复配涂封剂对低温胁迫下橡胶苗光合荧光特性的影响
2022-02-15程琳琳安锋谢贵水王纪坤王立丰张希财
程琳琳 安锋 谢贵水 王纪坤 王立丰 张希财

摘要:以橡胶树热研7-33-97芽接苗为材料,比较低温胁迫下市售、复配割面涂封剂处理对叶片叶绿素含量、光合作用及光化学效率的影响,以期为橡胶树新型割面涂封剂研发及抗寒机制研究提供理论依据。结果表明,低温胁迫导致叶片叶绿素含量、净光合速率(Pn)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)、PSⅡ最大光化学效率(Fv/Fm)下降,实际光化学效率(ΦPSⅡ)水平先升高后下降;低温胁迫1 d,以25 mg/L脱落酸、0.05 g/L壳聚糖、8.0 g/L CaCl2为主成分的复配割面涂封剂处理比市售割面涂封剂处理及对照的橡胶树叶片的ΦPSⅡ分别高出37.90%、80.29%,差异达到极显著水平(P<0.01);低温胁迫4 d,复配涂封剂处理比市售割面涂封剂处理及对照的橡胶树叶片的叶绿素a含量、叶绿素b含量和叶绿素总量分别高出23.49%、21.78%、23.07%和29.91%、26.04%、28.96%,差异均达到极显著水平(P<0.01);低温胁迫7 d,复配涂封剂处理比市售割面涂封剂处理及对照的橡胶树叶片的Pn、Gs、Ci、Tr、Fv/Fm高出74.53%、847.41%、90.94%、1 617.80%、6.57%和145.39%、7 700.23%、229.02%、18 009.72%、14.46%,差异达到显著或极显著水平。综合分析可知,复配割面涂封剂能够缓解由低温导致的橡胶树芽接苗叶片叶绿素含量、Pn、Gs、Ci、Tr、Fv/Fm及ΦPSⅡ的下降幅度,可以在一定程度上避免低温胁迫对光合系统的伤害,维持叶片光合能力及PSⅡ反应中心活性,有效缓解低温对橡胶树芽接苗的胁迫,提高其抗寒性。
关键词:复配割面涂封剂;橡胶苗;低温胁迫;光合作用;叶绿素荧光参数;抗寒性
中图分类号: S794.101 文献标志码: A
文章编号:1002-1302(2022)02-0102-06
收稿日期:2021-04-22
基金项目:国家天然橡胶产业技术体系(编号:CARS-33-ZP1);海南省重点研发计划(编号:ZDYF2020072)。
作者简介:程琳琳(1981—),女,河南太康人,硕士,助理研究员,主要从事橡胶树抗逆栽培生理生化研究。E-mail:chenglinlin2004@163.com。
通信作者:安 锋,博士,研究员,主要从事橡胶树栽培生理生态研究。E-mail:an-f@163.com。
