异黄酮类植物雌激素防治乳腺癌的研究进展*
2021-12-20黄照权
李 玉,黄照权
(桂林医学院 1.药学院;2.基础医学院,桂林 541199)
流行病学调查显示,乳腺癌是女性中最常见的癌症[1],占女性所有新诊断癌症的30%,是20~59岁癌症死亡的主要原因[2],严重危害女性的生命和健康。传统化疗和内分泌治疗是除外科手术外最常用的治疗方法,然而大多数治疗药物都有很强的副作用或耐药性[3]。因此,人们在积极寻找一种毒性更小、更有效的替代物。
异黄酮类植物雌激素化学结构与内源性雌激素十分相似,因此在最佳浓度时可以通过与雌激素受体(ER)结合,发挥抗雌激素的作用[4]。已有研究表明,异黄酮类植物雌激素通过多种靶点和机制发挥抗乳腺癌作用,且未出现明显副作用和耐药性,因此有望成为有价值的抗癌药物。
1 植物雌激素的结构与分类
植物雌激素是植物来源的膳食化合物,主要存在于大豆中。其结构类似于内源性雌激素17-β-雌二醇(E2),共同特征是有两个苯环,一个相对稳定和平坦的分子,以及2个或3个羟基,通常以共轭形式存在(图1),分子量也与E2相似。植物雌激素按结构可以分为异黄酮类、黄酮类、木脂素、香豆素和二苯乙烯类[5],而在豆类、葛根等中大量存在的异黄酮类植物雌激素是最重要的,主要包括刺芒柄花素(formononetin)、毛蕊异黄酮(calycosin)、染料木黄酮(genistein)、大豆苷元(daidzein)、生物禅宁A(biochanin A)等[6]。
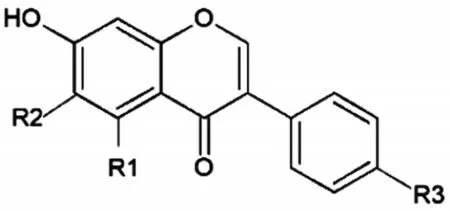
图1 异黄酮类植物雌激素的结构[7]
2 异黄酮类植物雌激素的流行病学研究
近年来流行病学研究表明,亚洲的乳腺癌发病率明显低于西方国家,推测与大豆中富含的异黄酮类植物雌激素的保护作用有关。据统计,亚洲居民平均每天摄入25~50 mg 异黄酮,而西方居民平均每天摄入低于2 mg 的异黄酮[7]。Zhao 等[8]通过对11 169 例乳腺癌病例和648 913 名参与者进行荟萃分析后发现,当异黄酮摄入量较高时,患病风险显著降低。Micek等[9]对15项队列研究进行的荟萃分析发现,异黄酮摄入量(<62.7 mg/d)与乳腺癌总死亡率和复发之间存在显著的负相关关系。Rienks等[10]对10项观察性研究进行的荟萃分析发现,高浓度的大豆苷元和染料木黄酮分别降低34%和28%乳腺癌风险。Veheus 等[11]的病例对照研究也表明,在荷兰女性中,血液样本中异黄酮水平的增加与患乳腺癌风险降低有关。此外每天10 mg大豆异黄酮摄入量能降低9%的乳腺癌死亡率[12]。因此,较高的异黄酮摄入量对乳腺癌有显著的防治作用。
3 异黄酮类植物雌激素抗乳腺癌的药理作用及作用机制
3.1 抑制乳腺癌细胞增殖及移植瘤生长 Choi等[13]研究表明,将大豆苷元(1~100 μmol/L)作用于MCF-7 和MDA-MB-453 细胞24 h、48 h 或72 h 后,大豆苷元以剂量和时间依赖的方式显著抑制细胞增殖(P<0.05)。Tian 等[14]研究发现,毛蕊异黄酮通过WDR7-7-GPR30 信号抑制乳腺癌细胞的增殖。染料木黄酮抑制MCF-7细胞增殖,而对非致癌细胞无明显影响[15]。生物禅柠A能降低MCF-7和T-47D癌细胞的增殖,也能抑制MCF-7乳腺癌异种移植动物模型中的肿瘤生长[16-17]。此外,生物禅柠A 与人参皂苷Rh2 联合应用可协同抑制MDA-MB-231 和MCF-7细胞的增殖[18]。在体内实验中,Moon等[17]研究表明,生物禅柠A每天腹腔注射一次能有效降低MCF-7小鼠移植瘤的发病率和生长,表明生物禅柠A可作为为乳腺癌的预防药物。刺芒柄花素在小鼠癌症模型中也表现出抑制肿瘤增殖的能力,与对照组相比,刺芒柄花素治疗组的肿瘤重量减少了39.6%[19]。此外,毛蕊异黄酮能显著降低T47D 小鼠移植瘤生长[20]。
