黑曲霉丝氨酸羧肽酶的重组表达及其水解大豆蛋白的效果分析
2022-02-15喻豆潘力王斌
喻豆,潘力,王斌
(华南理工大学生物科学与工程学院,广东省发酵与酶工程重点实验室,广东广州 510006)
关键字:丝氨酸羧肽酶;黑曲霉;重组表达;蛋白脱苦
羧肽酶(Carboxypeptidases,E.C.3.4.16-E.C.3.4.18)是一种外肽酶,能够依次水解释放蛋白质和多肽C末端的氨基酸[1]。根据羧肽酶的活性中心可以分为丝氨酸羧肽酶(E.C.3.4.16)、金属性羧肽酶(E.C.3.4.17)和半胱氨酸羧肽酶(E.C.3.4.18)[2]。丝氨酸羧肽酶是一种能够在酸性条件下逐个水解肽链C末端氨基酸的外肽酶,属于丝氨酸羧肽酶的S10家族,能够参与蛋白质在生物体内的运输、靶向和加工过程,在酸性条件下除了能够水解肽链末端之外还具有酯酶和脱酰胺酶的活性[3]。除此之外丝氨酸羧肽酶具有许多共同的结构特征,有一个S-D-H(丝氨酸、天冬氨酸、和组氨酸)组成的三联体结构,其中Ser是活性中心。目前已报道的丝氨酸羧肽酶来源很广泛,存在于动植物组织和各种微生物中[4-6]。丝氨酸羧肽酶在曲霉中的研究较少,且产量较低[7-14]。已报道的文献中表达量较高的是Morita[8]等将Aspergillus oryzae的丝氨酸羧肽酶基因cpI在Aspergillus nidulans中进行过量表达测得的85.31 U/mL。
大豆蛋白是食品加工行业中应用广泛的原材料,其水解液中含有许多活性多肽,可作为功能性食品成分和营养添加剂等。但在大豆蛋白水解过程中会产生大量的苦味肽,严重限制其在食品加工行业的应用。苦味主要源于蛋白水解过程中生成的苦味肽,而苦味肽中的疏水性氨基酸是苦味产生的主要因素。在蛋白质中由于疏水性氨基酸被包埋在分子内部,故而不会被品尝出苦味,这也是分子量大于6000 u的多肽不会呈现出苦味的原因。当疏水性氨基酸在分子表面的暴露程度越高,产生的苦味越重。但当疏水性氨基酸位于肽链内部和以游离形式存在时,其结构较少能与苦味受体结合,因此呈现的苦味较轻。与游离形式相比,当疏水性氨基酸位于肽链末端时的苦味较重。当疏水性氨基酸位于肽链的C末端时产生的苦味较位于N末端时更重,这表明氨基酸的顺序与苦味值相关。此外肽链的长度、疏水值和氨基酸的组成也会对苦味产生较大的影响[15,16]。在大豆蛋白的水解液中含有Gly-Leu、Leu-Lys、Arg-Leu、Phe-Leu、Arg-Leu-Leu等多种苦味肽。降低苦味的方法主要有物理法和生物酶解法,其中生物酶解法具有反应条件温和、产物安全可靠等优点。生物酶解法中使用的蛋白酶主要分为内肽酶、外肽酶和 plastein蛋白酶[17-20]。内肽酶主要包括胃蛋白酶、胰蛋白酶、木瓜蛋白酶和碱性蛋白酶等,氨肽酶是在脱苦研究中使用较多的一类外肽酶。单独使用一种生物酶时水解液中仍会存在较多苦味肽,但将内肽酶与外肽酶协同酶解大豆蛋白时的苦味降低最明显。
