入侵植物三裂叶豚草在中国的潜在适生区预测
2021-12-20李佳慧叶兴状张明珠赖文峰刘益鹏张天宇张国防
李佳慧, 叶兴状, 张明珠, 赖文峰, 刘益鹏, 张天宇, 张国防, 刘 宝*
1福建农林大学林学院,福建 福州350002; 2福建省顺昌县林业科学技术中心, 福建 顺昌 353200
外来入侵物种是当前生物学研究的热点,具有天敌数量少、繁殖能力强的特点 (Hejdaetal.,2009)。中国地域辽阔,生物栖息地的类型多样化、适宜的环境条件使外来物种入侵后较易存活、建立种群(郭勤等,2002)。近年来,全球生物安全问题愈加突出,新颁发的《中华人民共和国生物安全法》将外来物种入侵防控提升到国家重大发展战略(胡孔新,2020)。而气候变化是当前研究的热点问题,全球变暖对生物多样性的影响越来越突出,人类活动是引起气候变化的重要原因(Karletal.,2003)。
物种分布模型(species distribution models,SDMs)是指利用当前存在的物种分布点信息与所需的环境变量相结合,通过特定算法建立模型,得出物种分布与环境变量之间的关系,利用物种分布模型对入侵物种潜在适生区进行预测(Guisan & Zimmermann,2000)。最大熵模型MaxEnt (maximum entropy model)是物种分布模型中的一种,预测精度高,软件易于使用(Phillipsetal.,2006),广泛应用于气候变化下外来入侵物种,如喜旱莲子草Alternantheraphiloxeroides(Mart.) Griseb.(Yanetal.,2020)、银胶菊PartheniumhysterophorusL.(Maharjanetal.,2019)、马缨丹LantanacamaraL.(Zhangetal.,2020)、豚草AmbrosiaartemisiifoliaL.(Chungetal.,2020)等潜在分布区的预测,均取得了较好效果。
三裂叶豚草AmbrosiatrifidaL.是菊科豚草属的一年生粗壮草本植物,被列入《国家重点管理外来入侵物种名录(第一批)》,常见于田野、路旁或河边的湿地,是一种恶性外来入侵植物,主要靠水、车辆和人为携带传播(郭琼霞等,2004)。三裂叶豚草原产于北美,1930年首次在辽宁省铁岭地区发现。近年来,随着人口流动,三裂叶豚草在国内迅速蔓延,对农业、畜牧业、旅游产业乃至人类健康造成影响。三裂叶豚草的花粉易使人产生过敏反应,如咳嗽、皮肤瘙痒红肿,严重时会导致胸闷(Aseroetal.,2005)。气候变暖的加剧,导致该植物花粉引发的过敏现象越来越严重(Rasmussenetal.,2017)。研究三裂叶豚草潜在适生区分布格局变化可为制定遏制其扩散的措施提供理论依据,从而减轻因该物种蔓延导致的健康及经济风险。
国内外已有不少学者对三裂叶豚草的化感作用(Wangetal.,2005)、分子标记(Sharmaetal.,2020)、防治措施(Liebman & Nichols,2020)、生物学特性(Abul-Fatihetal.,1979)等进行相关研究,但对其潜在分布区域的研究仅局限于单个省份(马倩倩等,2020),或仅使用气候、降水、地形因子进行全国范围的预测(王福刚,2017; Qinetal.,2015),尚未发现同时利用人类活动因子和环境因子对其空间分布的报道。本研究利用3种典型温室气体排放浓度(representation concentration pathways,RCP),即RCP2.6(处于最低状态的温室气体排放情景,该气候情景下全球平均温度上升幅度限制在2 ℃以内)(Vuurenetal.,2011)、RCP4.5(温室气体排放浓度中等时的情景)(Thomsonetal.,2011)和RCP8.5(处于最高温室气体排放的情景)(Riahietal.,2011)情景,考虑了应对气候变化不同策略对未来温室气体排放的影响,预测当前及未来气候条件下三裂叶豚草的潜在分布区。