拟藁本属及近缘物种中国分类群花粉形态与质体基因组系统发育研究
2022-02-13李子宣周颂东何兴金
李子宣,周颂东,何兴金
(四川大学 生命科学学院,生物资源与生态环境教育部重点实验室,成都 610065)
拟藁本属(LigusticopsisLeute)隶属伞形科(Apiaceae)芹亚科(Apioideae),由Leute于1969年建立[1],最初包含14个报道于中国的物种,Leute以萼齿显著为属间形态诊断特征将拟藁本属与藁本属(LigusticumL.)相区分,并以玉龙拟藁本(LigusticopsisrechingeranaLeute)作为本属模式物种。由于Leute在之后1970年[2]所修订的一些藁本属物种中也具有显著的萼齿,致使拟藁本属的分类学地位自建立后一直备受争议,最大的争议在于该属独立地位以及属间界限的划分。《中国植物志》、《中国藁本属修订》以及《FloraofChina》均不承认拟藁本属的独立地位,并将属下物种归并入藁本属以及蛇床属(CnidiumCusson)中[3-5]。
以Pimenov为代表的多位俄罗斯植物学家则承认拟藁本属的独立地位,并依据果实解剖学形态多次对拟藁本属和近缘属进行分类修订和形态学分类边界澄清。这部分学者认为在拟藁本属下共有19个物种,其中有18种属于中国地区喜马拉雅-横断山脉分布,另一种分布于喜马拉雅山脉西侧(中国之外),且该属最重要的分类学鉴别特征为:分果爿强烈背面压扁,背部棱丝状或龙骨状突起、边棱呈翅状,棱槽内及合生面的长油管多数(非单生)[6-9]。
近年来基于核基因片段(如ITS序列)、质体基因片段以及血清学证据的系统发育研究已经证实《FloraofChina》所划分的广义藁本属(Ligusticumsensulato)以及Leute所划分的拟藁本属均为复系类群[10-13],而经Pimenov等分类修订后的部分拟藁本属物种(包括模式物种)在质体基因组系统发育树中位于亮蛇床部族(Selineae)并形成了高支持率的单系分支[14]。尽管这部分物种在基于ITS所构建的系统发育树中与近缘属(如PachypleurumLedeb.,CortiellaC. Norman,OreocomeEdgew.)相互嵌套在一起形成复系分支,但Pimenov等通过果实解剖学明确了这些属之间的形态分类界限,即承认这些近缘属各自的独立属地位[6, 7, 15],Li等通过质体全基因组系统发育结合核ITS基因以及果实解剖研究,结果表明拟藁本属是独立的且宜从广义藁本属中分离出来独立建属[16]。这些分类历史表明将拟藁本属直接并入藁本属的分类处理需谨慎对待并且需要更多的形态学和分子系统学证据。
虽然国内外学者对拟藁本属进行了较为全面的分子和果实解剖研究,但迄今为止,国内外学者并未单独对拟藁本属的花粉形态学进行系统的研究,主要的报道均来自于藁本属孢粉学研究中所涉及到的少数曾处理在拟藁本属中的物种,且这些研究同样不支持拟藁本属的独立[17-19]。花粉形态特征由于其在属间和物种间的稳定性,历来为分类学家重视和关注[20-22],本研究共报道拟藁本属8种及近缘类群2种(共10种)的花粉形态,8种为Leute(1969)所处理的拟藁本属物种,2种为Pimenov建议转入拟藁本属的物种。通过扫描电子显微镜和普通光学显微镜对拟藁本属及近缘类群10种植物的花粉进行观察比较,旨在为拟藁本属的系统分类地位提供花粉形态学方面的证据,同时,结合对应物种及近缘类群质体基因组所构建的系统发育树,为探究拟藁本属的独立属地位和演化提供花粉形态学佐证资料。
