牛支原体iTRAQ 定量蛋白质组学分析
2021-12-03李海利徐引弟朱文豪张青娴焦文强王治方王克领郎利敏张立宪
李海利,徐引弟,游 一,朱文豪,张青娴,焦文强,王治方,许 峰,王克领,郎利敏,张立宪,邹 靖
(1. 河南省农业科学院畜牧兽医研究所,河南郑州 450002;2. 河南省信阳市畜牧工作站,河南信阳 464000)
牛支原体肺炎是由牛支原体(Mycoplasma bovis,M.bovis)引起牛的一种高度接触性传染性呼吸道疾病[1],其感染可导致牛生长发育迟缓、饲料利用率降低、咳嗽、呼吸困难、高烧、喘气等,从而造成严重的经济损失。此外,牛支原体可引起免疫抑制,并因此导致其他呼吸道病原(如巴氏杆菌、链球菌)的混合或继发感染,使损失更加严重。牛支原体除可引起呼吸道疾病外,还可导致关节炎、乳房炎、子宫内膜炎、眼角膜结膜炎、耳炎、生殖道炎及流产与不孕等多种疾病[2‑5]。牛支原体肺炎首次于1961 年在美国发生[6‑7],当时从患乳腺炎的牛乳中分离到牛支原体病原[8‑9],其他国家也相继发生并暴发了牛支原体肺炎。湖北是我国首先发生牛支原体肺炎的省份,随后在全国流行。目前,牛支原体肺炎在世界范围内流行[10‑12],发病率和死亡率均较高。经过长途运输后的奶牛或者肉牛,更易诱发牛支原体肺炎,给全球养牛业造成了很大的经济损失[13‑16]。
蛋白质是细胞功能和生命活动的重要载体,也是生物体内遗传信息表达的最后形式。利用微生物主要免疫保护蛋白或者相应抗原制备亚单位苗或者基因工程疫苗,是动物疫病防控的研究方向之一。iTRAQ 差异蛋白质组学是目前常用的蛋白质组学研究工具,可以鉴定出生物体内几乎所有的蛋白质。目前应用较多的是iTRAQ 定量差异蛋白质组学,可以鉴定生物体或者微生物不同时间内的差异蛋白质,为揭示生命现象规律提供新的方法和思路。
