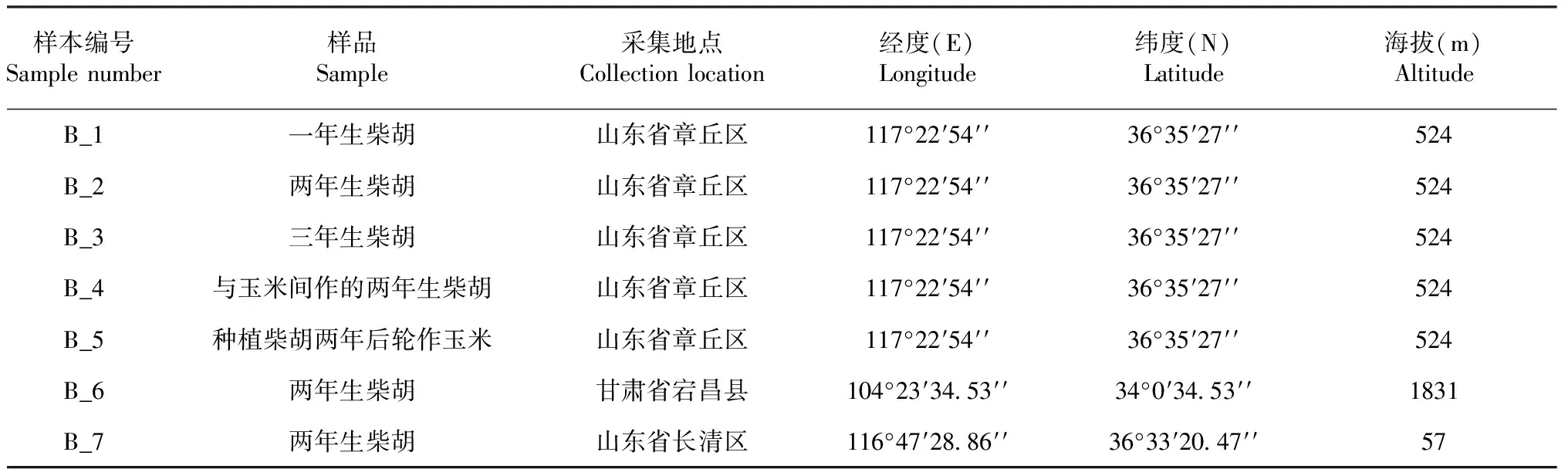
表1 不同生长时间和栽培模式下柴胡的根际土壤信息
2 结果与分析
2.1 高通量测序结果
7个根际土壤样品的稀释曲线(图1)显示,初始阶段OTU呈现急剧上升,但随着样本测序序列数目不断增加,OTU总数将不再随着序列数目的加入而显著增加,说明测序文库已经达到饱和,测序结果可用来后续分析。高通量结果见表2。
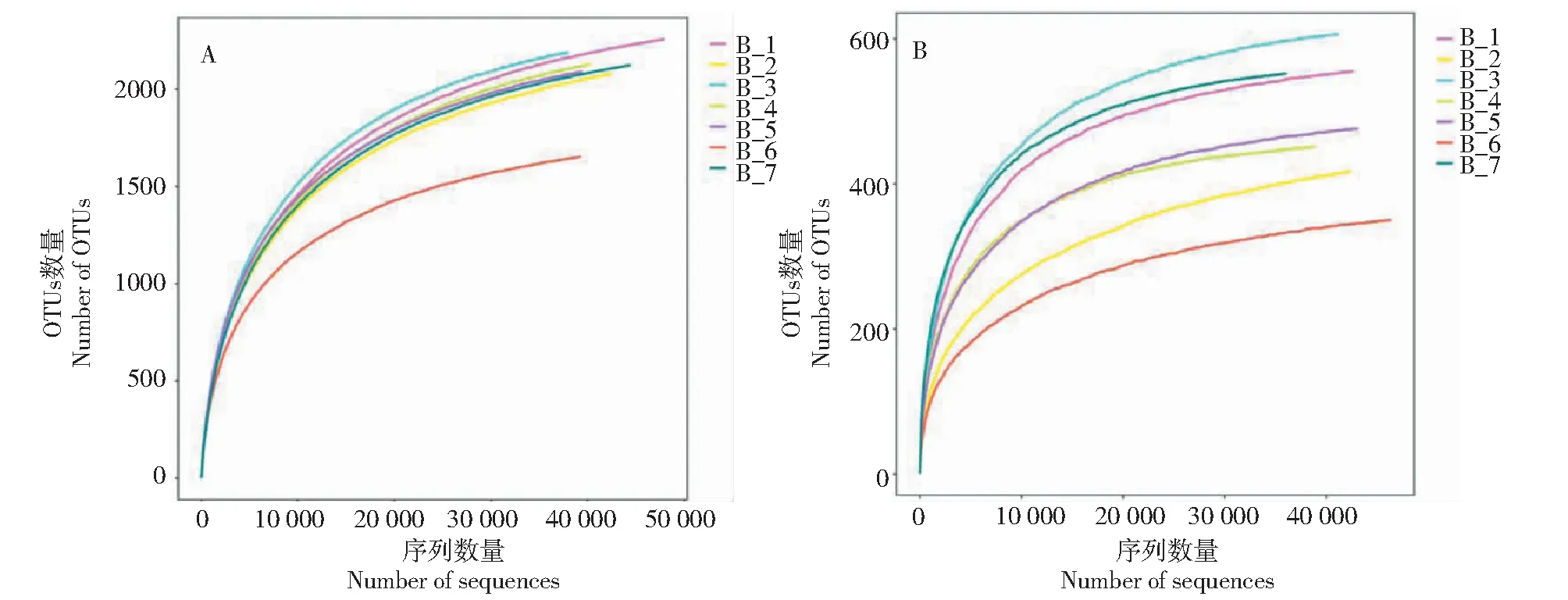
对于每个样品OTU,分别以97%的序列一致性进行分类分析。从统计学上确定每个样本常见或特有的OTUs(图2)。柴胡连续种植后土壤细菌的特有OTUs数目呈下降趋势,而与玉米间作和轮作后特有OTUs数目升高。土壤真菌在连续种植过程中特有OTUs数量呈现先减少后增加的趋势。与玉米间作后土壤真菌的特有OTUs数量增加,而轮作后减少。宕昌市和长清区两年生柴胡土壤中特有OTUs数量显着高于章丘区。
2.2 α-多样性分析
根际微生物的丰富度(ACE指数和 Chao指数)和群落多样性(Simpson指数,表 3)显示,柴胡连续种植后根际土壤细菌丰富度和多样性下降,尤其是生长二年的柴胡。而连续种植后柴胡的真菌丰富度和多样性呈现先降低后升高趋势。但柴胡与玉米间作或轮作后细菌的丰富度和多样性明显提高,并且真菌的多样性也增加。区域位置对柴胡根际土壤微生物丰富度和多样性影响较大,其中山东长清地区真菌丰富度和多样性为两年生中最高。
B_1、B_2和B_3分别代表中国山东章丘区一年生、两年生、三年生柴胡。B_4和B_5分别代表二年生柴胡与玉米间作和三年生柴胡与玉米轮作。B_6和B_7分别代表甘肃省宕昌市和山东省长清区的两年生柴胡。以下图中样品均相同。A.细菌,B.真菌B_1,B_2 and B_3 represent 1-year-old Bupleurum chinense DC.,2-year-old Bupleurum chinense DC.and 3-year-old Bupleurum chinense DC.in Zhangqiu areas,Shandong province,China,respectively.B_4 and B_5 represent 2-year-old intercropped Bupleurum chinense DC.,3-year-old rorated Bupleurum chinense DC.,respectively.B_6 and B_7 represent 2-year-old Bupleurum chinense DC.from Dangchang areas,Gansu province and Changqing Shandong province,respectively.The following soil samples in the figures are all the same.A.Bacteria,B.Fungi图1 不同生长时间和栽培模式下柴胡根际微生物稀释曲线Fig.1 Rarefaction curves of microbiome for rhizosphere soil samples from different growth time and cultivation pattern of Bupleurum chinense DC.