1 材料和方法
1.1 材 料
用于花粉形态研究的拟藁本属8种及近缘类群2种(共10种,表1)物种,分别为:1.归叶拟藁本[Ligusticopsisangelicifolia(Franch.) Leute]、2. 短片拟藁本[Ligusticopsisbrachyloba(Franch.) Leute]、3. 细苞拟藁本[Ligusticopsiscapillacea(H. Wolff) Leute]、4. 美脉拟藁本[Ligusticopsisintegrifolia(H. Wolff) Leute]、5. 玉龙拟藁本(LigusticopsisrechingeranaLeutoe)、6. 细裂拟藁本[Ligusticopsismodesta(Diels) Leute][23]、7. 细叶亮蛇床[Selinumwallichianum(de Candolle) Raizada & H. O. Saxena]、8. 毛藁本[Ligusticumhispidum(Franch.) Wolff]、9. 尖叶拟藁本[Ligusticopsisacuminata(Franch.) Leute]、10. 蕨叶拟藁本[Ligusticopsispteridophylla(Franch.) Leute]。野外观察并采集具有饱满花药的伞形花序,置于分子材料袋后于硅胶中快速吸水干燥,而后室温条件下干燥保存,对应的处理方法见后文。
为获得较客观的系统发育关系,参考前人研究中拟藁本属原始建属物种(1969年)在系统发育树中的分布情况,选取与其近缘的来自亮蛇床部族(17个,表1,No. 1~8、11~19)、滇藁本分支(HymenidiumClade, 6个, 表1,No. 9~10、20~23)以及丝瓣芹分支(AcronemaClade, 1个, 表1, No. 24)的24个物种(包含用于花粉形态研究的10个物种)以探究其系统发育关系,类群对应的质体基因组序列来自GenBank下载序列以及本团队先前测序、组装、注释后上传于GenBank的公开数据。凭证标本保存于四川大学标本馆(SZ),凭证材料保存于四川大学生命科学学院植物系统与分子进化生物学实验室。本研究中使用“Ligusticopsis”属名前缀的物种均为 Leute于1969年首次划分在拟藁本属下的物种,并使用“拟藁本”作为各自中文译名的后缀,其余物种的学名及名称参考《FloraofChina》。

表1 材料来源及GenBank登录号
1.2 方 法