本研究所采用的人类活动数据来源于国际地球科学信息网络中心(CIESIN)的人类足迹(human footprint)数据层,能全面反映人类活动的程度,该数据的生成由建筑环境的范围、耕地、牧场地、人口密度、夜间灯光数据、铁路、道路、通航水道共8个数据层归一化得到(塞依丁·海米提等,2019; Venteretal.,2016)。本研究以中国为研究区,将ENMeval数据包建立的MaxEnt优化模型与ArcGIS软件相结合,揭示人类活动因子和环境因子对三裂叶豚草空间分布范围的影响,预测当前以及未来气候条件下三裂叶豚草在中国的潜在地理分布区,以期为预防和控制三裂叶豚草的入侵提供科学依据。
1 三裂叶豚草分布点数据收集与处理
1.1 三裂叶豚草分布点数据收集与处理
通过检索教学标本平台(http:∥mnh.scu.edu.cn/)、中国数字植物标本馆(https:∥www.cvh.ac.cn/)、中国国家标本资源平台(http:∥www.nsii.org.cn/2017/home.php,NSII)、中国植物图像库(http:∥ppbc.iplant.cn/,PPBC)、Global Biodiversity Information Facility(https:∥www.gbif.org/,GBIF)、中国自然标本馆(http:∥www.cfh.ac.cn/)和相关文献(达良俊等,2008; 韩平,2018; 黄岩,2016; 李明等,2014; 林夕,1989; 栾晓青,2020; 马倩倩等,2020; 王娟等,2013; 魏守辉等,2006; 杨波等,2012; 杨国禄等,2011; 杨再华等,2017; 赵浩宇等,2020; 张德满等,2009),获取三裂叶豚草的地理分布数据,舍弃分布点信息不全、重复记录的地点,再将分布记录进行地理校正和地理坐标转化,最终得到225条分布点的信息,涵盖21个省(地区)。为降低采样偏差,采用Arc GIS软件进行缓冲区分析,分析过程中遵循1 km×1 km格网中保留1个分布点的原则,最终获得筛选后的有效分布点120个。
1.2 环境因子数据获取与筛选
本研究下载WorldClim数据库GCM-CCSM4模式(http:∥worldclim.org)中的当代(current)以及未来2050s(2041—2060年)、2070s(2061—2080年)中3种典型温室气体排放浓度(RCP2.6、RCP4.5和RCP8.5)情景下的19个生物气候因子,分别代表温室气体排量浓度低、中和高3种情景,不同情景考虑了应对气候变化不同策略对未来温室气体排放的影响(Mossetal.,2010)。海拔因子也来源于WorldClim数据库。土壤因子来源于世界土壤数据库(http:∥www.fao.org/ faostat/en/#data.),选取其表层土壤中的17个因子。人类活动强度(human activity factor, hfp)数据来源于国际地球科学信息网络中心(http:∥www.ciesin.org/)的人类足迹数据层,为避免建模过程中多重共线性导致模型过度拟合(Graham,2003),本研究将19个生物气候因子和17个土壤因子,共计36个环境因子的空间分辨率统一为30″(30 arc-second,约1 km2),按照1∶1400万中国地图矢量图剪裁为一致大小,后经掩膜、重采样等步骤,得到像元大小完全一致的环境因子,并以asc格式输出。最终确定10个生物气候因子、4个土壤因子、1个海拔因子、1个人类活动因子共计16个环境变量参与建模。分析底图为标准地图服务系统(http:∥bzdt.ch.mnr.gov.cn/)下载的1∶1400万中国地图。
1.3 模型优化与精度评定