随着规模化养牛业的不断发展,当前牛支原体肺炎已成为牛的严重呼吸系统疾病,并常常发生混合感染。牛支原体隐性持续感染,是牛场呼吸系统疾病发生的根本原因,是当前呼吸系统疾病的主要病因。牛肺炎支原体可在牛群中水平传播,也可垂直传播,必须采取综合防控措施。目前,牛支原体肺炎主要采取药物治疗,但是药物防治只能缓解症状和降低经济损失,难以彻底净化。因此,人们尝试通过疫苗接种来防治牛支原体的感染,研发用于牛支原体肺炎的预防、诊断和控制的新方法、新策略迫在眉睫,为牛支原体肺炎防控提供保障。采用iTRAQ 差异蛋白质组学研究策略,以临床分离的菌株HN12 和标准菌株PG45 的全蛋白质为研究对象,对牛支原体全谱蛋白质及差异蛋白质进行比较,旨在为研发中支原体蛋白质疫苗或研究其致病机制奠定基础。
1 材料和方法
1.1 材料
1.1.1 菌株 牛支原体标准菌株(PG45);临床分离牛支原体菌株(HN12)[17],来源于河南地区某养牛场肺脏。
1.1.2 主要试验仪器 质谱仪AB SCIEX Nano LC/MS/MS(Triple TOF 5600 Plus)。
1.2 方法
1.2.1 菌株复苏培养及保存 把标准菌株(PG45)和临床分离菌株(HN12)接种于PPLO 琼脂平板上,置于体积分数为5%的CO2培养箱中培养24~72 h,挑取单菌落进行纯化培养。
挑取单菌落置于PPLO 液体培养基,37 ℃、180 r/min 振荡培养24 h,4 000 r/min离心30 min,弃上清液,收集菌体,-80 ℃冷冻保存备用。