巴西橡胶树(下面简称为橡胶树)原产于巴西亚马逊河流域,热带雨林气候造就了其喜高温、高湿、静风、土壤肥沃环境的生长习性[1]。虽然,我国成功实现了橡胶树北缘栽培,但位于热带北缘和南亚热带是我国植胶区的地域特点,寒害仍是橡胶树栽培不可避免的主要自然灾害之一。橡胶树对低温虽有一定耐受力,但是当环境温度降至其所能忍受的温度以下或低温积累超出它能承受的范围,就会产生寒害。25~30 ℃是橡胶树光合作用最适温度,相对湿度大于80%为最适宜湿度,在10 ℃以下橡胶树光合作用停止[1]。橡胶树同其他高等绿色植物一样,年总量90%~95%的有机质(以碳水化合物为主)由光合作用合成,光合作用生成的蔗糖即为合成天然橡胶(聚异戊二烯)的原料[2]。外界低温造就了光合系统的敏感性[3],一旦气温降至光合作用的临界点,橡胶树光合作用就会停止,随着低温延续,树体的呼吸作用消耗掉储备物质(主要为糖和淀粉等),而新的光合作用物质无法合成,其耐寒力被削弱,寒害就不可避免。因此,如何降低寒害对橡胶树的危害,对橡胶树的生长、发育、产量提高具有重要的意义。
脱落酸(ABA)作为一种重要的植物激素,在植物的生理生化过程中的作用不言而喻,在低温逆境中也是重要的信号及维持细胞结构的因子[4]。田礼欣等研究发现,外源施用适当浓度的ABA可以有利于玉米低温胁迫下叶绿素合成,缓解低温下幼苗叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、光系统Ⅱ(PSⅡ)的最大光化学效率(Fv/Fm)的下降,保护了低温胁迫中的光合系统,有效抑制低温胁迫对幼苗的伤害,使得幼苗耐冷性增强[5]。黄宇等用外源脱落酸叶面喷施雷公藤幼苗,发现由于脱落酸的施用,低温胁迫下雷公藤叶片Pn、Tr、Gs和胞间二氧化碳浓度(Ci)下降幅度得到有效减缓,PSⅡ的Fv/Fm、实际光化学量子产量(ΦPSⅡ)叶片光合、光合电子传递能力得到一定程度的维持,最终雷公藤的抗寒性得到了提高[6]。作为天然的碱性多糖,壳聚糖具有抑菌、无毒、生物相容性、保湿等诸多优点[7]。外源壳聚糖喷施剑叶全展时期的水稻叶片,可以缓解低温胁迫下水稻叶片叶绿素含量的降低,减缓光系统Ⅰ(PSⅠ)和PSⅡ电子传递活性、Fv/Fm、光化学淬灭(qP)下降的幅度,叶片光化学效率得到了维持,水稻抵御低温胁迫的能力得以提高[8]。吴琼等研究发现,壳聚糖拌种和叶面喷施后,玉米叶片的SPAD值、净光合速率、蒸腾速率、气孔导度和胞间二氧化碳浓度均增加,产量也有所增加[9]。钙是植物非胁迫及胁迫条件下生长、发育必需的基本元素,因此,它具有稳定膜系统、生理和发育过程第二信使的双重作用[10]。Ca2+是叶绿体的组成及功能成分,大多数钙被认为与类囊体膜和类囊体腔内的大分子有关[11],外源Ca2+可以缓解植物非生物胁迫下的光抑制,提高植物在不同胁迫下的光合作用[12-13]。Zhang等研究了外源氯化钙对夜间低温脅迫下番茄叶片光合系统活性、循环电子流、质子动力的影响,发现外源钙提高了夜间低温胁迫下的净光合速率、有效量子产率和光化学淬灭,最终减轻了夜间低温胁迫下的光抑制[14]。合适浓度的CaCl2可以有效减缓低温胁迫下西瓜叶片净光合速率、PSⅡ最大光化学效率和叶绿素a含量的降幅,缓解低温胁迫对西瓜幼苗的伤害[15]。