黑曲霉(Aspergillus niger)具有强大的蛋白分泌能力,使其能够承担蛋白高表达时的分泌压力;具有完善的转录后和翻译后修饰系统(在表达真核基因产物时具有明显的优势);被认为是 GRAS(Generally Regarded as Safe)安全菌株,具有生物安全性;基因编辑技术在黑曲霉中广泛应用,使得基因操作更加简单可靠;黑曲霉因环境适应能力较强,发酵时所用的培养基由价格相对廉价的基础营养物质组成;最后黑曲霉可进行高密度发酵且发酵放大技术较成熟[21-25]。因此黑曲霉是优秀的蛋白表达宿主。
本研究以低蛋白背景的无孢黑曲霉A.nigerHL-1为宿主,表达A.nigerCBS 513.88中3个丝氨酸羧肽酶基因(CPA、CPF、CPG),筛选得到高活力的丝氨酸羧肽酶基因CPG,并研究了重组酶CPG的酶学性质,最后将其应用于大豆蛋白脱苦,研究了胃蛋白酶与重组酶CPG复配所产生的脱苦效果,为解决大豆蛋白水解液脱苦处理工艺提供支持。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒
黑曲霉宿主菌株A.nigerHL-1(ΔpyrG、ΔglaA)由本实验室构建并保藏,通用表达载体 UEV由本实验构建并保藏,大肠杆菌Escherichia coliMatch1 T1购自美国Invitrogen公司
1.1.2 试剂限制性内切酶ApaⅠ和XbaⅠ,美国 Thermo Fisher Scientific公司;Prime STAR HS DNA Polymerase (premix),日本TaKaRa公司;HiFi DNA Assembly Cloning Kit试剂盒,美国 NEB公司;DreamTaq Green PCR Master Mix,美国Thermo Fisher Scientific公司;胃蛋白酶、茚三酮和氯化镉,上海麦克林生化科技有限公司;FA-Phe-Ala,成都云希化工有限公司合成;大豆分离蛋白,广州普博欣生物科技有限公司;DNA marker(250 bp Ladder、DL 5000TM)和26616 Protein Marker,日本TaKaRa公司。

表1 引物Table 1 Primer

表2 丝氨酸羧肽酶基因序列Table 2 The sequence of serine carboxypeptidase genes
1.1.3 培养基
LB培养基:1%蛋白胨、1%氯化钠、0.5%酵母提取物。
DPY培养基:2%葡萄糖、1%蛋白胨、0.5%酵母提取物、0.1% KH2PO4、0.05% MgSO4·7H2O。
CD培养基:2%葡萄糖、0.3% NaNO3、0.2% KCl、0.1% KH2PO4、 0.05% MgSO4·7H2O 、 0.001%FeSO4·7H2O、2%琼脂粉。
发酵培养基:5%玉米淀粉、3%玉米浆、2%豆粕粉。
1.1.4 引物
实验中所使用的引物如表1所示。
1.2 仪器
Veriti 96-Well Thermal Cycler聚合酶链式反应(polymerase chain reaction,PCR)仪,美国Applied Biosystems公司;AKATA层析仪,美国通用电气公司;浸入式水平电泳系统,美国Bio-Rad公司;M200多功能酶标仪,德国TECAN公司;移液枪和高速离心机,德国Eppendorf公司。
1.3 方法
1.3.1 丝氨酸羧肽酶表达载体的构建