3.2 抑制乳腺癌细胞迁移和侵袭 肿瘤坏死因子-α(TNF-α)通过增强Gli1核转位和转录活性进而激活Hedgehog(Hh)信号,诱导乳腺癌细胞侵袭。研究表明,大豆苷元通过抑制TNF-α 诱导的Gli1/Hh 信号的激活,从而抑制癌细胞迁移和侵袭[21]。Wu等[22]研究表明,毛蕊异黄酮能够使致癌基因Rab27B 依赖性信号失活进而抑制MDA-MB-231 细胞的迁移和侵袭。Li等[20]研究表明,通过靶向Foxp3(在乳腺癌中高度表达的转录因子)介导的血管内皮生长因子(VEGF)和基质金属蛋白酶(MMP)-9 的表达,高剂量的毛蕊异黄酮治疗能够降低MCF-7和T47D的迁移和侵袭能力。Zhang 等[20]研究证明,毛蕊异黄酮通过BATF/转化生长因子β1(TGFβ1)抑制上皮—间质转化(EMT),进而呈时间和剂量依赖性地抑制T47D 和MCF-7 细胞迁移和侵袭。此外,染料木黄酮和大豆苷元通过抑制MMPs 活性在高侵袭性MDA-MB-231细胞中发挥抗侵袭活性[21]。
3.3 阻滞乳腺癌细胞周期进程 细胞周期受不同的细胞周期蛋白(Cyclins)、细胞周期蛋白依赖性激酶(CDKs)和细胞周期蛋白依赖性激酶抑制剂(CDKI)的表达调控,各个时期会因生长因子或致癌刺激而转变。Cyclin D1是研究最广泛的癌基因,其在超过50%的乳腺中高表达[23],控制着细胞周期从G1到S 的相变。研究表明,刺芒柄花素可降低MCF-7和MDA-MB-453细胞CyclinD1mRNA和蛋白表达,使细胞周期阻滞在G0/G1 期[24]。高浓度大豆苷元(100μmol/L)能降低MCF-7 和MDA-MB-453 细胞中CyclinD1、CyclinA、CyclinB 和CDK1、CDK2、CDK4 的表达,使细胞周期阻滞在G1 和G2/M 期从而发挥其在人乳腺癌细胞中的抗癌作用[13]。
研究最多的CDKI 是p21、p27 和p57,它们属于Cip/Kip 家族。大豆苷元可上调CDKIp21、p57 在MCF-7细胞的表达,也能增加MDA-MB-453细胞中p27 的表达。但大豆苷元不能增加MCF-7 细胞中p27 的表达[25]。这也表明异黄酮类植物雌激素在不同的乳腺癌细胞系中可能会产生不同的作用。
总之,异黄酮类植物雌激素通过降低Cyclins和CDKs 的表达,以及增加不同的CDKI 的表达,从而阻滞多种乳腺癌的细胞周期进程。
3.4 诱导乳腺癌细胞凋亡 细胞凋亡主要通过外源性死亡受体途径和内源性线粒体途径。外源性途径包括配体如TNF-α、Fas 配体(Fas-L)等与死亡受体结合,导致受体分子寡聚化。而线粒体介导的凋亡完全受Bcl-2 家族蛋白的调控,Bcl-2 家族分为Bcl-2、Bax和BH3超家族。Bcl-2是细胞凋亡最重要的抑制因子之一,Bax 则是决定细胞凋亡的关键[26]。有研究报道,染料木黄酮通过显著增加Fas-L、FADD、细胞色素C(cytc)、截短型BID、caspase3和caspase9的表达来诱导细胞凋亡[27]。大豆苷元通过激活caspase9和caspase7,上调Bax、降低Bcl-2蛋白的表达,导致cytc的释放,最终使MCF-7和MDAMB-453细胞凋亡[13]。Tian等[26]研究证明,毛蕊异黄酮能增强细胞凋亡调节蛋白RASD1的表达,进而上调Bax和下调Bcl-2来诱导人乳腺癌细胞MCF-7凋亡。以上研究表明,异黄酮类植物雌激素通过内源性或外源性途径诱导乳腺癌细胞凋亡,增加促凋亡蛋白的表达,减少抗凋亡蛋白的表达。
3.5 对ERα 和β 表达的调节 ERα 的激活能刺激乳腺组织中的细胞增殖,而ERβ抑制细胞增殖和促进凋亡,与健康组织相比,在乳腺癌中ERα/ERβ 比值明显上升[28]。Marik等[29]研究表明,染料木黄酮显著降低3 种不同ER 阳性乳腺癌细胞MCF-7、T47D和21PT中ERα 的表达,并增加ERβ 的表达,从而抑制癌细胞增殖。在ER 阳性乳腺癌细胞中,毛蕊异黄酮和刺芒柄花素可通过上调ERβ、下调胰岛素样生长因子1受体(IGF-1R)、激活凋亡标志物PARP-1以及下调miR-375发挥抗肿瘤活性[30]。提示异黄酮类植物雌激素也可以通过提高乳腺癌细胞中ER-β/ER-α比值,从而抑制乳腺癌细胞增殖。