目前,市售橡胶树割面涂封剂多以成膜剂、单一生长调节剂与各种植物油脂为主要成分,具有结合生长调节剂促进树体生长及利用油脂膜将橡胶树割面与外界环境隔离的特点,以被动防御来抵御外界环境的低温。防护膜一旦因大风、雨水等外力脱落,其防寒效果就会大打折扣。王纪坤等研究发现,25 mg/L的ABA预处理橡胶树芽接苗可以有效提高其抗寒性[16]。王立丰等研究认为,0.05 g/L壳聚糖在提高橡胶树芽接苗和萌条的防寒效果方面表现较好[17]。张希财等研究认为,适宜浓度的CaCl2溶液可以有效保护橡胶树枝干免受低温伤害[18]。有关脱落酸、壳聚糖、CaCl2混合使用对低温胁迫下橡胶树光合系统、叶绿素荧光参数机制的研究还未见报道。本试验以橡胶树热研7-33-97芽接苗为材料,结合前人的研究及试验筛选的结果[16-18],研究了ABA、壳聚糖、CaCl2混配的割面涂封剂对低温胁迫下橡胶树叶片叶绿素含量、光合参数及叶绿素荧光参数的影响,以期为新型割面涂封剂研发及橡胶树抗寒栽培机制研究提供依据。
1 材料与方法
1.1 试验材料
本试验所用材料为橡胶树热研7-33-97芽接大型苗(2018年8月芽接,高度为2.1~3.5 m,2019年8月从苗圃移植至育苗容器)。试验选取生长正常、生长势基本一致的植株。
1.2 试验方法
1.2.1 试验设计
T1:用市面上销售的橡胶树割面涂封剂涂抹橡胶树芽接苗,T2:用25 mg/L ABA+0.05 g/L壳聚糖+8.0 g/L CaCl2混合液与毛棕榈油混配后(即新型复配割面涂封剂,由中国热带农业科学院橡胶研究所油棕种质资源课题组研发)涂抹橡胶树芽接苗,以清水作为对照(CK)。将各涂封剂和清水分别涂抹在橡胶树植株已木质化的茎干部位,每个处理20株。自然条件下,2种试剂与清水涂抹2 d后,将所有植株移入人工气候室中,进行低温胁迫。控制人工气候室的条件:12 h光照、12 h黑暗,相对湿度80%;光照时段的光照度为 75~100 μmol/(m2·s),温度控制在(16±1) ℃;黑暗时段的温度控制为(4±1) ℃。低温胁迫1、4、7 d后分别选取植株相同部位的叶片,用便携式叶绿素荧光仪测定叶绿素荧光参数,用便携式光合仪测定叶片光合参数;选取植株相同部位的叶片(每个植株选2张,每次选取5个植株采样),测定叶绿素含量。
1.2.2 测定方法
叶片叶绿素荧光参数测定利用WALZ PALM-2500便携式叶绿素荧光仪。叶片光合参数测定利用LI-6400XT便携式光合仪。其中,光照强度设置为100 μmol/(m2·s),采用标准叶室,开放式通路,流速设定为500 μmol/s。叶绿素含量采用混合液法[19]测定。
1.3 数据处理
所有试验数据为3次重复的平均值,采用DPS软件对数据进行分析,采用OriginPro 2018绘图,并采用Duncan’s新复极差法进行多重比较。
2 结果与分析
2.1 不同处理对低温胁迫下橡胶树叶片叶绿素荧光参数的影响