1.3.1.1 本章所选取的羧肽酶基因
从 Uniprot上筛得到的黑曲霉丝氨酸羧肽酶基因序列如表2所示。
1.3.1.2 表达载体构建
通过SignalP 4.0 server信号肽预测基因原有信号肽,分别构建信号肽为自身信号肽和糖化酶信号肽的丝氨酸羧肽酶表达载体。以黑曲霉A.nigerCBS 513.88的基因组为模板,以 CPG-F和 CPG-R,CPF-F和CPF-R,CPA-F和CPA-R为引物扩增信号肽为自身信号肽的CPG,CPF和CPA基因片段,以CPG-g-F和CPG-g-R,CPF-g-F和CPF-g-R,CPA-g-F和CPA-g-R为引物扩增信号肽为糖化酶信号肽的CPG-g、CPF-g和CPA-g基因片段(每个目的基因的C端均通过引物加入6ⅹHis标签),用PnaⅡ-F和PnaⅡ-R为引物扩增杂合启动子片段PnaⅡ。将目的基因(CPG,CPF、CPA、CPG-g、CPF-g或CPA-g)、杂合启动子PnaⅡ和线性化的通用载体 UEV(含pyrG筛选标记、Ttef终止之和amyA同源臂)通过HiFi DNA连接酶的作用连接成环形质粒,转化至大肠杆菌E.coliMatch1 T1感受态细胞,经过氨苄青霉素的抗性筛选、菌液电泳、酶切验证和测序验证后筛选出正确的表达载体。构建正确的表达载体命名为 UEV-CPG/CPF/CPA/CPG-g/CPF-g/CPA-g。
1.3.2 丝氨酸羧肽酶重组菌株的构建
将 6个丝氨酸羧肽酶表达载体(UEV-CPG、UEV-CPF、UEV-CPA、UEV-CPG-g、UEV-CPF-g和UEV-CPA-g)经过质粒大量提取、浓缩后,通过PEG介导转化至无孢黑曲霉A.nigerHL-1原生质体中,30 ℃恒温培养。将转化子挑至CD板上继续培养。待转化子长大之后提取基因组。以转化子基因组模板,利用验证引物CP-F和CP-R验证目的基因表达框是否完整整合至转化子基因组上。筛选正确的丝氨酸羧肽酶重组菌株,并将正确的菌株命名为 HL-CPG/CPF/CPA/CPG-g/CPF-g/CPA-g。
1.3.3 丝氨酸羧肽酶酶活测定
将鉴定正确的丝氨酸羧肽酶菌株在发酵培养基中发酵168 h(30 ℃,250 r/min),每24 h取样一次。利用镉-茚三酮法测定丝氨酸羧肽酶的酶活。以FA-Phe-Ala为底物,将其溶于pH 4.5,45 mmol的乙酸钠缓冲液中配制成0.5 mmol的底物溶液。取已用乙酸钠缓冲液稀释的粗酶液 100 μL加入 900 μL FA-Phe-Ala溶液,混匀,30 ℃反应30 min。向450 μL无菌水中加入50 μL反应液,然后加入1 mL显色液即镉-茚三酮溶液,84 ℃水浴5 min,冰上冷却至室温,用酶标仪测量A506 nm的吸光值,对照组将100 μL粗酶液煮沸 15 min使酶失活,其余操作与实验组相同。酶活定义:30 ℃,pH 4.5的条件下,每小时水解1 μmoL丙氨酸所需要的酶量为1个酶活力单位(U)。
1.3.4 丝氨酸羧肽酶的纯化