表2 柴胡根际土壤中细菌和真菌的高通量结果
A.细菌,B.真菌A.Bacteria,B.Fungi图2 不同生长时间和栽培模式下柴胡根际微生物的操作分类单元(OTU)相似度和重叠度的Venn分析Fig.2 Venn analysis of similarity and overlap of operational taxa (OTU) of rhizosphere microorganisms of Bupleurum Chinese DC.under different growth times and cultivation patterns

表3 根际微生物的 ACE、Chao和 Shannon 指数
2.3 β-多样性分析
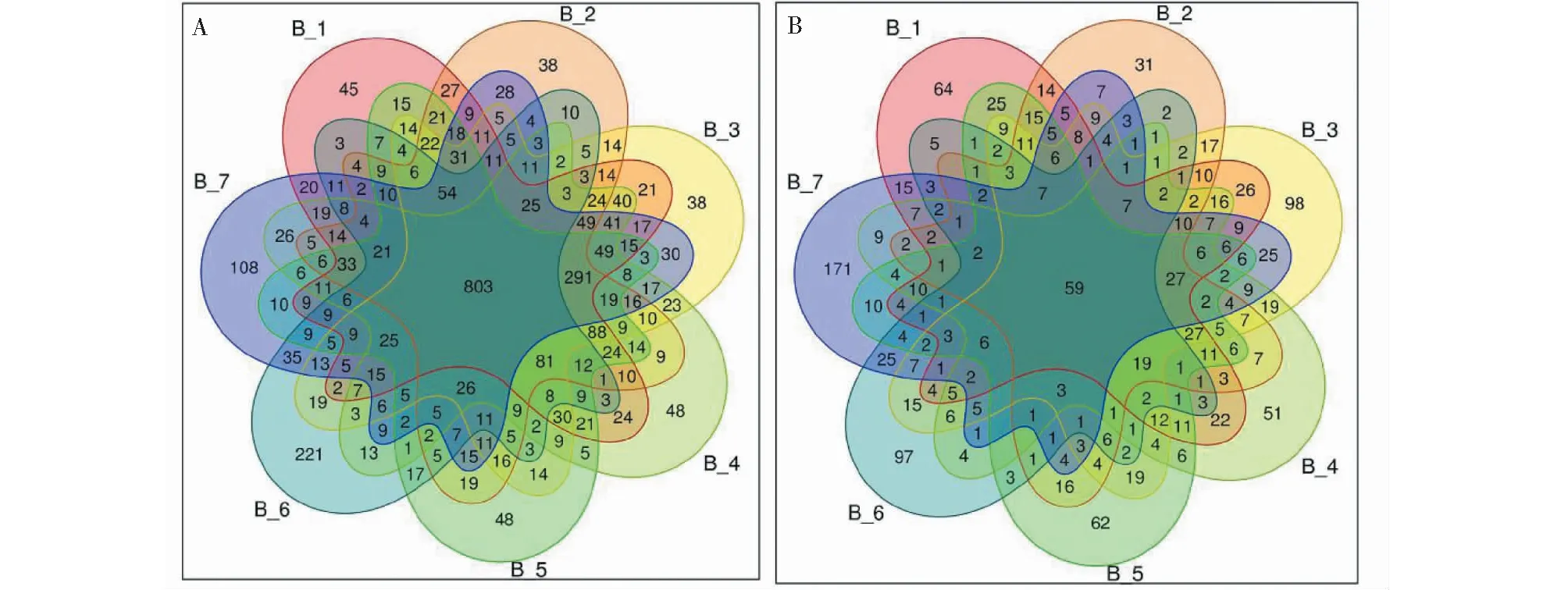
柴胡根际细菌的PCoA图(图3)可以较好的反映出细菌多样性的差异,因为两个主坐标轴的总解释度达到75.98%。同样,真菌变化趋势与细菌基本一致。柴胡经过间作和轮作后,样品间的距离增大,表明土壤根际微生物多样性差异增大。不同产地生长两年的柴胡样本分布相对离散的,说明样品间细菌和真菌多样性有显著差异。
基于样品间的聚类分析,7个柴胡样品的根际土壤细菌样品分为4组(图4-A)。B_2、B_5为第一组,B_1、B_3、B_4为第二组,B_6、B_7各单独成组。组内细菌相似性较高,但组间细菌群落差异较大,而不同生长时间柴胡根际细菌群落的差异最为明显。同样,7个柴胡根际土壤真菌样品被分六组,其中B_5和B_4聚为一类,说明这两个样品中的真菌群落组成具有较高的相似性,B_1、B_3、B_2、B_6和B_7分别单独成组,这说明柴胡样品根际真菌群落明显不同。
2.4 根际微生物组成
为了阐明柴胡根际土壤中微生物群落结构,在两个分类水平(门和属)进行了分析。7个土壤样品中共检测到 14 个细菌门和 8 个真菌门。由图5-A可知,柴胡根际细菌的优势门为变形菌门(Proteobacteria,34%),其次是酸杆菌门(Acidobacteria,21%)、放线菌门(Actinobacteria,14%)和拟杆菌门(Bacteroidetes,7%)。变形菌门的相对丰度在柴胡连续种植后增加,但与玉米间作和轮作后相对丰度较低。两年生柴胡的变形菌门相对丰度普遍高于其它生长时间。山东长清区变形菌门相对丰度在所有地区中最高。酸杆菌门随柴胡生长时间相对丰度逐年下降,但与玉米间作或轮作后相对丰度明显升高。值得注意的是,甘肃宕昌市柴胡根际土壤中酸杆菌