1.2.1 芹亚科拟藁本属及近缘类群10种植物花粉的扫描电镜观察在解剖镜下挑选拟藁本属及近缘类群10种植物饱满的花药,将其粘贴于电镜样品台上的双面导电胶,并用解剖针将花药捣碎使花粉平铺于导电胶平面,随后镀膜喷金,于JSM-7500F型扫描电镜下观察花粉粒, 并拍摄保存有代表性的花粉粒整体观、极面观、赤道面观和外壁纹饰等特征。
1.2.2 芹亚科拟藁本属及近缘类群10种植物花粉的光学显微镜观察使用Erdtman[24]的醋酸酐分解法处理花粉,并根据前人所报道的伞形科花粉资料[25, 26]进行参数调整,具体步骤为:(1)饱满花药于1 mL冰醋酸中浸没24 h,而后用玻璃棒充分碾碎,100目铜网过滤;(2)滤液加冰醋酸至5 mL,离心(5 000 r/min, 10 min)两次沉淀花粉,弃上层液;(3)加入浓硫酸与醋酸酐的混合液(体积比为1∶9,现用现配)5 mL,80 ℃水浴分解4 min;(4)分解后,离心(5 000 r/min, 10 min)弃上层液,加入4 mL蒸馏水清洗一次,离心(5 000 r/min, 10 min)并弃上层液;(5)滴入适量50%甘油保存,滴2~3滴1%的苯酚溶液防腐。取花粉悬浊液一滴制作装片,甘油胶封片,于光学显微镜下观察并拍照。花粉形态学数据测量于KaryoType[27]软件中进行,40倍光镜物镜镜头下每个物种随机选取40粒花粉测量。使用Excel软件统计花粉的极轴长(P)、赤道轴长(E),记录常见值和变化范围,并取常见值作为花粉大小数据,计算极轴长与赤道轴长的比值(P/E)。
1.2.3 基于质体CDS序列的系统发育构建使用24个来自于伞形科芹亚科的质体序列系统发育重建,材料来源见表1,位于丝瓣芹分支的云南藏香芹[Meeboldiayunnanensis(H. Wolff) Constance & F. T. Pu] 作为外类群:(1)使用Geneious v9.0.2和PhyloSuite v1.2.2. 软件对质体基因编码区(CDS)序列进行提取与连接[28-29],随后使用MAFFT v7.402软件对连接好的序列进行对齐比对,使用ModelFinder软件确定序列数据集的最适建树模型为GTR+G模型[30-31],(2)RAxML v7.2.8软件用于进行最大似然(maximum likelihood, ML)分析,于软件中进行自举检验1000次[32],(3)贝叶斯推断法(Bayesian inference, BI)于MrBayes v.3.2.7a[33]软件中进行,运行1 × 107代马尔可夫-蒙特卡尔罗(Markov Chain Monte Carlo, MCMC),其中每1000代保存一个树文件,前20%的树设置为Burn-in,余80%的树用于构建一致树(Consensus tree)。
1.2.4 专业术语描述花粉形态术语参考《孢粉学手册》、许新锐、孟德玉、陈薇薇[24-26, 34]等的研究;系统发育树的主要部族分支名参考Downie[35]、苟巍等[36]的研究。
2 结果与分析
2.1 拟藁本属及近缘类群10种植物花粉的形态特征