因在构建表达载体时在目的基因中加入 6ⅹHis标签,故可以利用镍柱亲和层析的方法进行丝氨酸羧肽酶的纯化。将重组菌株在发酵培养基中培养96 h,将发酵培养基离心取上清后用0.45 μm的滤膜过滤,将其用于纯化。使用HisTrapTMHP层析柱,上样量为20 mL,流速为1 mL/min。采用梯度洗脱法(洗脱液中咪唑的浓度为0~0.5 mol/L)收集不同的洗脱峰。利用SDS-PAGE检验每个洗脱峰,得到单一的丝氨酸羧肽酶溶液。
1.3.5 丝氨酸羧肽酶的酶学性质研究
1.3.5.1 温度对丝氨酸羧肽酶的影响
最适温度测定:在pH值为4.5的条件下,将纯化得到的蛋白酶液分别在20、40、50、60、70、80 ℃条件下测定丝氨酸羧肽酶的酶活,以最高的酶活力为100%。温度稳定性测定:将酶液在上述温度下保温处理2 h,测定剩余的酶活力,以未经处理组的酶活力为100%。
1.3.5.2 pH对丝氨酸羧肽酶的影响
最适pH值测定:配制不同pH值(2.5、3.0、3.5、4.0、4.5、5.0、5.5、6.0、6.5、7.0、8.0)0.5 mmol的FA-Phe-Ala底物溶液,在测量得到的最适温度下,测定不同 pH值条件的酶活变化,以最高的酶活力为100%。pH稳定性测定:将酶液在上述pH条件下保温处理2 h,测定酶液剩余的酶活力,以未经处理组的酶活力为100%。
1.3.5.3 金属离子对丝氨酸羧肽酶的影响
在标准的酶液反应体系下,加入不同的金属离子(Ba2+、Mn2+、K+、Mg2+、Ni2+、Co2+、Ca2+、Na+、Zn2+、Cu2+),在反应体系中每种离子的终浓度为 5.0 mmol,测定丝氨酸羧肽酶的酶活,以未经处理组的酶活力为100%。
1.3.6 丝氨酸羧肽酶在大豆蛋白脱苦中的应用
1.3.6.1 甲醛滴定法测定水解度
取5 mL SPI水解样品加入60 mL超纯水,用0.1 mol/L NaOH调节样品pH至8.2。向溶液中缓慢加入10 mL pH值为8.2中性甲醛溶液,用0.01 mol/L NaOH进行滴定实验,滴定至pH 9.2,记录NaOH标准溶液的消耗体积V。空白对照组为将5 mL SPI样品换成5 mL超纯水,其他步骤与样品组一致,记录NaOH标准溶液的消耗体积V0。按以下公式计算水解样品的水解度[23]:
式中:
C——样品中原大豆分离蛋白浓度,g/L;
0.33——大豆分离蛋白中游离氨基的浓度,mmol/g;
7.8——每克大豆分离蛋白的肽键当量数,mmol/g。
1.3.6.2 不同pH值条件下胃蛋白酶水解大豆分离蛋白的研究
配制pH为3、3.5、4、4.5的缓冲液溶解大豆分离蛋白。胃蛋白酶的添加量为1:100(酶:底物),温度为37 ℃,每隔1 h测定其水解度,酶解时间为8 h。
1.3.6.3 胃蛋白酶与丝氨酸羧肽酶复配水解大豆分离蛋白
在pH为3.5时测定胃蛋白酶与丝氨酸羧肽酶复配对大豆分离蛋白的影响。实验组:向经过预处理的大豆分离蛋白中按照1:100的酶量加入胃蛋白酶,酶解处理6 h,再加入300 U/g的丝氨酸羧肽酶酶液CPG反应6 h,反应结束时测量水解度。对照组处理方法为经过预处理的SPI中加入相同酶量的胃蛋白酶,反应12 h,反应结束时测量水解度并测定水解液中游离氨基酸的含量。
1.3.7 数据处理与分析
所有试验重复三次,所有的实验数据采用采用SPSS 19.0进行方差分析,Origin进行图表的绘制。