使用R语言中的ENMeval数据包,对调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)2个参数进行优化,将调控倍频设置为0.5~8,依次递增0.5,共16个调控倍频,采用11个特征组合(L、QT、H、HP、PT、QH、LQH、LPT、QHP、LQPT、LQHPT),其中L为线性(linear),Q为二次型(quadratic),H为片段化(hinge),P为乘积型(product),T为阈值型(threshold)(Velasco & Gonzalez-salazar,2019)。ENMeval数据包将上述176种参数组合进行测试,最终用训练AUC (area under the curve)和测试AUC值之差(the difference between training and testing AUC, AUCDIFF)和10%测试遗漏率(10% test omission rate, or 10pct)来测试模型对物种分布点的拟合程度,Akaike信息量准则(akaike information criterion correction, AICc)模型中的delta.AICc评估不同参数组合的复杂度和拟合度,最终选取具有最低AICc值(delta.AICc=0)时的最佳模型参数组合用于建模。
运用MaxEnt进行建模时,将包含120个三裂叶豚草分布点经纬度数据的csv格式文件与筛选出的16个环境因子数据同时导入MaxEnt中,随机选取25%的分布点作为测试数据集来验证模型,将剩余的75%的分布点作为训练数据集来建立模型,重复运行类型(replicated run type)设为Bootstrap,软件其余设置保持默认设置,重复10次。本研究采用受试者操作特征曲线(receiver operating characteristic curve, ROC)的面积AUC来评估模型的精确性。AUC值不依赖于正确预测的存在比例和正确预测的不存在比例,因此被认为是模型评价最高效的方法(Mcphersonetal.,2004)。AUC值越大则模型预估效果越好。AUC的取值范围为[0,1],一般认为,当AUC值<0.8,表示模拟结果较差,0.8≤AUC<0.9,模拟结果一般,0.9≤AUC<1.0。模拟结果较好。
1.4 适生度等级分区及面积统计
将当代和未来不同气候情境下的数据在MaxEnt软件中使用最优化模型进行模拟,利用ArcGIS软件的重分类工具,结合三裂叶豚草当前主要分布区范围以及物种分布概率值,采用人工(manual)分级方法(刘艳梅等,2018),将三裂叶豚草适宜生境的适宜度指数(habitat suitability index, HIS)(冉巧等,2019)划分为4个等级:HIS<0.2为非适生区;0.2≤HIS<0.4为低适生区;0.4≤HIS<0.7为中适生区;HIS≥0.7为高适生区。最后,利用Arc GIS栅格计算对重分类的图层进行面积制表,从而得到每个分区的权重,根据裁剪出的国土实际面积换算为每个等级的面积。
1.5 适生区空间格局变化
通过ArcGIS软件,对未来不同情景数据模拟出来的平均值结果文件进行重分类。将三裂叶豚草分布概率值<0.2的空间单元定为不适生区,赋值为0;分布概率值≥0.2为适生区,赋值为1,以此建立未来气候变化情景下三裂叶豚草潜在地理分布的存在/不存在(0,1)矩阵。矩阵值0→1为迁入新增区,1→0为迁出丧失区,1→1为恒有稳定区。其中:迁入新增区是指当前气候情景下不存在但在未来气候情景下存在的气候区域;迁出丧失区是指在当前气候情景下存在但在未来会消失的区域;恒有稳定区是指在当前和未来气候情景下均保留的区域。最后,在ArcGIS软件中导入矩阵变化值,使用软件绘制出三裂叶豚草空间迁移格局的变化图。
1.6 适生区质心(centroid)迁移
