黄带犀猎蝽成虫对斜纹夜蛾幼虫的捕食功能反应
2021-12-20只佳增周劲松刘学敏张建春岳建伟
杜 浩, 只佳增, 周劲松, 刘学敏, 张建春, 岳建伟
云南省红河热带农业科学研究所,云南 河口 661300
黄带犀猎蝽SycanuscroceovittatusDohrn,又称中黄猎蝽,属半翅目Hemiptera猎蝽科Reduviidae,是农林业中一种重要的捕食性天敌。黄带犀猎蝽为不完全变态昆虫,分为卵、若虫和成虫3个虫态,若虫共5个龄期。黄带犀猎蝽在国外分布于缅甸和泰国,在国内分布于福建、广东、广西、云南等省(区)(王亚楠等,2020)。其猎物范围广泛,包括蚜虫、姬缘蜷、蝗虫、蟋蟀、甲虫、蠼螋、叶蜂、菜青虫、蚊和蜘蛛等(黄增和等,1991; Hoffmann,1934)。
斜纹夜蛾SpodopteralituraFabricius隶属鳞翅目Lepidoptera夜蛾科Noctuidae,俗称莲纹夜蛾、夜盗虫、夜老虎等(张根顺等,2003)。据统计,斜纹夜蛾的寄主植物有109科389种(包括变种),主要以幼虫取食植物叶片、蛀茎(果)等(罗凯等,2015)。长期以来,对斜纹夜蛾的治理主要以化学防治为主,但随着3R问题的日益突出,生物防治越来越引起植保工作者的关注(田耀加等,2011)。关于斜纹夜蛾的生物防控已有较多报道,包括自然天敌的利用、植物源提取物、性信息素、转基因技术和抗虫品种选育等(罗凯等,2015)。自然天敌的发掘和利用是生物防治中一个重要的方向,研究人员已相继研究了叉角厉蝽Cantheconideafurcellata(Wolff)(陈然等,2015)、红彩真猎蝽Harpactorfuscipes(Fabricius)(邓海滨等,2012)、拟环纹豹蛛Pardosapseudoannulata(Boesenberg et Strand)(田耀加等,2011)、蠋蝽Armachinensis(Fallou) (唐艺婷等,2020)对斜纹夜蛾的捕食能力。
笔者在前期香蕉病虫调查中常观察到黄带犀猎蝽成虫捕食斜纹夜蛾幼虫的现象,而关于黄带犀猎蝽对斜纹夜蛾的捕食情况还未见报道。为探明黄带犀猎蝽对斜纹夜蛾的捕食能力、寻找效应和干扰反应,在室内开展了黄带犀猎蝽成虫对2、3、4龄斜纹夜蛾幼虫的捕食功能反应试验,以期为斜纹夜蛾的天敌防控提供理论参考。