1.2.2 蛋白质提取及定量 取-80 ℃冷冻保存菌体,在菌体中加入适量裂解液(8 mol/L 尿素、1%SDS);漩涡振荡混匀,置于冰上超声4 min,冰上裂解30 min;4 ℃离心30 min,取上清。上清采用Bradford法定量,并绘制标准曲线。
1.2.3 酶解和标记及质谱分析 取经过定量的蛋白质上清,使样品中的蛋白质含量为100 μg,进行酶解标记及质谱分析,显著性差异蛋白质数目按照P<0.05、差异倍数(FC)<0.5 或>2.0 条件筛选,质谱分析在武汉金开瑞公司进行。对鉴定出的所有蛋白质进行生物信息学分析。
2 结果与分析
2.1 牛支原体蛋白质的提取及含量测定
采用TCA-丙酮法对样品进行蛋白质的提取。选择常规的Bradford 定量方法测定总蛋白质,并绘制标准曲线,所得曲线方程为y=0.006+0.025x,R2=0.997。根据标准曲线计算样品的含量(表1)。然后进行SDS-PAGE电泳(图1)。从图1可以看出,提取的蛋白质电泳条带清晰,符合下一步质谱分析需求,临床分离菌株和标准菌株在个别条带处存在明显差异。
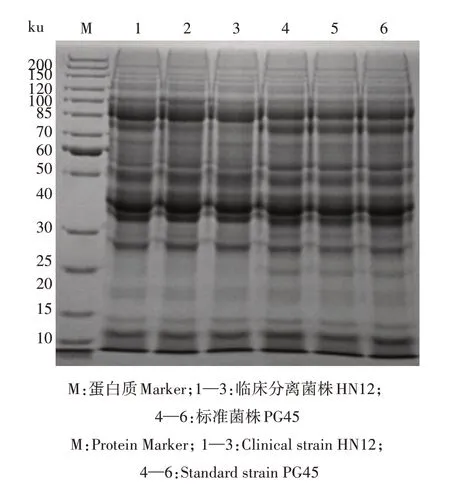
图1 牛支原体临床分离菌株和标准菌株蛋白质SDSPAGE电泳结果Fig.1 SDS-PAGE results of protein from the clinical isolate and standard isolate of Mycoplasma bovis
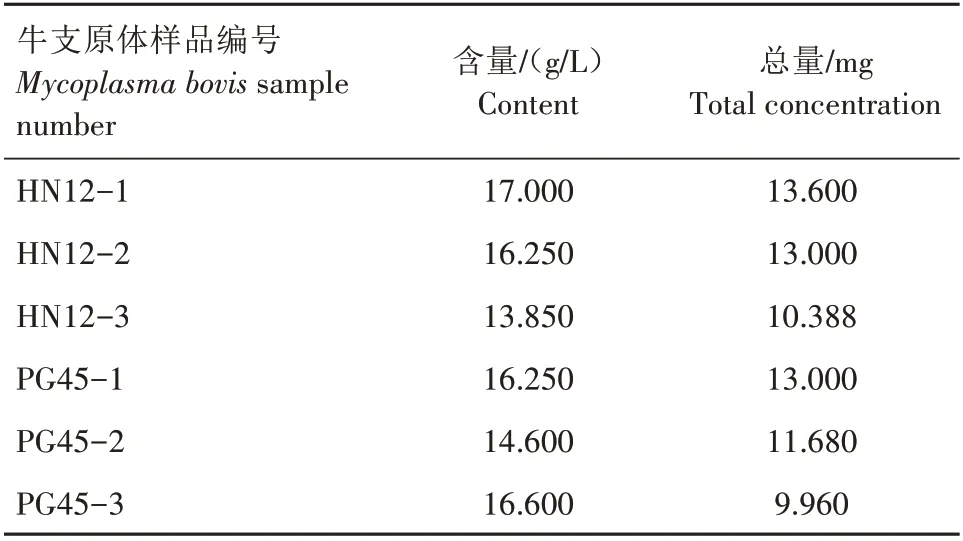
表1 样品蛋白质含量Tab.1 Protein concentration of sample
2.2 牛支原体蛋白质鉴定信息