根据光学显微镜和扫描电镜的观察结果(图版Ⅰ、图版Ⅱ、图版Ⅲ,表2),花粉的极轴长为27.05~38.87 μm,赤道轴长为13.37~18.57 μm,极轴长与赤道轴长的比值P/E为1.89~2.31,花粉大小变化范围25.45~41.70 × 11.29~20.52 μm;10种花粉均具三孔沟,沟长且细,孔类型为边孔,其中尖叶拟藁本(图版Ⅱ,9a)、蕨叶拟藁本(图版Ⅱ,10a)、短片拟藁本(图版Ⅰ,2a)以及细叶亮蛇床(图版Ⅱ,7a)的孔边缘突起。光学显微镜和扫描电镜下的花粉形态,归叶拟藁本(表2、图版Ⅰ、图版Ⅲ,1)和细裂拟藁本(表2、图版Ⅱ、图版Ⅲ,6)为近矩圆-赤道微收缩形,尖叶拟藁本为近长椭圆形(表2、图版Ⅱ、图版Ⅲ,9)、蕨叶拟藁本为菱形(表2、图版Ⅱ、图版Ⅲ,10),其余6种皆近矩圆形,极面观除毛藁本(图版Ⅱ,8b)的花粉为近三瓣形外,其余9种花粉的极面观均为三裂圆形。在赤道面纹饰方面,蕨叶拟藁本为短棒条纹(图版Ⅱ,10c),尖叶拟藁本为凹凸粗条纹(图版Ⅱ,9c),其余8种的赤道面纹饰类型为条网纹和网纹。而在极面纹饰类型上,蕨叶拟藁本为条状纹饰(图版Ⅱ,10b),尖叶拟藁本为平滑穴网纹(图版Ⅱ,9b),其余8种均为有孔分布的网纹或条网纹纹饰。并首次报道了玉龙拟藁本(表2、图版Ⅰ、图版Ⅲ,5)和细裂拟藁本的花粉形态特征观察结果。
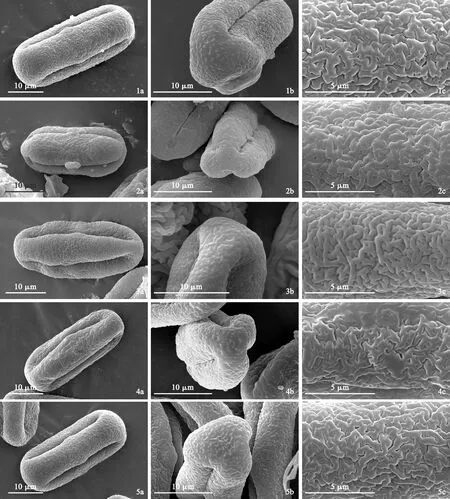
图版Ⅰ 扫描电镜下拟藁本属及近缘类群10种植物的花粉形态Ⅰ1. 归叶拟藁本;2. 短片拟藁本;3. 细苞拟藁本;4. 美脉拟藁本;5. 玉龙拟藁本;a. 赤道面观;b. 极面观;c. 赤道面纹饰Plate Ⅰ Pollen morphology of ten species from Ligusticopsis and its relatives under SEM Ⅰ1. Ligusticopsis angelicifolia; 2. L. brachyloba; 3. L. capillacea; 4. L. integrifolia; 5. L. rechingerana; a. Equatorial view; b. Polar view; c. Equatorial sculpture

图版Ⅱ 扫描电镜下拟藁本属及近缘类群10种植物的花粉形态Ⅱ6. 细裂拟藁本;7. 细叶亮蛇床;8. 毛藁本;9. 尖叶拟藁本;10. 蕨叶拟藁本;a. 赤道面观;b. 极面观;c. 赤道面纹饰Plate Ⅱ Pollen morphology of ten species from Ligusticopsis and its relatives under SEM Ⅱ6. Ligusticopsis modesta; 7. Selinum wallichianum; 8. Ligusticum hispidum; 9. Ligusticopsis acuminata; 10. Ligusticopsis pteridophylla; a. Equatorial view; b. Polar view; c. Equatorial sculpture
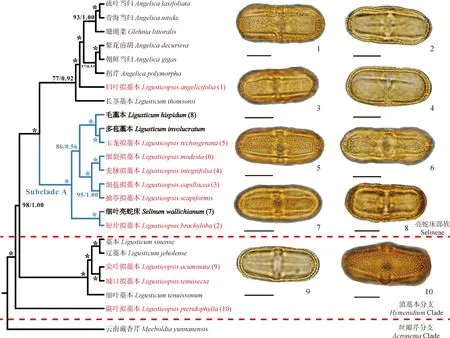