A.细菌,B.真菌A.Bacteria,B.Fungi图3 不同生长时间和栽培模式下根际土壤微生物群落结构主坐标分析(PCoA)Fig.3 The principal coordinate analysis (PCoA) of rhizosphere soil microbiome community structure under different growth times and cultivation patterns

A.细菌,B.真菌A.Bacteria,B.Fungi图4 不同生长时间和栽培模式下微生物群落操作分类群的聚类分析Fig.4 Cluster analysis of microbiome operating taxa under different growth times and cultivation patterns
门相对丰度在所有地区中最低。
由图5-B可知,柴胡根际真菌的优势门依次是子囊菌门(Ascomycota,78%)、担子菌门(Basidiomycota,9%)和接合菌门(Zygomycota,4%)。子囊菌门相对丰度随柴胡生长时间的增加有所下降,二年生柴胡尤为明显。但与玉米间作和轮作后相对丰度增加。不同产地种植的两年生柴胡中山东章丘区与长清区的子囊菌门相对丰度偏低,而甘肃宕昌县的子囊菌门相对丰度高于其他组。担子菌门的相对丰度随着柴胡生长时间显著增加,但与玉米间作或轮作后相对丰度较低。山东章丘区柴胡根际土壤担子菌相对丰度普遍高于其它地区。
在属水平(图6)上,7个根际土壤中检测到57个细菌属和50个真菌属。优势细菌属为假杆菌属(Pseudarthrobacter,2%)、布氏杆菌属 (Bryobacter,1%)、 类固醇杆菌属(Steroidobacter,1%)、硝化螺菌属 (Nitrospira,1%)。Pseudoarthrobacter的相对丰度随柴胡生长时间逐年增加,但与玉米间作和轮作后减少。布氏杆菌属、 类固醇杆菌属和硝化螺菌属的相对丰度在连续种植3年后下降,但在间作和轮作后相对丰度发生逆转。