Fv/Fm为植物最大光合效率,也叫光系统Ⅱ的最大光化学量子产量,它反映了植物的最大光合潜能。Fv/Fm的降低程度反映了光抑制的程度。从图1-A可以看出,不同试剂处理后,在低温胁迫过程中,植株叶片PSⅡ最大光化学效率呈逐渐下降的趋势,T2处理的植株叶片PSⅡ最大光化学效率显著高于T1处理和CK(P<0.05)。在低温胁迫过程中,T2处理1、4、7 d后植株叶片PSⅡ最大光化学效率分别比T1处理和CK高出3.81%、4.50%、6.57%和4.57%、7.27%、14.46%,差异均达到显著水平。结果表明,与T1处理和CK相比,T2处理的植株在低温胁迫下保持了较高的光合潜能和光能利用率。
ΦPSⅡ表示植物實际光化学效率,它反映光系统Ⅱ的电子传递量子产率与光合电子传递速度。从图1-B可以看出,不同试剂处理后,在低温胁迫过程中,随着低温胁迫时间的延长,植株叶片实际光化学效率(ΦPSⅡ)呈先升高后下降的趋势,T2处理的植株叶片实际光化学效率明显高于T1处理和CK。在低温胁迫1、4、7 d后,T2处理的植株叶片ΦPSⅡ分别比T1处理和CK高出37.90%、5.43%、8.83%和80.29%、8.80%、20.77%,差异均达到显著或极显著水平。结果表明,与T1处理和CK相比,T2处理的植株在低温胁迫下保持了较高的光合电子传递速度和光化学效率。
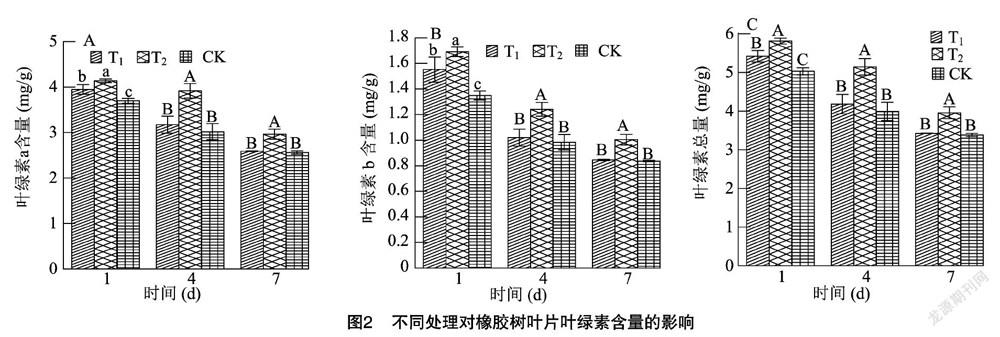
2.2 不同处理对低温胁迫下橡胶树叶片叶绿素含量的影响
从图2可以看出,不同试剂处理后,在低温胁迫的过程中,植株叶片叶绿素a含量、叶绿素b含量、叶绿素总量均呈逐渐下降的趋势,T2处理植株叶片的叶绿素a含量、叶绿素b含量、叶绿素总量明显高于T1和CK。在低温胁迫1、4、7 d后,T2处理的植株叶片叶绿素a含量分别比T1和CK高出4.81%、23.49%、14.44%和11.89%、29.91%、15.76%;T2处理的叶绿素b含量分别比T1处理和CK高出9.04%、21.78%、18.79%和25.34%、26.04%、20.05%;T2处理的叶绿素总量分别比T1处理和CK高出7.26%、23.07%、15.51%和15.48%、28.96%、16.81%,差异达到了显著或极显著水平。以上结果表明,与T1处理和CK相比,T2处理可以有效缓解低温胁迫下橡胶树叶片叶绿素含量的下降。
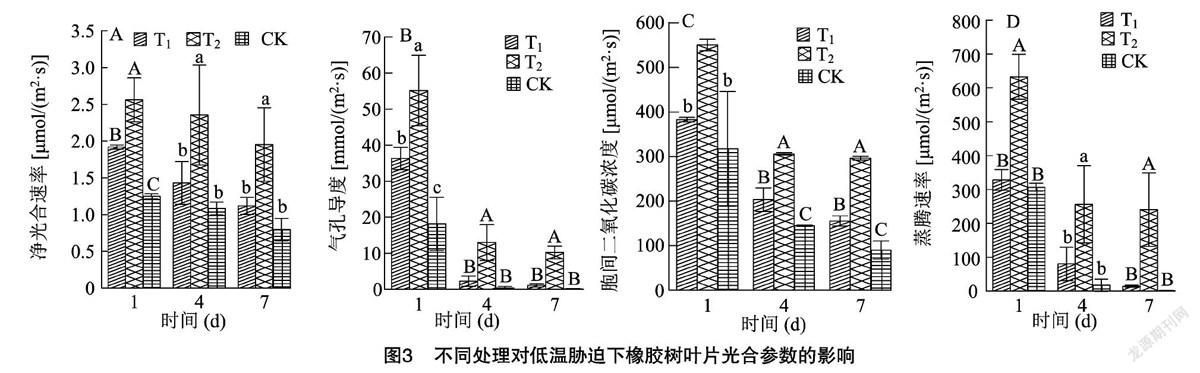
2.3 不同处理对低温胁迫下橡胶树叶片光合参数的影响
在低温胁迫过程中,随着时间的延长,叶绿体受损伤,叶片的光合性能也相应受到影响。从图 3-A 可以看出,不同试剂处理后,随着低温胁迫时间的延长,植株叶片净光合速率呈逐渐下降的趋势。在低温胁迫1、4、7 d后,T2处理的植株叶片净光合速率分别比T1处理和CK高出33.50%、64.56%、74.53%和105.00%、117.43%、145.39%,差异均达到了显著或极显著水平。以上结果表明,T2处理可以有效减缓低温胁迫下橡胶树叶片净光合速率的下降。