图版Ⅲ 光学显微镜下拟藁本属及近缘类群10种植物的花粉形态与基于质体基因组CDS序列构建的系统发育树1. 归叶拟藁本;2. 短片拟藁本;3. 细苞拟藁本;4. 美脉拟藁本;5. 玉龙拟藁本;6. 细裂拟藁本;7. 细叶亮蛇床;8. 毛藁本;9. 尖叶拟藁本;10. 蕨叶拟藁本;标尺为10 μm;系统发育树上节点前后的数字分别为BI-PP和ML-BS,*代表在两种分析方法中均为最大值;Leute (1969) 所划分的拟藁本属物种用红色标记,Pimenov等人后来划入拟藁本属的物种用粗体标记Plate Ⅲ Pollen morphology of ten species from Ligusticopsis and its relatives under LM and phylogenetic tree based on plastome CDS sequences1. Ligusticopsis angelicifolia; 2. Ligusticopsis brachyloba; 3. Ligusticopsis capillacea; 4. Ligusticopsis integrifolia; 5. Ligusticopsis rechingerana; 6. Ligusticopsis modesta; 7. Selinum wallichianum; 8. Ligusticum hispidum; 9. Ligusticopsis acuminata; 10. Ligusticopsis pteridophylla; Scale bar = 10 μm; The numbers before and after the nodes on the phylogenetic tree are BI-PP and ML-BS, respectively, (*) represents maximum support in both two analyses; Species named with Ligusticopsis from the treatments by Leute (1969) were marked in red, species later were placed into Ligusticopsis by Pimenov et al. were marked in bold
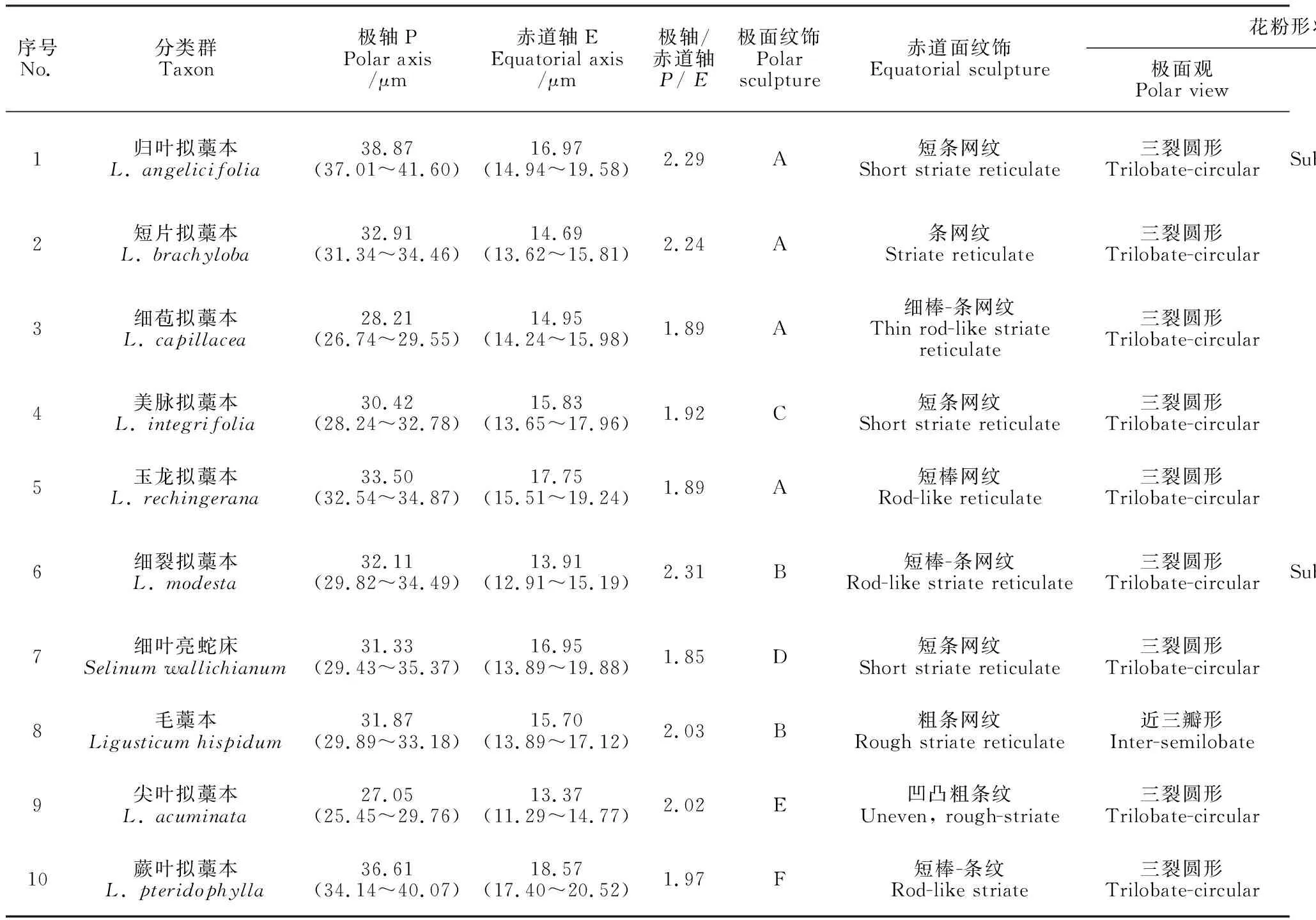
表2 拟藁本属及近缘类群10种植物的花粉形态
2.2 系统发育结果
基于两种方法所构建的质体系统发育树拥有一致的拓扑结构,见图版Ⅲ,而Leute(1969)原始处理为拟藁本属的10个物种呈现多系分布,其系统关系如下:归叶拟藁本位于亮蛇床部族分支,并与珊瑚菜以及5种当归属(AngelicaL.)物种近缘;尖叶拟藁本、城口拟藁本以及蕨叶拟藁本均位于滇藁本分支,前两者与其他3种藁本属物种结盟,而蕨叶拟藁本则单独分支并成为亮蛇床部族和滇藁本分支的姐妹物种;其余6个拟藁本属物种(包括该属模式物种玉龙拟藁本)与毛藁本、多苞藁本以及细叶亮蛇床在亮蛇床部族分支内形成具有强支持率的单系分支(Subclade A),在该分支内,美脉拟藁本与细裂拟藁本近缘并形成姐妹次级分支,细苞拟藁本与抽葶拟藁本近缘,多苞藁本、玉龙拟藁本与毛藁本结盟形成一个强支持率的次级分支且前两者有更近的亲缘关系,短片拟藁本与细叶亮蛇床结盟并形成强支持率的次级分支并形成上述6个物种的姐妹分支。