根际土壤真菌的优势属为光黑壳属(Preussia,7%)、毛壳属(Chaetomium,7%)、赤霉菌属(Gibberella,7%)、镰刀菌属(Fusarium,6%)、被孢霉属(Mortierella,4%)、尾柄孢壳属(Cercophora,3%)、隐球菌属(Cryptococcus,2%)。光黑壳属、毛壳属和被孢霉属的相对丰度随柴胡生长时间下降,而与玉米在间作和轮作后相对丰度增加。

A.细菌,B.真菌A.Bacteria,B.Fungi图5 不同生长时间和栽培模式下微生物水平的物种相对丰度Fig.5 Relative abundance of species at the microbial level under different growth times and cultivation modes
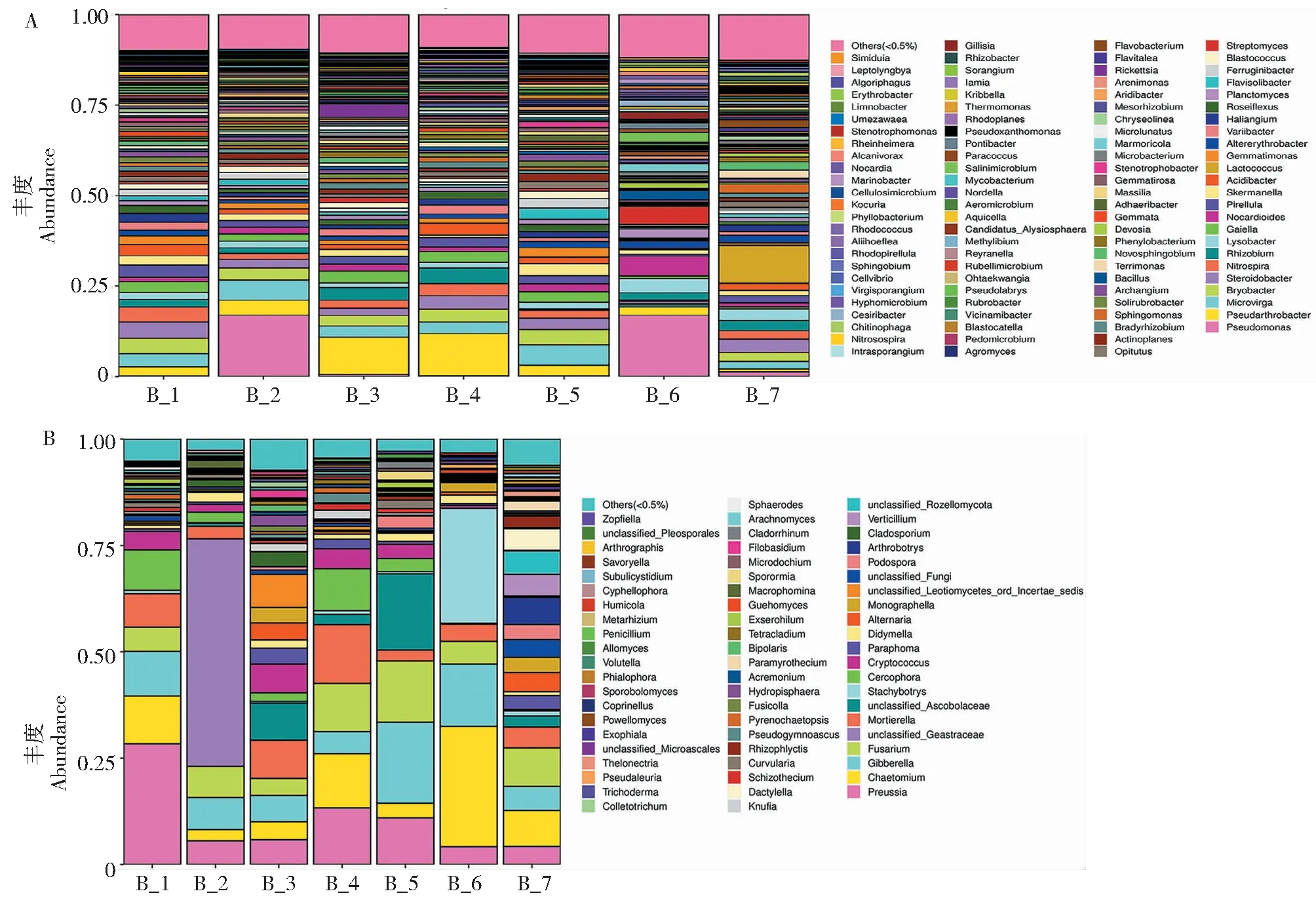
A.细菌,B.真菌A.Bacteria,B.Fungi图6 不同生长时间和栽培模式下微生物属水平的物种相对丰度Fig.6 Relative abundance of species at the level of microbial genus under different growth times and cultivation modes
柴胡根际微生物群落的组成受样品生长时间、地点以及栽培模式的影响很大。例如,Lactococcus是山东省长清区的特有属,而Salinimicrobium、Gillisia、Cesiribacter、Marinobacter和Alcanivorax则是甘肃省宕昌市的特有属。Rickettsia、Dactylella、Subulicystidium和Savoryella仅存在于两年生柴胡中。Allomyces是柴胡轮作的特有属。
3 讨 论
在农业生态系统中,土壤微生物多样性对维持生态系统的平衡起着重要作用。土壤微生物群落是土壤生态功能的基础[8]。本研究采用Illumina高通量技术对柴胡不同生长时间、不同栽培模式下的根际土壤微生物群落结构和组成进行了分析。表明生长时间和栽培模式对柴胡根际微生物的多样性和丰富度有显著影响。柴胡连续生长3年以及连坐后根际土壤中细菌和真菌的多样性和丰富度明显降低,但与玉米轮作或间作后则逆转了这一变化趋势。这与许多药材根际微生物的变化规律是基本一致的[9-11]。本研究进一步丰富了栽培模式对柴胡根际微生物动态变化规律,这对于调节柴胡种植结构、优选柴胡种植模式、加强种植管理具有重要的理论和实践意义[12]。