从图3-B可以看出,不同试剂处理后,随着低温时间的持续,植株叶片气孔导度呈逐渐下降的趋势,但是不同处理下降的幅度存在差异。在低温胁迫1、4、7 d后,T2处理的植株叶片气孔导度分别比T1处理和CK高出51.98%、470.91%、847.41%和203.61%、3 276.53%、7 700.23%,差异均达到显著或极显著水平。结果表明,与T1处理和CK相比,T2处理可以有效减缓低温胁迫下橡胶树叶片气孔导度的下降。
光合速率较大时,光合作用相应需求较大量的CO2,CO2较多地存储于叶片细胞间,所以胞间CO2浓度高。从图3-C可以看出,随着低温胁迫时间的延长,植株叶片胞间二氧化碳浓度呈逐渐下降的趋势,但是T2处理对胞间CO2浓度影响较大,T2处理的植株叶片胞间二氧化碳浓度分别比T1处理和CK高出43.74%、50.64%、90.94%和73.32%、111.62%、229.02%,差异均达到显著或极显著水平。以上结果表明,T2处理可以在一定程度上有效维持低温胁迫下橡胶树叶片对胞间CO2的需求量,减缓胞间二氧化碳浓度的降低。
植物吸收水分的能力在一定程度上决定了蒸腾作用的强弱,植株蒸腾能力的强弱体现于蒸腾速率。低温逆境致使蒸腾作用减弱,从图3-D可以看出,不同试剂处理后,低温胁迫的过程中,植株叶片Tr呈逐渐下降的趋势。蒸腾速率与气孔导度的变化趋势基本一致,不同处理植株的蒸腾速率差异显著或极显著。随着低温逆境时间的延伸,与T1处理和CK相比,T2处理有效减缓了植株叶片Tr的下降,分别比T1处理和CK高出92.71%、218.87%、1 617.80% 和106.41%、1 397.98%、18 009.72%,差异均达到显著或极显著水平。以上结果表明,T2处理可以有效减缓低温胁迫下橡胶树叶片Tr的降低。
3 结论与讨论
植物各种活动所需的有机物质与能量来源于光合作用。对于橡胶树而言,它的总生长量和经济产量(天然橡胶)的来源均是蔗糖,而蔗糖的合成能力由光合作用的能力来决定[2]。光合作用对外界环境变化比较敏感[20],一旦植物受到低温逆境的胁迫,光化学反应不仅受抑制,相关酶的活性与CO2固定、同化速率也受影响,导致植物受伤害[21]。低温胁迫对植物光合作用最直接的影响就是Pn下降,气孔和非气孔2种因素导致光合速率下降[22]。气孔因素中,Pn、Gs与Ci变化趋势一致,即低温使得氣孔关闭,气孔导度降低,从而进一步影响植物光合速率(Pn)与蒸腾速率(Tr)[23]。本研究中,不同处理的低温胁迫造成了橡胶树芽接苗叶片的Pn、Gs下降的同时,Ci也相应下降,说明橡胶树芽接苗叶片Pn的下降是由低温胁迫下气孔限制因素而导致的光合作用受抑制。复配涂封剂的使用,显著降低了低温胁迫下橡胶树芽接苗叶片Pn、Gs、Ci、Tr下降的幅度, 缓解了叶片光合作用受抑制的程度。低温胁迫发生后,植物膜系统就会受到损伤,随之,叶绿体膜受损伤导致叶绿素含量的下降。作为重要的光合色素,叶绿素的合成一旦受到抑制或因低温导致分解[24],植株的光能利用与转化能力就会降低[25],光合速率和叶片最大光合效率(Fv/Fm)随之下降[26]。