2 结果与讨论
2.1 重组丝氨酸羧肽酶菌株的筛选
将黑曲霉A.nigerCBS 513.88中3个丝氨酸羧肽酶基因(CPA、CPF、CPG)在 SignalP 5.0 server(http://www.cbs.dtu.dk/services/SignalP)分析工具上进行信号肽预测发现3个基因均含有N端的分泌信号。
为构建正确的丝氨酸羧肽酶表达载体,根据表 1中的引物进行 PCR扩增得到分别含自身信号肽和糖化酶信号肽的丝氨酸羧肽酶基因片段(图 1a),将丝氨酸羧肽酶基因片段、启动子PnaⅡ和线性化的UEV通用载体片段进行连接构建表达载体,最终得到测序正确的表达载体 UEV-CPG/CPF/CPA/CPG-g/CPF-g/CPA-g。为了筛选得到高表达的丝氨酸羧肽酶重组菌株,将 6个表达载体(UEV-CPG/CPF/CPA/CPG-g/CPF-g/CPA-g)转化至黑曲霉HL-1宿主中得到6种丝氨酸羧肽酶重组菌株,经PCR验证正确后的阳性转化子接种至发酵培养基培养96 h后测定丝氨酸羧肽酶酶活(图 1c)。信号肽为自身信号肽的重组菌株(HL-CPG、HL-CPF、HL-CPA)的酶活均比信号肽为糖化酶信号肽的重组菌株(HL-CPG-g、HL-CPF-g、HL-CPA-g)的酶活高,可能是因为在丝氨酸羧肽酶的N端含有约90个残基的失活前结构域,该结构域能够保护蛋白质不被其他内肽酶切割且与蛋白质的折叠有关,更换信号肽之后可能会影响蛋白质的结构导致羧肽酶的活性部位被剪切[25]。在两个系列的重组菌株中含CPG基因的重组菌株酶活最高,且在6株不同的丝氨酸羧肽酶菌株中酶活最高的为HL-CPG菌株,酶活为163.71 U/mL,与Morita等[8]报道的85.31 U/mL相比,丝氨酸羧肽酶酶活提高了96.61%,说明筛选得到的丝氨酸羧肽酶重组菌株能达到较高的表达水平。
2.2 重组酶CPG的纯化
将经过预处理的HL-CPG菌株的发酵上清液经过镍柱亲和层析并将纯化之后的样品进行SDS-PAGE电泳检测,结果如图2所示,泳道3为纯化之后的蛋白样品,蛋白条带大小与发酵上清液中的大小一致。
2.3 重组丝氨酸羧肽酶CPG的酶学性质
2.3.1 温度对重组酶CPG的影响
重组酶CPG在40 ℃时酶活最高,当温度为80 ℃时仅具有10%左右的相对酶活力(图3a)。另外重组酶在20~50 ℃时酶活相对稳定,在此范围内处理2 h后能保持60%以上的相对酶活力(图3b)。
2.3.2 pH值对重组酶CPG的影响
重组酶CPG在pH为3.5时酶活最高且随着pH值的升高酶活迅速下降,在pH值为9时仅保持10%的相对酶活(图4a)。将重组酶CPG在各pH条件下反应2 h后,在pH值为3~5时保持50%以上的相对酶活力,在中性条件及碱性条件下不能稳定存在(图4b)。
2.3.3 金属离子对重组酶CPG的影响
在各金属离子的浓度为5 mmol/L时,实验结果如表3所示。Cu2+对重组酶CPG具有明显的抑制效果,Ba2+、Mn2+、Mg2+、Ni2+、Co2+、Ca2+、Zn2+对重组酶CPG具有轻微的抑制效果,K+对CPG的作用效果非常小,Na+对羧肽酶CPG有激活的作用,能够提高10%的酶活力。