根际微生物受环境影响很大,如土壤类型、温度、含水量、生长时间和栽培模式等,都会影响微生物群落的类型和结构[13]。与其他产地的微生物群落相比,山东长清区柴胡根际真菌丰富度最高,但山东章丘区真菌多样性最高。这与二者环境因子明显不同有关[14]。尽管柴胡根际微生物在物种多样性、相对丰度、结构和分类组成等方面存在一定差异,但同一生境下微生物群落结构是相似的。同时研究发现,药用植物根际土壤细菌主要来源于变形菌门和酸杆菌门,真菌主要来源于子囊菌门和担子菌门,且物种丰度也较高,属于优势菌属[15-17]。与柴胡根际土壤微生物群落比较后发现,不同药用植物根际土壤微生物在物种组成上具有较高的相似性,其主要的差别在于物种的丰度差异,这可能也与土壤本身的微生物种群有关系[18]。
微生物群落结构和组成的变化通常会导致代谢能力、生物降解、疾病抑制和其他功能的变化[19]。柴胡连续种植大大减少了有益微生物的丰度,例如,布氏杆菌属(Bryobacter)、硝化螺菌属(Nitrospira)、光黑壳属(Preussia) 、毛壳属(Chaetomium)和被孢霉属(Mortierella),这些根际微生物在氮循环、微生物代谢和生物防治方面起着重要作用[20-21]。Bryobacter是柴胡根际微生物中的高丰度菌属,具有促进土壤碳循环作用[22]。此外,丰度较高的光黑壳属和毛壳属对植物病原物的生物防治具有重要意义[23-24]。相反,一些植物病原体,例如链格孢属和炭疽菌属在连续种植后大量增加[25-26]。链格孢属和炭疽菌属在连续种植3年后丰度分别为2.11%和0.66%。链格孢属为重要植物病原菌,是根腐病[27]和黑斑病的致病菌[28],可造成多种作物的严重病害。炭疽菌属可引起苹果、茶树、铁皮石斛等多种植物产生炭疽病,导致农产品的品质下降,造成巨大经济损失[29-33]。
4 结 论
生长时间和栽培模式是影响柴胡的根际微生物群落差异的重要原因,而地理位置也是影响群落结构与组成的原因之一。可以通过调整柴胡与玉米间作或轮作等栽培模式来保持根际微生物的菌落结构稳定,促进柴胡种植业的健康发展。