通过计算某些属性的质心迁移可以反映它们在时空尺度上的发展特征。在做质心迁移前,假设三裂叶豚草具备完全迁移能力,忽略物种间的相互作用以及地理阻隔作用等因素,设定一个用于分析空间格局变化的极端无限制迁移假设条件。利用ArcGIS中的相关技术工具,统计当前以及未来6个气候变化情景下适宜区域的质心,每个质心在三裂叶豚草分布概率值≥0.2的空间单元内生成,并分别比较当前与2050s和2070s不同RCP的气候变化情景下物种质心位置的变化,计算质心位置迁移的距离。
1.7 多元环境相似度面和最不相似变量
利用多元环境相似度面(multivariate environmental similarity surface,MESS)和最不相似变量(the most dissimilar variable,MoD)对未来不同气候情景和引起三裂叶豚草适生区发生变化的主要环境因子进行探讨。MESS用来衡量特定时期某点的气候与参考图层气候的相似程度,若多元相似度为正值(最大值为100),代表该点气候无异常,若出现负值,表明该点为气候异常点,即至少一个环境变量的值超出了参考图层对应值的范围。MoD为某点相似度最低或异常程度最高的变量,异常程度最高的环境变量即有可能成为引起物种地理分布变迁的关键因素(Elithetal.,2010)。
2 结果与分析
2.1 模型优化结果与准确性评价
基于120条三裂叶豚草的当代分布记录和筛选出的16个环境因子图层,利用MaxEnt模型对其当代潜在分布区进行模拟,MaxEnt的默认调控倍率RM=1,特征组合FC=LQHPT,delta.AICc=NA。当RM=3.5、FC=LQH时,delta.AICc=0,根据AICc信息量准则,此模型在本次模拟中为最优模型。同默认参数下的模型相比,最优模型的训练集AUC与测试集AUC平均差值(0.0240)和10%测试遗漏率平均值(0.1255)均低于默认参数下的模型(分别为0.0340、0.2135),并且下降了29.3%和41.2%,说明与默认参数的模型相比,该参数设置下模型的过拟合程度有所下降。因此,此次研究选择RM=3.5、FC=LQH为最终的设置参数。在最优参数设置条件下使用MaxEnt模拟预测当代地理分布中可以看出,10次重复训练的AUC值的平均值为0.9287,测试AUC值的平均值为0.9127,训练AUC值与测试AUC值均在0.9~1.0,表明此次模型预测较好。
2.2 影响三裂叶豚草空间分布的环境变量
确立影响三裂叶豚草当前地理分布区的主导环境因子主要通过参与建模的16个环境因子的贡献率、置换重要值和刀切图进行综合考虑(图1、表1)。由贡献率(percent contribution)可看出,人类活动因子(hfp,69%)、等温性(bio3,8.9%)和年降水量(bio12,7.3%)数值排名靠前,贡献率累计达85.2%,表明这3个因子是影响分布的主要环境因子;置换重要值(permutation importance)排名靠前的是人类活动因子(hfp,51.7%)、等温性(bio3,26.6%)和温度季节变动系数(bio4,9.7%),累计置换重要性为88.0%;在刀切法检验中,当用仅此变量模拟时,模型的正则化训练增益、测试增益和AUC值最高的3个因子均为人类活动因子(hfp)、年降水量(bio12)和海拔(ELEV),说明人类活动因子、年降水量和海拔对物种分布贡献最大;使用除此变量以外的其他变量时,正规化训练增益、测试增益降低最多的3个变量依次为人类活动因子(hfp)、等温性(bio3)和温度季节变化系数(bio4),AUC值下降幅度最大的3个变量人类活动因子(hfp)、温度季节变化系数(bio4)和等温性(bio3),表明这些变量包含更多其他变量所不存在的信息。综合以上,影响三裂叶豚草潜在适生区分布的环境因子主要有人类活动因子(hfp)、等温性(bio3)、温度季节变动系数(bio4)、年降水量(bio12)和海拔(ELEV)。
2.3 不同气候条件下三裂叶豚草适宜生境分布和空间变化格局