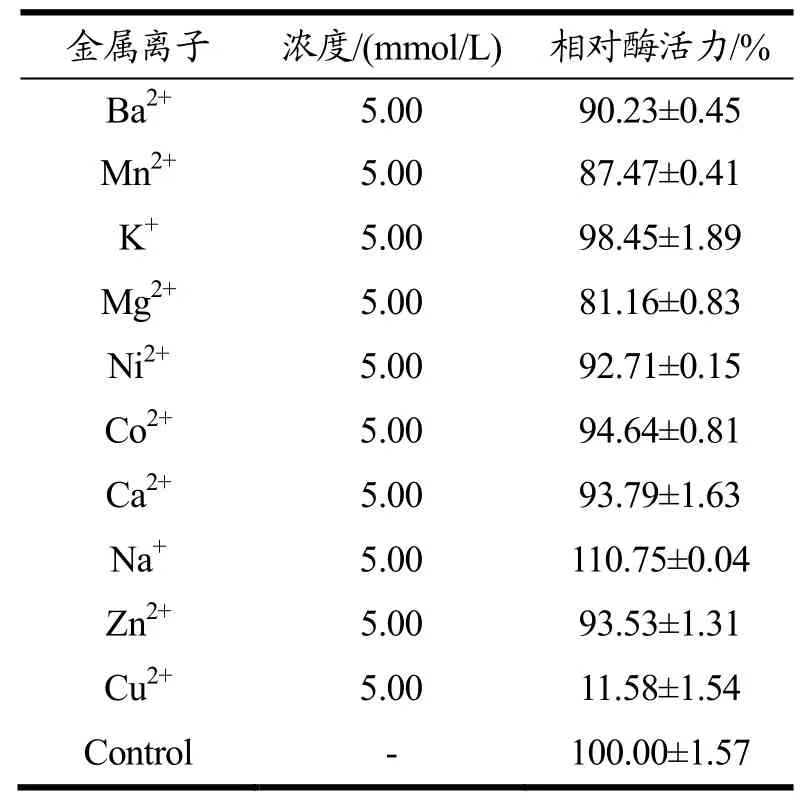
表3 金属离子对纯化后重组酶CPG酶活的影响Table 3 The effect of metal ions on the activity of purified recombinant CPG
2.4 重组丝氨酸羧肽酶CPG在大豆蛋白脱苦中的应用
2.4.1 胃蛋白酶在不同pH值条件下水解大豆蛋白
用胃蛋白酶水解不同pH值条件的大豆分离蛋白,结果如图5所示。在pH值为3时大豆分离蛋白的水解最高。且随着pH值的升高,胃蛋白酶的水解能力逐渐下降,大豆分离蛋白的水解度降低。在所有条件下,前3 h的水解度变化较大,随后趋于稳定并在第6 h时水解度达到最高。在pH值为3.5时大豆分离蛋白的水解度略低于pH值为3时的水解度,但考虑重组酶CPG的酶学性质,选择pH 3.5作为重组酶CPG与胃蛋白酶复配实验的水解条件。
2.4.2 重组酶CPG与胃蛋白酶联合水解大豆蛋白
实验组中加入胃蛋白酶处理6 h之后继续加入重组酶CPG进行处理6 h,对照组中加入相同酶量的胃蛋白酶处理12 h后分别测量水解液的水解度和氨基酸含量。对照组中水解液的水解度为10.17%,而实验组的水解度为19.72%,水解度明显提高。实验组中氨基酸的种类和含量明显高于对照组(图6)。在实验组中Glu、leu、Phe、Tyr和Arg氨基酸的含量较高,疏水性氨基酸 Ala、Val、Met、Ile、Leu、Tyr和 Phe 的含量增加明显,特别是Phe的含量增加最明显,其次是Leu 和 Tyr,含量分别增加 606.47 μg/mL、434.06 μg/mL和205.11 μg/mL。另外实验组中鲜味氨基酸Glu等的含量也明显增加。因此将胃蛋白酶和重组酶CPG复配水解大豆蛋白会明显降低水解液的苦味同时水解液中鲜味也明显增加。
3 结论
3.1 黑曲霉是优秀的蛋白表达宿主,具有强大的蛋白分泌能力,完善的蛋白修饰体系和生物安全性等优点。本研究利用启动子PnaⅡ、终止子Ttef和双向筛选标记pyrG筛选高表达量的丝氨酸羧肽酶基因,并尝试更换信号肽来增高丝氨酸羧肽酶的表达量。通过PCR鉴定挑选表达正确的重组丝氨酸羧肽酶菌株,通过摇瓶发酵筛选高表达量的菌株,最终成功筛选得到重组丝氨酸羧肽酶菌株 HL-CPG,其酶活达到 163.71 U/mL。对重组丝氨酸羧肽酶CPG进行镍柱亲和层析并进行SDS-PAGE检测,得到单一的蛋白条带。对重组酶CPG进行酶学性质研究,重组酶CPG在40 ℃时酶活最高,在20 ℃~50 ℃时具有较好的热稳定性;在pH值为3.5时酶活最高,pH值在3.0~5.0时具有较好的稳定性;Na+对重组酶CPG具有激活作用,而Cu2+具有较强的抑制作用。
3.2 将胃蛋白酶和丝氨酸羧肽酶CPG复配后应用于大豆蛋白脱苦,其水解度比仅使用胃蛋白酶时高9.55%,水解液中疏水性氨基酸(Ala、Val、Met、Ile、Leu、Tyr和Phe)含量明显增加,鲜味氨基酸(Glu)含量也明显增加,不仅降低了水解液的苦味,还明显增加了鲜味。因此重组酶CPG能够改善苦味,在食品工业上拥有较好的应用前景。