经iTRAQ 标记及质谱分析,从HN12 和PG45 菌株中共鉴定出2 402 个蛋白质,存在显著差异的有66 个。其中,相对于标准菌株,临床分离菌株上调蛋白质数量为25个(表2),下调蛋白质数量为41个(表3)。上调蛋白质主要为无特征的蛋白质、超家族蛋白的水解酶、假定的膜蛋白、缬氨酸连接酶、转座酶家族蛋白、假定的脂蛋白、依赖DNA 的RNA 聚合酶β 亚基、无机焦磷酸酶、50 S 核糖体蛋白L10、跨膜蛋白、果糖三磷酸醛缩酶、50 S 核糖体蛋白L14、三甲胺脱氢酶、转座酶、磷酸甘油酸酯激酶、延长因子、30S 核糖体蛋白S10、酰胺酶、尿苷三磷酸-葡萄糖-1-磷酸尿苷酰基转移酶、50 核糖体蛋白L13、磷酸酯酶家族蛋白。下调蛋白质主要为亮氨酰氨基肽酶、乙醇脱氢酶、丙酮酸脱氢酶复合物E1-β 亚基、甘油醛-3-磷酸脱氢酶、延长因子Tu、膜脂蛋白P81、延长因子Ts、丙酮酸脱氢酶复合物E1-β亚基、50 S 核糖体蛋白L11、50 S 核糖体蛋白L7/L12、依赖ATP 的锌金属蛋白酶FtsH、伴护蛋白质DnaK、氧化还原酶锌结合脱氢酶、L-乳酸脱氢酶、LemA 家族蛋白、变量表面脂蛋白、假定的脂蛋白、ABC 转运蛋白ATP 结合蛋白、甘油醛-3-磷酸脱氢酶无特征的蛋白质、假定底物结合蛋白、ATP 合成酶亚基β 蛋白、磷酸戊糖变位酶、30S 核糖体蛋白S7、二氢利坡酰胺琥珀酰(酸)转移酶、烯醇酶、乙醇脱氢酶、磷酸戊糖变位酶、可变表面脂蛋白VspF、膜脂蛋白P81、二氢硫辛酸脱氢酶、NADH 氧化酶、延长因子G、假定的脂蛋白、NADH脱氢酶等。
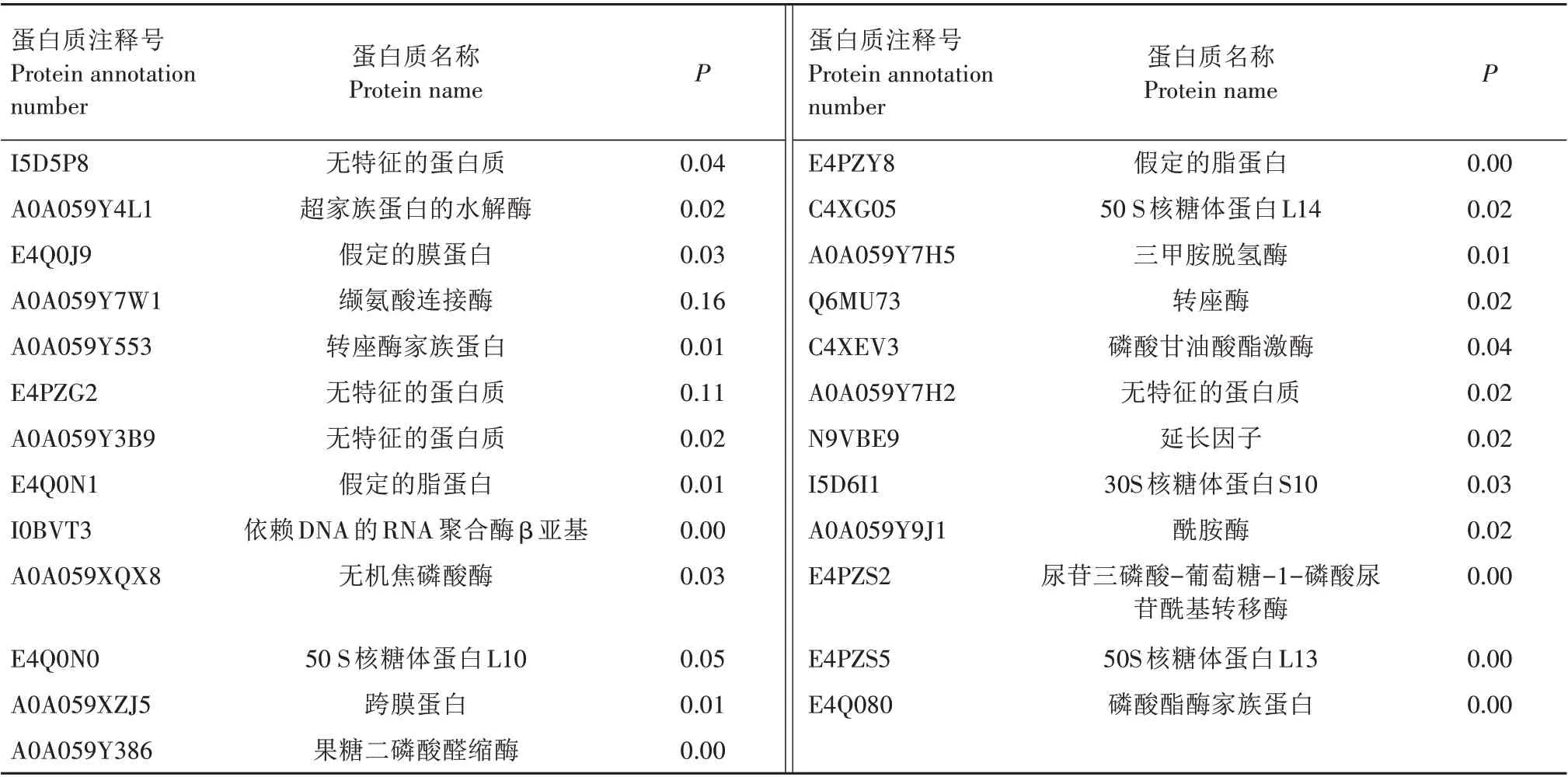
表2 牛支原体中鉴定出的上调蛋白质Tab.2 The identified upregulated proteins of Mycoplasma bovis