类囊体膜流动性随低温胁迫时间的延长而逐渐降低,导致其上的电子传递链和光合磷酸化相关组分受影响,PSⅡ反应中心受损伤,其光能转换和电子传递速率下降或受抑制,即实际光化学效率下降[27]。本研究表明,低温胁迫破坏了叶绿体质膜结构,抑制了叶片的光合作用,导致橡胶树芽接苗叶绿素a含量、叶绿素b含量和叶绿素总量均持续下降,PSⅡ反应中心受到伤害,所以叶片最大光化学效率发生了下降。最初,橡胶树对低温具有一定的适应性,然而,随着低温胁迫的加剧和PSⅡ反应中心的受损,实际光化学效率最终降低,所以实际光化学效率水平呈现先升高后降低的趋势。但是,T2处理可以有效减缓低温胁迫对叶绿体质膜的伤害,在低温胁迫期间对PSⅡ反应中心提供了一定的保护,减缓低温诱导对橡胶树芽接苗的光抑制作用和电子传递速率的下降程度。
ABA在逆境环境下会及时清除氧自由基,减少质膜过氧化作用,从而对细胞结构维持起着重要作用。ABA可以防止叶绿体受低温胁迫的破坏,保证光合色素的正常合成和光合系统有条不紊地运行[28-29]。ABA可以有效减缓低温胁迫下薄皮甜瓜净光合速率、叶绿素含量和最大光化学效率的下降[30]。刘旭梅等研究发现,外施ABA可以有效减缓西洋杜鹃花叶片Gs、Ci、Pn、Tr的下降,提高其抗寒性[31]。伍宝朵等研究发现,对胡椒扦插苗叶片喷施外源ABA,可以有效缓解其Fv/Fm的下降,胡椒的光合系统受到一定程度的保护,从而提高胡椒的耐冻性[32]。壳聚糖是甲壳素脱乙酰基的产物,用它处理作物种子,可以提高叶绿素含量[33],提高种子萌发后叶片的Pn、Tr、Gs和Ci[34];减缓逆境下植物叶绿素含量、Pn、Gs、ΦPSⅡ的下降[8,35-36]。偶联胞外信息与胞内生理生化反应,从而引起植物对外界环境的响应,这是钙调节植物抵御外界胁迫能力的主要途径[37-39]。李烨等研究发现,外源喷施CaCl2可以提高低温胁迫下茄子幼苗叶绿素含量和光合速率,有效缓解茄子幼苗的冷害[40]。Zhang等研究发现,外源CaCl2预处理可以有效缓解夜间低温对番茄叶片造成的PSⅡ净光合速率、有效量子产率的下降,减缓夜间低温对番茄叶片的光抑制[14]。本研究结合ABA、壳聚糖及CaCl2对植物逆境下光合系统的保护作用,复配这3种成分为割面涂封剂。复配割面涂封剂可以有效缓解低温胁迫对橡胶树芽接苗的光抑制作用,提高其光能转化率,使其维持较高的叶绿素荧光产额,保证橡胶树芽接苗在低温胁迫下光合作用的进行,与前人利用外源ABA[28-32]、壳聚糖[8,33-36]及CaCl2[14,37-40]缓解低温胁迫对其他植物光合作用影响的研究结果相一致。
综上所述,复配割面涂封剂的使用,可以使低温胁迫下橡胶苗保持较高的光化学效率,减缓叶片叶绿素含量的下降,一定程度上避免了低温对PSⅡ反应中心的伤害,抑制低温胁迫下Pn、Gs、Ci、Tr等光合参数的下降,进而缓解了低温胁迫对橡胶苗的伤害,有效提高橡胶苗的耐寒性。这对橡胶树新型割面涂封剂的研发及抗寒机制探索具有重要的参考意义。
参考文献:
[1]何 康,黄宗道. 热带北缘橡胶树栽培[M]. 广州:广东科技出版社,1987.