由图2可知,当前气候条件下三裂叶豚草在我国的潜在地理分布十分广泛,除西藏外,其他地区均有其潜在适生区,其中青海、甘肃西北部、云南省和新疆南部为零星分布。高适生区主要集中在东北平原、江淮地区、京津冀、山东半岛、四川盆地、关中平原、长江三角洲、珠江三角洲及福建沿海等经济发达地区。三裂叶豚草总适生区面积(176.93×104km2)约占国土面积的18.43%,其中高、中和低适生区分别占1.05%、5.01%、12.37%。

图2 MaxEnt模型预测的当前气候条件下三裂叶豚草潜在适生区[审图号:GS(2021)6506号]
由图3可知,与当前气候条件下的分布预测结果相比,在未来RCP2.6、RCP4.5和RCP8.5 3种气候情景下,三裂叶豚草在我国的潜在适生区范围均呈现逐渐减少趋势,总适生区面积也逐渐减少,与Qinetal.(2015)的研究结果相一致。在RCP2.6气候情景下,2050s和2070s的低适生区面积较当前气候条件下分别减少18.27%、20.62%,中适生区面积分别减少21.30%、29.62%,高适生区面积分别减少21.72%、32.47%;RCP4.5气候情景下,2050s和2070s的低适生区较当前气候条件下分别减少23.21%、2.28%,中适生区面积分别减少26.93%、9.10%,高适生区面积分别减少29.53%、15.21%;RCP8.5气候情景下,2050s和2070s的低适生区面积分别减少22.42%、8.81%,中适生区面积分别减少19.29%、5.21%、高适生区面积分别减少27.42%、15.29%。

图3 MaxEnt模型预测的未来气候条件下三裂叶豚草潜在适生区[审图号:GS(2021)6506号]
由图4可知,未来3种不同浓度排放情景中,三裂叶豚草潜在适生区范围均呈减少的趋势,即丧失面积大于增加面积。单从增加率来看,2070sRCP4.5的气候情景下面积增加率最大,为4.63%,增加区域主要位于江西、湖南、福建和浙江。RCP2.6的气候情景下,2070s面积的变化率大于2050年,主要表现为保留率减少、丧失率增加,说明在2050sRCP2.6的气候情景下三裂叶豚草更能稳定存活。RCP4.5气候情景下,2050s面积的变化率大于2070年,丧失面积比较大,丧失区域主要分布于长江以南的省(地区)。RCP8.5气候情景下,2050s面积的变化率大于2070年,丧失面积较未来气候条件下的任何时期都大,为45.55×104km2,丧失区域主要分布于新疆、河南、河北南部和山东西部。
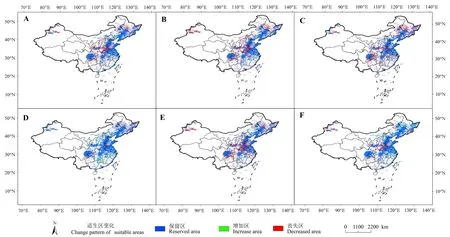
图4 不同时期三裂叶豚草空间变换格局[审图号:GS(2021)6506号]
2.4 三裂叶豚草质心转移分析
从图5可知,当前气候条件下,三裂叶豚草的质心点在山东省临清市(36°47′31.64″N,115°41′19.08″E)。RCP2.6情景下,当前—2050年,三裂叶豚草质心点位置向东南方向迁移了68.77 km,位于山东省齐河县(36°36′43.07″N,116°25′35.91″E);2050—2070年,质心向东南方迁移46.72 km,位于山东省长清区(36°25′19.22″N,116°53′34.66″E)。RCP4.5情景下,当前—2050年,质心点位置向东南方向迁移26.70 km,位于山东省茌平区贾寨镇(36°38′43.17″N,115°55′32.79″E);2050—2070年,质心向西偏南方迁移56.77 km,位于山东省莘县罗庄村(36°20′25.65″N,115°24′58.97″E)。RCP8.5情景下,当前—2050年,质心点位置向东北方向迁移了84.80 km,位于山东省禹城市(36°51′18.98″N,116°38′17.34″E);2050—2070年,质心向东南方迁移16.22 km,位于山东省齐河县(36°47′34.68″N,116°48′10.53″E)。综上,在不同排放情景下,三裂叶豚草的质心均有高纬度向低纬度迁移的趋势,这说明温度因子对三裂叶豚草的分布格局有重要影响。