3.6 调控miRNA表达 多项研究表明,miRNAs在癌症中异常表达或突变,其可作为一类新的癌基因或肿瘤抑制基因发挥抗肿瘤作用[31]。Avci 等[32]报道,染料木黄酮上调MCF-7 乳腺癌细胞中miR-23b的表达进而抑制乳腺癌细胞的生长。在另一项研究中,染料木黄酮(1~5 μmol/L)下调肿瘤细胞miR-155 的表达,并上调miR-155 靶点(如促凋亡叉头家族转录因子FOXO3及其下游靶点p27、酪蛋白激酶和PTEN)在MDA-MB-435 细胞中的表达来降低乳腺癌细胞的活性[33]。
3.7 对血管生成的影响 血管生成是指从现有的血管系统中形成新的血管,向肿瘤细胞输送营养物质和氧气[34]。VEGF 是血管生成的中枢介质,刺激肿瘤血管的生长,血管生成失控是肿瘤生长、侵袭和转移的关键[34]。余小平等[35]研究发现,经染料木黄酮处理后,乳腺癌细胞中VEGF 表达显著降低。Su 等[36]也发现,在常氧和低氧条件下,染料木黄酮剂量依赖性地降低了VEGF的表达,在低氧条件下,染料木黄酮抑制细胞缺氧诱导因子-1(HIF-1)的活化,表明染料木黄酮的抗血管生成作用是通过抑制HIF-1激活、下调VEGF表达来实现的[34]。
3.8 抑制NF-κB信号通路和PI3K/Akt信号通路
NF-κB是一种转录因子,参与调控细胞增殖、炎症、肿瘤形成和肿瘤进展等关键生物学过程[37]。研究表明,大豆异黄酮通过剂量依赖性地降低NF-κB/p65 蛋白水平和NF-κB 的DNA 结合活性,下调Bcl-2,上调Bax 抑制MDA-MB-231 细胞增殖,诱导凋亡[37]。染料木黄酮通过Notch-1 途径以剂量依赖的方式抑制NF-κB 的活性,并影响抗凋亡基因(Bcl-2和Bcl-XL)的表达,从而抑制MDA-MB-231 细胞活性[38]。
PI3K/Akt 信号转导途径的异常激活已在包括乳腺癌在内的多种癌症中得到证实,并在细胞增殖、生长、迁移、代谢和凋亡中起关键作用。毛蕊异黄酮和刺芒柄花素通过显著降低MCF-7和T47D细胞中IGF-1R蛋白表达,抑制MCF-7细胞中Akt的磷酸化[39-41]。刺芒柄花素亦能显著抑制MDA-MB-231细胞中PI3K和Akt的磷酸化表达[42]。毛蕊异黄酮和染料木黄酮通过抑制PI3K/Akt信号通路,降低下游靶基因HOTAIR 的表达进而抑制乳腺癌细胞的生长[43]。此外,PI3K/Akt通路可导致mTOR过度激活,从而导致肿瘤细胞的无限增殖[44]。刺芒柄花素通过抑制PI3K/Akt/mTOR 通路增强依维莫司对乳腺癌MDA-MB-468细胞的抑制作用[44]。NF-κB信号通路和PI3K/Akt 信号通路可作为抗癌药物的新的治疗靶点。
4 结语和展望
异黄酮类植物雌激素通过多种靶点和机制抑制乳腺癌细胞的活性,已然成为抗乳腺癌的新候选药物。然而,目前大量的研究集中在植物雌激素对乳腺癌的体外作用,未来仍需要更多体内和临床研究进一步评估异黄酮类植物雌激素防治乳腺癌的有效性和安全性,并确定这些化合物安全有效的最佳浓度,为临床乳腺癌的治疗提供更多方案。
猜你喜欢
杂志排行

广西医科大学学报的其它文章
- miR-508-3p靶向AFF4/TGFβ1信号通路调控人牙髓细胞分化的机制研究*
- 腹壁剖宫产瘢痕子宫内膜异位症恶变为透明细胞癌1例*
- 急性心肌梗死与2型糖尿病相关差异基因的GEO芯片分析*
- Effect of curcumin on palmitic acid-induced apoptosis and steroid production of ovarian granulosa cells by inhibiting HMGB1-mediated autophagy
- 白藜芦醇对人甲状腺乳头状癌细胞IHH4细胞恶性生物学特征及PI3K/Akt信号通路影响*
- 金丝桃苷对糖尿病肾病大鼠TGF-β1/smad通路及肾上皮间质转化的影响*