[2]于俊红,黄绵佳,田维敏. 巴西橡胶树橡胶生物合成调控的研究进展[J]. 安徽农学通报,2007,13(12):38-40,153.
[3]Grzybowski M,Adamczyk J,Jończyk M,et al. Increased photosensitivity at early growth as a possible mechanism of maize adaptation to cold springs[J]. Journal of Experimental Botany,2019,70(10):2887-2904.
[4]Bravo L A,Zúiga G E,Alberdi M,et al. The role of ABA in freezing tolerance and cold acclimation in barley[J]. Physiologia Plantarum,1998,103(1):17-23.
[5]田礼欣,杨 晔,左师宇,等. 脱落酸对低温胁迫下玉米幼苗生长和光合特性的影响[J]. 作物杂志,2018,34(6):76-82.
[6]黄 宇,林智勇,荣俊冬,等. 脱落酸对低温下雷公藤幼苗光合作用及叶绿素荧光的影响[J]. 应用生态学报,2011,22(12):3150-3156.
[7]王继业,张明宇,霍 倩,等. 壳聚糖的性能及应用[J]. 纺织科学与工程学报,2020,37(3):110-116.
[8]孙 磊,陈国祥,吕川根,等. 壳聚糖对低温处理下水稻剑叶光合特性的影响[J]. 南京师大学报(自然科学版),2010,33(4):75-79.
[9]吴 琼,郑殿峰,冯乃杰,等. 壳聚糖不同处理方式对玉米光合特性及产量的影响[J]. 安徽农学通报,2020,26(10):18-22.
[10]Thor K. Calcium-nutrient and messenger[J]. Frontiers in Plant Science,2019,10:440.
[11]Bayer R G,Stael S,Rocha A G,et al. Chloroplast-localized protein kinases:a step forward towards a complete inventory[J]. Journal of Experimental Botany,2012,63(4):1713-1723.
[12]Xu C B,Li X M,Zhang L H.The effect of calcium chloride on growth,photosynthesis,and antioxidant responses of Zoysia japonica under drought conditions[J]. PLoS One,2013,8(7):e68214.
[13]齐明芳,刘玉凤,周龙发,等. 钙对亚高温下番茄幼苗叶片光合作用的调控作用[J]. 中国农业科学,2011,44(3):531-537.
[14]Zhang G X,Liu Y F,Ni Y,et al. Exogenous calcium alleviates low night temperature stress on the photosynthetic apparatus of tomato leaves[J]. PLoS One,2014,9(5):e97322.
[15]韩晓村. 缓解西瓜幼苗期冷害物质的筛选及处理技术研究[D]. 杨凌:西北农林科技大学,2018.
[16]王纪坤,张盈盈,王立丰,等. 脱落酸预处理提高橡胶树芽接苗期抗寒性的研究[J]. 热带农业工程,2016,40(3):16-20.
[17]王立丰,王纪坤,安 锋,等. 一种橡胶树抗寒剂及其应用:CN106900728B[P]. 2019-02-22.
[18]张希财,林位夫,安 峰,等. 一种橡胶树枝干防寒方法:CN101697699A[P]. 2010-04-28.
[19]陳福明,陈顺伟. 混合液法测定叶绿素含量的研究[J]. 浙江林业科技,1984,4(1):19-23,36.
[20]许大全. 光合速率、光合效率与作物产量[J]. 生物学通报,1999,34(8):8-10.