1 材料与方法
1.1 供试昆虫
黄带犀猎蝽成虫和斜纹夜蛾2龄幼虫均采自云南省红河热带农业科学研究所香蕉园。斜纹夜蛾3、4龄幼虫以新鲜香蕉叶续代饲养建立种群。以下试验均在室内自然条件下开展,试验时间为2020年9—10月。
1.2 试验方法
1.2.1 黄带犀猎蝽对斜纹夜蛾幼虫的捕食功能反应 试验在透明盒子中进行(盒子上开有数个小孔,长150 cm,宽15.0 cm,高8.5 cm),在盒子底部放入新鲜洗净香蕉叶供斜纹夜蛾幼虫取食,在盒子角落放一蘸水棉花团,供黄带犀猎蝽补充水分。斜纹夜蛾2、3、4龄幼虫均设置每盒3、6、9、12、15、18、21头7个梯度,分别放入1头饥饿处理24 h的黄带犀猎蝽,每处理重复4次,24 h后统计捕食量。
1.2.2 捕食者自身密度对捕食功能反应的影响 每个盒子中放入40头3龄斜纹夜蛾幼虫,分别放入1、2、3、4、5头经饥饿处理24 h的黄带犀猎蝽成虫,24 h后统计捕食情况。每处理重复3次。
1.3 数据统计
1.3.3 Hassell干扰模型E=QP-m(E=Ne/NP)。式中,E为竞争下的捕食作用率;Q为搜寻常数;P为捕食者的密度;m为干扰常数;Ne为捕食的猎物总数;N为初始猎物密度(范悦莉等,2019; Hassell & Verley,1969)。
1.3.4 分摊竞争强度I=(E1-Ep)/E1。式中,E1为1头捕食者的捕食作用率;EP为捕食者密度为P头时的捕食作用率(罗春萍等,2018; 周军辉等,2020)。
以上数据均采用Excel 2007和SPSS 22.0进行统计分析。
2 结果与分析
2.1 黄带犀猎蝽对斜纹夜蛾幼虫的捕食功能反应

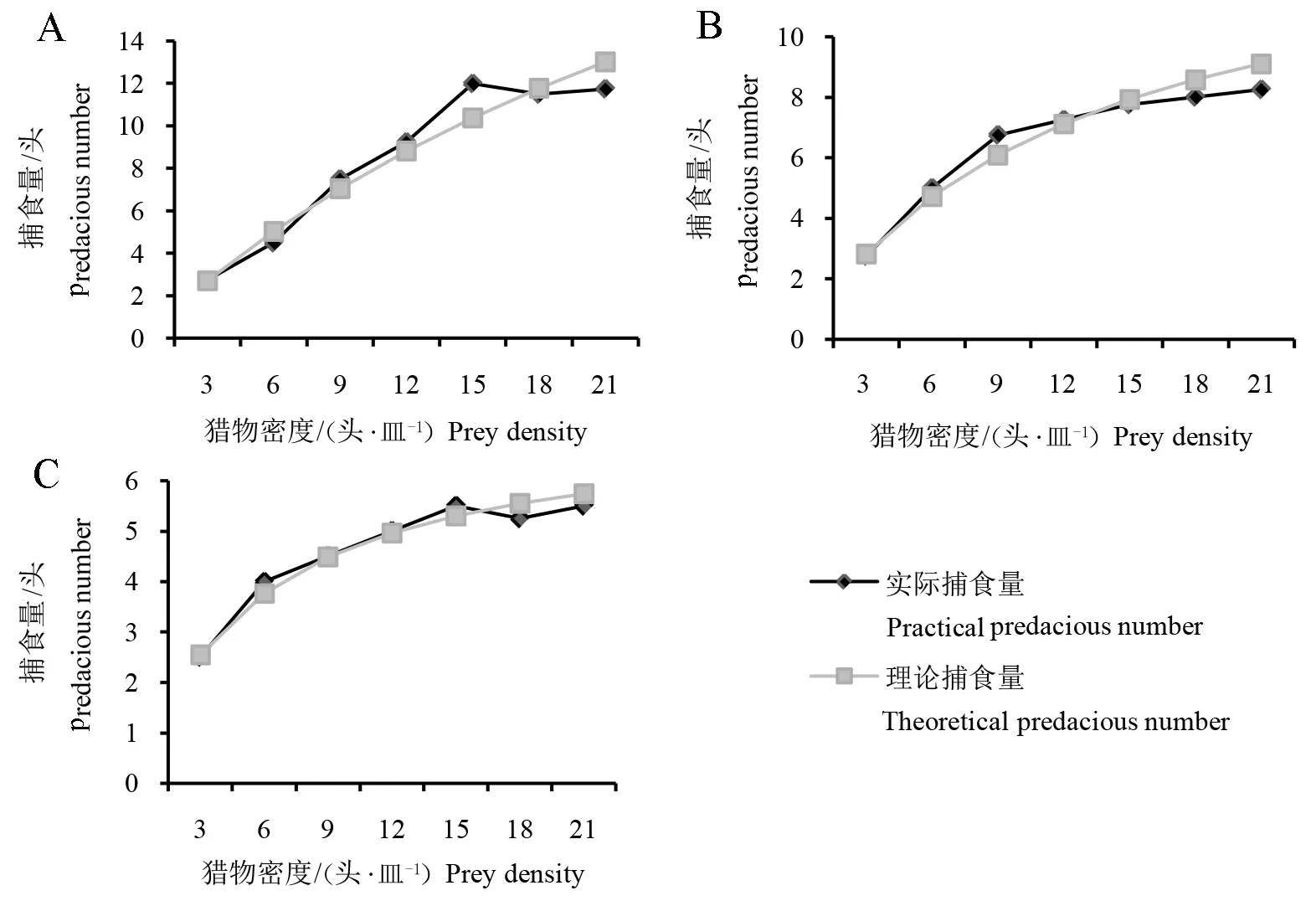
图1 黄带犀猎蝽对斜纹夜蛾2龄(A)、3龄(B)、4 龄(C)幼虫的捕食功能曲线