3 讨 论
3.1 系统学意义
分子系统发育研究结果与花粉形态及扫描电镜观察结果一致,本研究位于Subclade A内的7个物种(玉龙拟藁本, 细裂拟藁本, 美脉拟藁本, 细苞拟藁本、短片拟藁本、毛藁本、细叶亮蛇床)无论从光镜花粉形态(近矩圆形至赤道收缩形)、扫描电镜观察的纹饰(条网状或细孔网状纹饰)方面,还是从分子系统发育树来看,均表明该亚分支为一单系类群,且该部分物种在花粉形态上与系统发育树也有较好的契合,即支持将拟藁本属从广义藁本属中划出独立成属,位于亮蛇床部族内的Subclade A即代表了真正的拟藁本属,而位于该单系分支外的曾归于拟藁本属的物种(归叶拟藁本、城口拟藁本、尖叶拟藁本、蕨叶拟藁本)应从真正的拟藁本属中转出。同时,花粉形态上这7个物种为近矩圆形至赤道收缩形,属于伞形科中较为进化的花粉形态[18, 20, 26],且属下物种的花粉形态仍有向赤道收缩形进化的趋势;扫描电镜下纹饰为条网状纹饰(赤道面)以及具孔的条网状纹饰(极面),其网状-条网状纹饰的性状在属下较为稳定,是可用于分属的重要微形态证据。花粉形态学与基于质体基因组的系统发育研究结果的一致性和相互佐证,说明了花粉形态的分类学价值和系统学意义。
3.2 一些疑似拟藁本属物种的系统地位及分类问题
首先从本研究结果看:归叶拟藁本从花粉形态上为近矩圆-赤道收缩形花粉,而赤道收缩形花粉是伞形科中较为进化的花粉类型。单独从花粉形态特征看与前人报道的当归属花粉形态完全一致[26];另一方面,基于质体序列的分子系统发育结果表明,归叶拟藁本插入到当归属的分支中,且植株形态方面归叶拟藁本也与当归属有极高的相似性;在果实形态上,归叶拟藁本的果实背面压扁程度较低,其3条背部棱均凸起呈窄翅状,无萼齿宿存,这与真正的拟藁本属(即系统树上的Subclade A)果实所具有的“背棱丝状或龙骨状凸起、强烈背面压扁以及有萼齿宿存”形态相区别。因此,本研究支持将归叶拟藁本转入当归属[6, 37]。
其次,从分子系统发育关系来看,尖叶拟藁本以及城口拟藁本这两个种构成的单系分支插入到滇藁本属分支中并与藁本等物种近缘,即这两个种应该划入藁本属中而不属于真正的拟藁本属;从花粉形态上看尖叶拟藁本花粉为长椭圆形,扫描电镜下观察其纹饰为平滑穴网纹(极面)和凹凸粗条状纹饰(赤道面),与拟藁本属的近矩圆形花粉及条网状纹饰明显不同,而与王萍丽等[17]发表的藁本属的花粉形态及纹饰一致。因此,从花粉形态学以及分子系统发育分析结果的角度均支持将这两个种从拟藁本属中转出并纳入藁本属中。且从外部形态上看来[16],尖叶拟藁本和城口拟藁本的总苞片与小总苞片均为线形,植株根颈部无纤维状枯萎叶鞘覆盖,两个种的果实背棱以及侧棱均为宽翅状,无萼齿宿存,这些性状都与拟藁本属的性状相背但与藁本属性状相似[3, 7]。