图5 三裂叶豚草当代适生区分布及质心转移[审图号:GS(2021)6506号]
2.5 多元环境相似度面和最不相似变量分析
MaxEnt模型预测未来不同气候条件时期三裂叶豚草多元环境相似度面与最不相似变量结果(图6、7),在2050sRCP2.6、2070sRCP2.6、2050sRCP4.5、2070sRCP4.5、2050sRCP8.5、和2070sRCP8.5 6个气候环境条件下,三裂叶豚草120个当代分布点的多元相似度为负值的点所占比例分别为30.00%、25.83%、25.00%、25.00%、23.33%和15.83%。2050sRCP2.6多元相似度值最低,负值点比例最高,表明其气候异常程度最高,主要气候异常区出现在黑龙江、吉林、内蒙古、辽宁大部分区域、河北、山西、四川北部、新疆除中部以外的地区和甘肃西北部,最不相似变量为最冷月最低温度(bio6);2070sRCP2.6气候情景下,气候异常区与2050sRCP2.6基本相同,但在内蒙、甘肃和新疆区域呈略微缩减趋势,最不相似变量为最冷月最低温度(bio6);2050sRCP4.5和2070sRCP4.5气候情景下,气候异常区域主要集中区域同2070sRCP2.6相同,但在内蒙、辽宁和甘肃有略微收缩,最不相似变量为最冷月最低温度(bio6); 2050sRCP8.5气候情景下,主要气候异常区集中在黑龙江、吉林东部、内蒙东部、四川西北部、新疆除中部以外地区、甘肃、山西、河北和海南小部分区域,最不相似变量为最冷月最低温度(bio6)、年均温(bio1); 同2050sRCP8.5相比,2070sRCP8.5气候情景下气候异常区的区域在新疆、甘肃、吉林、山西和四川呈减少趋势,湖南、广西、广东和海南有增加趋势,最不相似变量主要为最冷月最低温度(bio6)、最湿季度平均温度(bio8)和年均温(bio1)。
综合图6、7结果,最冷月最低温度(bio6)在未来不同时期三裂叶豚草潜在适生区内作为主要最不相似变量,出现频次最多且占据适生区域绝大部分,其次为最湿季度平均温度(bio8)、年均温(bio1)。综上,最不相似变量出现最多的均为温度因子类型,说明温度因素对引起三裂叶豚草潜在地理分布变迁起着重要作用。

图6 MaxEnt模型预测的未来6个时期多元环境相似度面[审图号:GS(2021)6506号]
3 讨论
3.1 预测结果评估
研究发现,MaxEnt模型在默认设置下易过度拟合,不利于模型转移,因此,Muscarellaetal.(2015)基于R语言开发了ENMeval数据包,辅助选择模型最优参数设置,并得到了一定应用(Salvà-Catarineuetal.,2021)。在本研究中,通过调用ENMeval数据包,选出最佳优化参数,利用优化模型来预测当前和未来气候条件下三裂叶豚草在中国的潜在分布区域。三裂叶豚草在各个时期的潜在分布域的AUC值>0.9,说明模型预测结果较准确。
3.2 限制三裂叶豚草分布的主导环境变量
由图1、表2综合比对分析可得,影响三裂叶豚草潜在分布区的环境因子主要有人类活动因子、温度因子、降水因子、海拔因子。其中,人类活动影响对三裂叶豚草分布的贡献最大,约占69%,可见人类活动强度对其入侵分布影响很大,这与三裂叶豚草在耕地、路边、公路沿线、河岸渠道两侧、住宅区等地出现较为频繁的结论一致(黄可辉等,2006; 马倩倩等,2020)。迈迪娜·吐尔逊(2017)在新疆地区的意大利苍耳XanthiumitalicumMoretti适生区研究中指出,入侵物种的传播和扩散易受人类活动的影响。本研究发现,除人类活动因子对该物种的分布起到重要影响外,降水和温度因子也起到了重要作用,这与王福刚(2017)和Qinetal.(2015)的结论一致。年降水量(bio12)在贡献率中排名第四。