根据拟合得到的HollingⅡ公式计算可得,黄带犀猎蝽成虫对斜纹夜蛾2、3、4龄幼虫的瞬间攻击率(a)依次为0.9750、1.1654、1.3058,处置时间(Th)依次为0.0280、0.0690、0.1378 h,理论最大捕食量(1/Th)依次为35.7、14.5、7.3头。可见,随着斜纹夜蛾幼虫龄期的增加,黄带犀猎蝽成虫捕食斜纹夜蛾幼虫的瞬间攻击率逐渐增强,处置时间逐渐延长,理论最大捕食量逐渐减小。从评价天敌对猎物捕食效能的综合指标a/Th来看,黄带犀猎蝽成虫对斜纹夜蛾幼虫的捕食能力随着斜纹夜蛾幼虫龄期的增加而逐渐减弱(表1)。

表1 黄带犀猎蝽成虫对斜纹夜蛾幼虫的捕食功能反应参数
2.2 黄带犀猎蝽对斜纹夜蛾幼虫的搜寻效应
将瞬间攻击率(a)和处置时间(Th)带入搜寻效应方程,黄带犀猎蝽成虫对斜纹夜蛾2、3、4龄幼虫的搜寻效应方程分别为S=0.9750/(1+0.0273N)、S=1.1654/(1+0.0804N)、S=1.3058/(1+0.1799N)。通过搜寻效应方程计算可得不同斜纹夜蛾幼虫密度下的搜寻效应值,得到的搜寻效应曲线如图2所示。除猎物密度为3头·盒-1时,黄带犀猎蝽成虫对斜纹夜蛾幼虫的搜寻效应随斜纹夜蛾幼虫龄期的增加而逐渐减弱;当斜纹夜蛾幼虫处于同一龄期时,黄带犀猎蝽成虫对斜纹夜蛾幼虫的搜寻效应随着斜纹夜蛾幼虫密度的增加而逐渐减弱。当猎物密度为3头·盒-1时,黄带犀猎蝽成虫对斜纹夜蛾3龄幼虫的搜寻效应略大于其对2龄幼虫的搜寻效应,考虑是由于在猎物密度极低的情况下,其搜寻效应值均较大,趋近于1所致。

图2 黄带犀猎蝽成虫对斜纹夜蛾幼虫的搜寻效应
2.3 黄带犀猎蝽自身密度对捕食功能的干扰反应
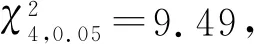

表2 黄带犀猎蝽自身密度对捕食效应的影响
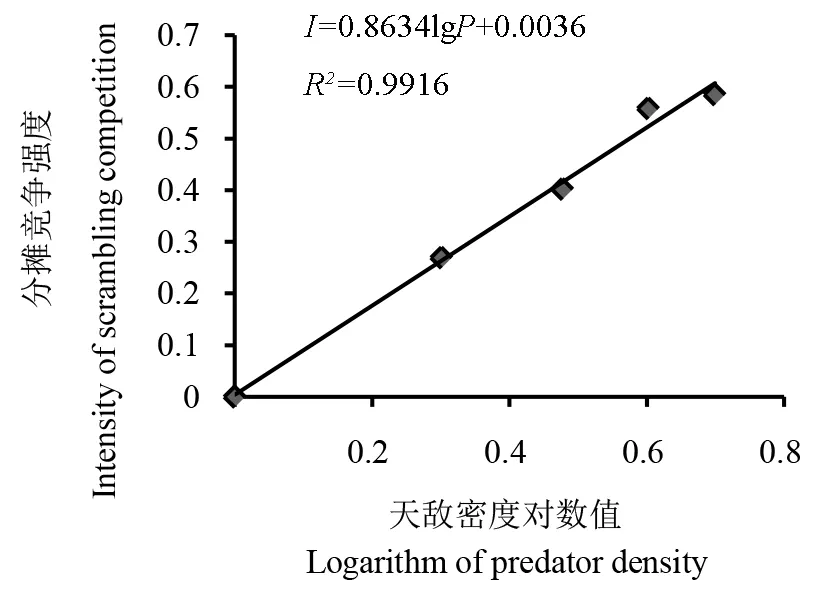
图3 黄带犀猎蝽种内分摊竞争强度(I)与自身密度(P)的关系
3 讨论与结论
本研究对黄带犀猎蝽成虫捕食斜纹夜蛾2、3、4龄幼虫的行为观察,发现黄带犀猎蝽成虫以口针插入斜纹夜蛾幼虫虫体吸食其体液,被取食后的斜纹夜蛾幼虫呈干瘪状死亡,有时并不取食完全,将口针收回后接着寻找下一头猎物,该捕食过程与王亚楠等(2020)报道的黄带犀猎蝽4、5龄若虫对草地贪夜蛾3龄幼虫的捕食行为相似,同时结合黄增和等(1991)关于黄带犀猎蝽的生物学特性研究,认为黄带犀猎蝽若虫和成虫的捕食行为特征无明显差异,而主要表现为捕食能力和捕食量上的差异。