[21]李新国,毕玉平,赵世杰,等. 短时低温胁迫对甜椒叶绿体超微结构和光系统的影响[J]. 中国农业科学,2005,38(6):1226-1231.
[22]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology,1982,33(1):317-345.
[23]Ikkonen E N,Shibaeva T G,Sysoeva M I,et al. Stomatal conductance in Cucumis sativus upon short-term and long-term exposures to low temperatures[J]. Russian Journal of Plant Physiology,2012,59(5):696-699.
[24]项洪涛,郑殿峰,何 宁,等. 植物对低温胁迫的生理响应及外源脱落酸缓解胁迫效应的研究进展[J]. 草业学报,2021,30(1):208-219.
[25]郑春芳,陈 威,刘伟成,等. 低温胁迫后红树植物秋茄幼苗光合特性及蔗糖代谢的恢复机制[J]. 生态学杂志,2020,39(12):4048-4056.
[26]Li X N,Pu H C,Liu F L,et al. Winter wheat photosynthesis and grain yield responses to spring freeze[J]. Agronomy Journal,2015,107(3):1002-1010.
[27]陶宏征,赵昶灵,李唯奇.植物对低温的光合响应[J]. 中国生物化学与分子生物学报,2012,28(6):501-508.
[28]李 进,张军高,刘梦丽,等. 4种外源物质对低温胁迫下棉花(Gossypium hirsutum)幼苗的缓解效应分析[J]. 分子植物育种,2019,17(17):5792-5800.
[29]曾继娟,种培芳,朱 强. 外源脱落酸对荒漠植物红砂幼苗光合及叶绿素荧光特性的影响[J]. 江苏农业科学,2020,48(7):136-141.
[30]王 乾. 喷施外源物质对薄皮甜瓜苗期低温胁迫的缓解作用研究[D]. 沈阳:沈阳农业大学,2020:17-18.
[31]刘旭梅,赵 冰. 不同浓度外源ABA对西洋杜鹃花抗寒性的影响[C]//中国园艺学会. 中国观赏园艺研究进展2018.哈尔滨,2018:225-232.
[32]伍宝朵,崔 鑫,胡丽松,等. 叶面喷施外源ABA对胡椒抗寒生理生化及ABA信号转导相关基因的影响[J]. 南方农业学报,2020,51(11):2764-2772.
[33]张伟伟,王 玺,曹 宏,等. 壳聚糖包衣对水分胁迫下玉米种子萌发和幼苗生长的影响[J]. 玉米科学,2011,19(3):98-101.
[34]卢洁春. 生物源植物生长物质对大豆生长发育及产量的影响[D]. 大庆:黑龙江八一农垦大学,2019.
[35]张志刚,尚庆茂.水杨酸、壳聚糖对盐胁迫下黄瓜叶片光合参数的调节作用[J]. 西北农业学报,2010,19(3):174-178.
[36]王 聪,杨恒山,董永义,等. 外源壳聚糖对NaCl胁迫下菜用大豆光合作用及荧光特性的影响[J]. 西北植物学报,2015,35(6):1198-1205.
[37]郝正剛,王志恒,魏玉清,等. 外源钙镉处理对甜高粱幼苗叶片光合作用的影响[J]. 江苏农业科学,2019,47(19):75-80.
[38]苗永美,简 兴,钱立生,等. 氯化钙和水杨酸对弱光胁迫下甜瓜幼苗生长及光合的影响[J]. 安徽科技学院学报,2018,32(1):25-31.
[39]夏 蕴,谷 巍,巢建国,等. 外源氯化钙对高温胁迫下茅苍术生长生理指标及叶片关键酶基因表达的影响[J]. 南方农业学报,2020,51(2):267-274.
[40]李 烨,谢立波,陈永琴,等. 外源氯化钙和脱落酸处理对茄子低温胁迫相关指标的影响[J]. 北方园艺,2012(7):22-25.