表3 牛支原体中鉴定出的下调蛋白质Tab.3 The identified down-regulated proteins of Mycoplasma bovis
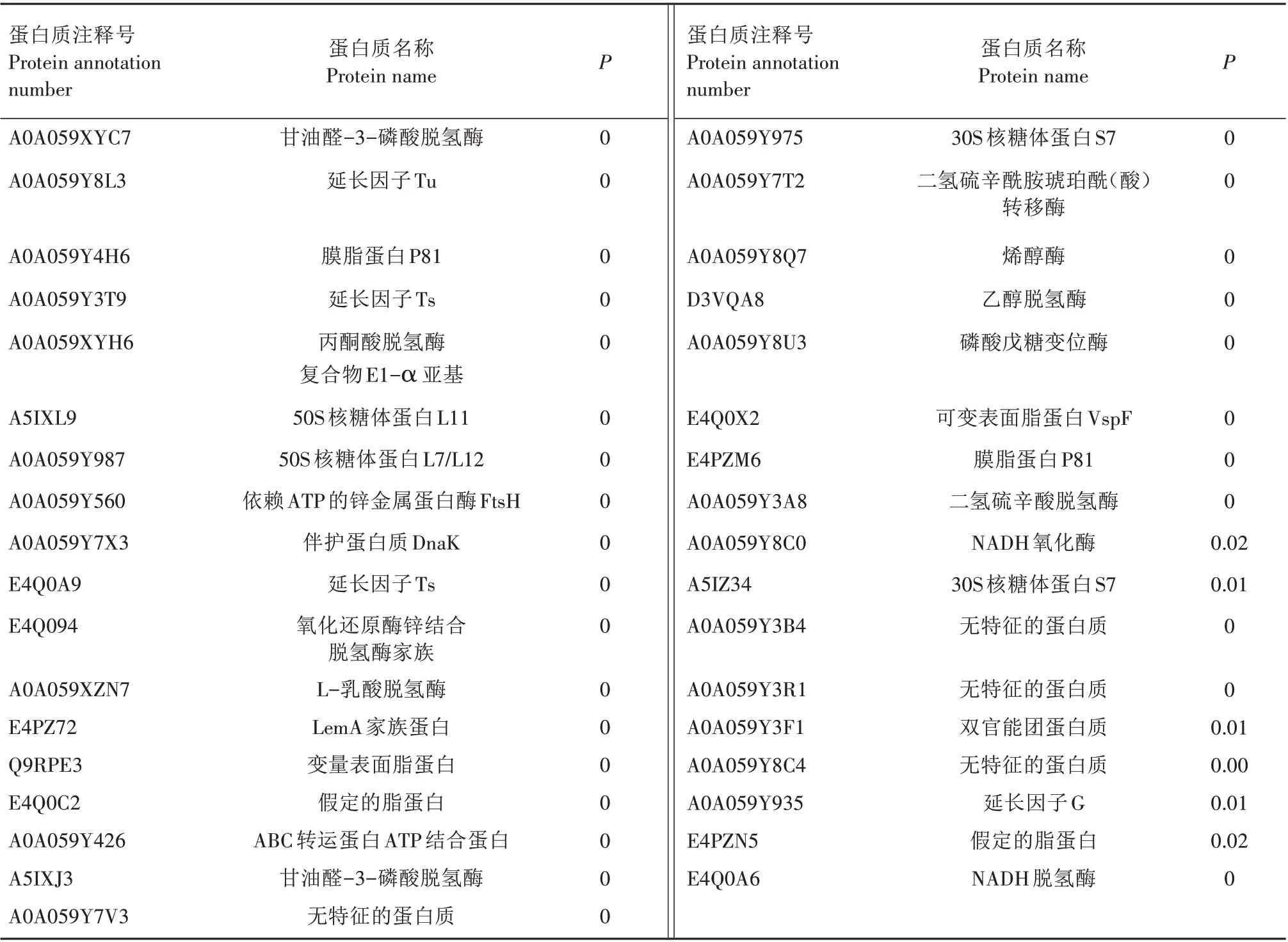
续表3 牛支原体中鉴定出的下调蛋白质Tab.3(Continued) The identified down-regulated proteins of Mycoplasma bovis
2.3 牛支原体蛋白质生物信息学分析
2.3.1 全谱分析 依据分子功能、细胞组分和参与的生物学过程,将定量得到的2 402 个牛支原体蛋白质进行功能注释(图2)。从图2 可以看出,分子功能的蛋白质主要参与核酸结合转录因子活性、结合、运输活动、催化活性等,其中结合功能的蛋白质最多。细胞组分包括的蛋白质主要参与细胞器、细胞组分、细胞膜组分、大分子复合物等,其中参与细胞组分的蛋白质最多。生物学过程蛋白质主要参与生物过程调节、细胞组成、组织或生物形成、代谢过程和生物代谢过程等,其中参与代谢过程的蛋白质最多。
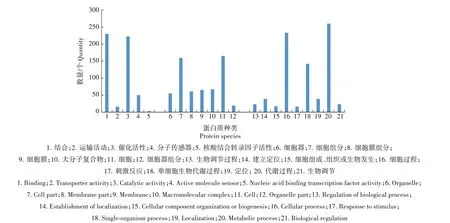
图2 牛支原体蛋白质的GO二级分类统计Fig.2 GO secondary classification statistics of Mycoplasma bovis protein