本研究运用HollingⅡ捕食功能反应方程和HasseⅡ干扰模型方程评价了黄带犀猎蝽成虫对斜纹夜蛾2、3、4龄幼虫的捕食反应、搜寻效应和自身密度干扰反应。结果显示:黄带犀猎蝽成虫对斜纹夜蛾幼虫的捕食功能反应符合HollingⅡ型,即其捕食量随猎物密度的增加而增加,其搜寻效应随猎物密度的增加而减小;黄带犀猎蝽成虫自身密度干扰反应符合HasseⅡ型,即随着其自身密度的增加,平均捕食量逐渐减少,捕食作用率逐渐降低。本研究利用捕食功能反应参数对比了黄带犀猎蝽对斜纹夜蛾2、3、4龄幼虫的捕食能力差异,结果发现,随着斜纹夜蛾幼虫龄期的增加,黄带犀猎蝽成虫捕食斜纹夜蛾幼虫的瞬间攻击率逐渐增强,但搜寻效应逐渐减弱,处置时间逐渐延长,理论最大捕食量逐渐减小。从评价天敌对猎物捕食效能的综合指标a/Th来看,黄带犀猎蝽成虫对斜纹夜蛾幼虫的捕食能力随着斜纹夜蛾幼虫龄期的增长而逐渐减弱,结合斜纹夜蛾幼虫在空间分布格局上表现为随着幼虫龄期的增长聚集强度逐渐减弱,1~4龄幼虫的空间分布型为聚集分布,5~6龄幼虫空间分布型由聚集分布逐渐趋向于均匀分布(杜浩等,2020)。因此,在利用黄带犀猎蝽成虫控制斜纹夜蛾幼虫的实践中,应尽可能在斜纹夜蛾幼虫低龄期实施防控。
天敌对害虫的控制能力除了受到天敌自身的密度、害虫的密度影响外,还受到天敌和猎物的性别、虫态、龄期、丰富度,环境温度、湿度、光周期,寄主植物种类、生育期等诸多因素的影响(邱海燕等,2020; 唐敏等,2019)。本研究只开展了室内条件下一定空间内黄带犀猎蝽对斜纹夜蛾的捕食功能反应及自身密度干扰反应研究,未进行不同黄带犀猎蝽虫态、虫龄的捕食功能差异试验;未开展一定空间下,黄带犀猎蝽与斜纹夜蛾比例相同、密度成倍增加情况下的相互干扰反应试验;同时,本研究建立的Holling捕食功能反应方程和HasseⅡ自身密度干扰反应方程也未考虑田间环境因素,以及天敌和猎物种群的多样性等因素干扰。