第三,花粉形态和扫描电镜观察结果表明,细叶亮蛇床和毛藁本所具有的花粉形态和纹饰与真正的拟藁本属完全一致,从花粉形态学角度建议将细叶亮蛇床划入拟藁本属中,这一结果也得到了质体分子系统发育的强烈支持。从外部形态上看,这两个种的根颈部均覆盖纤维状枯萎叶鞘,总苞片和小总苞片均为羽状分裂或先端三裂,果实均存在宿存萼齿、强烈背面压扁、背部棱均丝状或龙骨状凸起[3, 6],其中毛藁本的伞幅虽然极度拉长,但两者的其他共有形态与真正的拟藁本属高度一致,进一步支持这2个种纳入拟藁本属的分类处理。
第四,本花粉形态研究结果表明蕨叶拟藁本的花粉形态为近长菱形型,属于伞形科较为原始的花粉类型[34],扫描电镜下纹饰为短棒条纹(赤道面)、条状纹饰(极面),与真正的拟藁本属的近矩圆-赤道收缩形花粉及条网状纹饰明显不同。而分子系统发育研究结果中,蕨叶拟藁本位于质体基因组系统发育分支树上较为原始的分支,为整个大分支(即滇藁本分支和亮蛇床部族)的基出分支,几乎与来自丝瓣芹分支的外类群云南藏香芹同为基出分支,而云南藏香芹的花粉也为较原始的菱形花粉[18]。这些结论说明蕨叶拟藁本宜从拟藁本属中划出去,且与藁本属也存在明显差异,这一结果与任婷等[14]的研究结果一致,蕨叶拟藁本的系统分类地位需要进一步研究。
3.3 似拟藁本属物种的演化趋势
本文的花粉形态学研究结果表明,从真正属于拟藁本属的6个种的花粉形态角度,其花粉形态均为近矩圆形至赤道收缩形、扫描电镜下为条网或细孔网状纹饰,为一较为自然的分类群,且与已报道的藁本属花粉形态不同,从花粉形态学角度支持Leute(1969)和Pimenov等关于将拟藁本属独立的观点[1, 6-7, 9]。结合分子系统发育研究结果,同样表明拟藁本属单独形成一个单系分支(Subclade A,图版Ⅲ),而且在拟藁本属下物种间的亲缘关系上来看,花粉形态和纹饰性状较为稳定;质体分子系统发育树也显示在亮蛇床部族内,5种拟藁本(Leute于1969年的源分类处理)、2种藁本(毛藁本与多苞藁本),共7个种组成了2个较为高级的姊妹类群关系,然后再与另外2个物种(细叶亮蛇床与短片拟藁本)构成真正的拟藁本属的分支,显示拟藁本属为一年轻的同一祖先分化的属。结果同样显示拟藁本属仍处于进化的阶段,其花粉形态的进化趋势为由近矩圆形(如短片拟藁本和细叶亮蛇床)的过渡态向赤道收缩形(如细裂拟藁本)的进化态[17]发展,这一点也与系统发育树相契合,且这与植株外部形态也相吻合,如小总苞片的进化趋势由线形与先端二裂或三裂并存(如短片拟藁本和细叶亮蛇床)向多回羽状分裂(如玉龙拟藁本、细苞拟藁本等)进行。花粉形态结合系统发育与植物地理区系及植株外部形态的演化趋势表明,拟藁本属可能是起源于泛东亚的祖先种,在向西及更高海拔的青藏高原迁移扩散过程中,于东亚至中国-喜马拉雅地区适应性快速辐射物种形成的结果。
作者贡献:李子宣开展了野外采样、花粉悬浊液制备、系统发育树构建、扫描电镜花粉形态观察以及文章撰写工作,周颂东提供了花粉制备的方法及以往参数参考,何兴金审阅了文章并在进化讨论部分提供了建设性意见。