研究表明,三裂叶豚草喜欢生长在肥沃、湿润的土壤(黄可辉等,2006),对水分条件较为敏感,其最佳生长区域位于雨量充足的湿润地区(王国骄等,2014)。因此,降水因子对其分布起到重要的作用。温度因子中排名较前的有等温性(bio3)、温度季节变动系数(bio4),马倩倩等(2020)的研究也指出,温度季节性变化对三裂叶豚草有重要作用。三裂叶豚草的种子具有二次休眠的特性,需经过长时间的低温冷冻才能打破休眠开始萌发,长时间的恒温条件不利于其种子的发芽,适当的变温会增加种子的发芽率(Bassett & Crompton,1975)。因此,温度对三裂叶豚草分布格局起着重要制约作用。
由多元环境相似度面和最不相似变量结果可得,2050sRCP2.6气候异常程度最高,最冷月最低温度(bio6)为该时期气候异常区的唯一存在的最不相似变量,说明该因子对三裂叶豚草的分布具有巨大的影响。2070sRCP2.6、2050sRCP4.5、2070sRCP4.5这3个时期中,最冷月最低温度(bio6)对物种分布格局影响较大。研究表明,低温层积处理对解除其种子的休眠具有重要作用,不同休眠深度的种子对于解除休眠所需时间不同(魏守辉等,2006)。因此,三裂叶豚草潜在地理分布格局主要制约环境因子为最冷月最低温度(bio6)。
气候因子并非相互独立影响三裂叶豚草的适生区分布范围,多元环境相似度面和最不相似变量表明,未来6种气候变化情景下(RCP2.6、RCP4.5、RCP8.5)适生区内气候异常的关键性因子有最冷月最低温度(bio6)、最湿季度平均温度(bio8)、年均温(bio1)。说明三裂叶豚草地理分布迁移是多种因子综合作用的结果,其主导因子的作用效果也受到其他因子的影响。
3.3 不同气候背景下三裂叶豚草的空间格局变化
三裂叶豚草在未来6个气候情景下分布范围总体来说有不同程度的缩减,即丧失率大于增加率。从结果中可以看出,时期相同,温室气体排放浓度不同时,三裂叶豚草的缩减程度也不一样,其丧失区面积随温室气体排放浓度升高而增加。研究表明,未来气候情景条件下,部分生长在高纬度地区的外来入侵物种数量将呈减少趋势(Bellardetal.,2013)。由于高温条件下,三裂叶豚草种子胚胎的呼吸受到细胞膜的抑制,因此,其在高温条件下易休眠、不易发芽(Davisetal.,1930)。这也验证了随着全球气温的上升,该入侵物种的适生区分布范围逐渐减小。三裂叶豚草的种子成熟后即进入休眠状态,需冬季低温条件才能解除其休眠现象,开始萌发。并且,三裂叶豚草耐湿润而不耐干旱,其所生存环境的土壤含水量降低会对叶片的生物膜造成严重损害(Davisetal.,1930)。因此,三裂叶豚草在未来高温的气候情景下呈现逐渐南移或靠沿海移动的趋势。
3.4 三裂叶豚草的防治策略
由MaxEnt模型预测结果可得,三裂叶豚草当前潜在总适生区面积约占国土面积的18.43%,主要分布在中国东部地区,并有逐渐向内陆地区扩散的趋势。虽然当前气候条件下潜在分布区域较小,但其繁殖能力强,抗逆性强(郭琼霞等,2005),一些种子在埋藏9年后仍能存活(Harrisonetal.,2007),甚至有些种子能存活15年以上(Hartnettetal.,1987),这种休眠特性是三裂叶豚草暴发的基础,有利于其扩散分布,还为该入侵植物的防治增添许多难度。因此,对于当前三裂叶豚草的潜在分布地区,如东北平原、江淮地区、京津冀、新疆地区、山东半岛、四川盆地、关中平原、长江三角洲、珠江三角洲及福建沿海等地区,当地检疫部门应做好本职工作,在入侵地域开展防除工作,在开花期前对现有的种群实行全面清除,应积极贯彻“预防为主,综合防治”的植物保护方针,注重化学防治、生物防治和物理防治措施相结合,注重防治后的生态恢复问题;对中低潜在适生区的地域,应积极开展三裂叶豚草的科普工作,树立社会大众对于入侵物种的防治观念。完善海关检疫制度,做好监测防控工作。加强对进口货物、车辆流通以及人口流动的检疫力度,注重河流、道路等人畜扰动大、水分充足地域的监测防控工作。