2.3.2 KEGG 注释通路分析 对蛋白质数目最多的前20个通路进行分析,了解蛋白质和基因的生物学功能(图3)。从图3 可以看出,包含蛋白质最多的前20 个通路为核糖体、转氨酰基生物合成、糖酵解和糖异生、嘌呤代谢、嘧啶代谢、ABC 转运蛋白质、丙酮酸代谢、氧化磷酸化、磷酸戊糖途径、同源重组、光合生物碳固定、RNA 降解、甲烷代谢、光合作用、DNA 复制、RNA 聚合酶、视黄醇代谢、错配修复蛋白质输出和丙酸乙酯代谢。其中包含蛋白质最多的通路为核糖体通路。
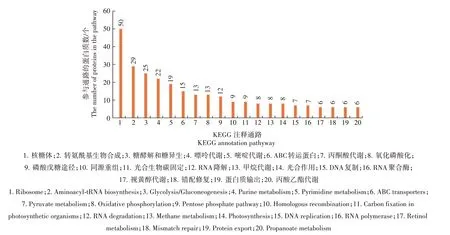
图3 牛支原体蛋白质KEGG注释通路Fig.3 KEGG annotation pathway of Mycoplasma bovis proteins
2.3.3 差异蛋白质表达模式聚类分析和差异蛋白质模块表达趋势分析 用R 语言中gplot 包将牛支原体标准菌株与临床分离菌株差异显著的蛋白质进行表达模式聚类分析(图4)和模块(Cluster)表达趋势折线分析(图5)。从图4 可以看出,41 个下调蛋白质在标准菌株中的表达量高于临床分离菌株中的表达量,其中,甘油醛-3-磷酸脱氢酶、L-乳酸脱氢酶、乙醇脱氢酶、NADH氧化酶和延长因子G的差异倍数分别为0.48、0.48、0.48、0.47 和0.46;25 个上调蛋白质在临床分离菌株中的表达量高于标准菌株,其中,30S 核糖体蛋白S10、延长因子、假定的脂蛋白和假定的膜蛋白的差异倍数分别为10.76、9.80、6.25和4.04。
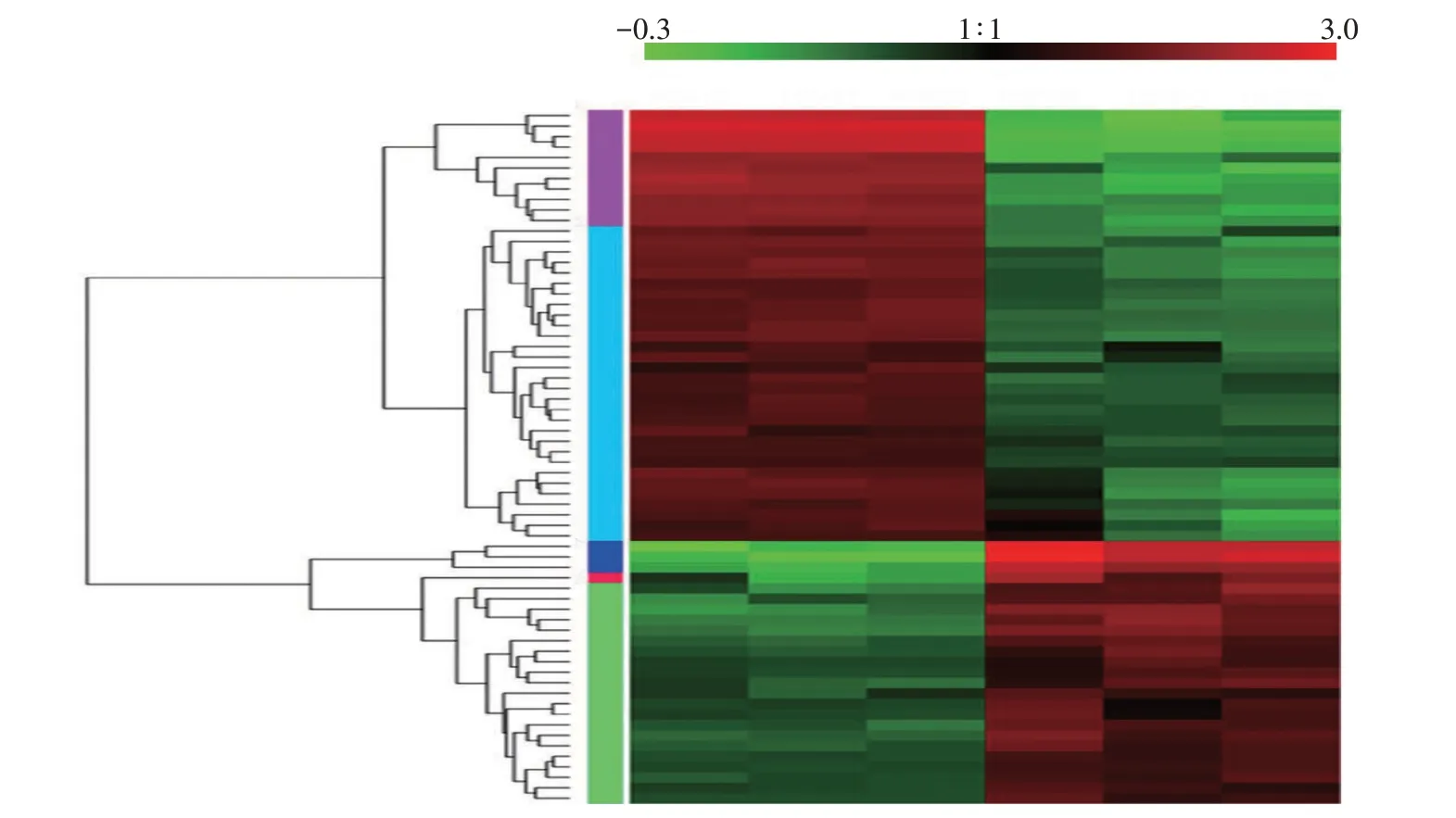
图4 牛支原体差异蛋白质表达模式聚类Fig.4 Clustering diagram of differential protein expression of Mycoplasma bovis
图5 中,模块1 为21 个上调蛋白质(2.0<差异倍数<4.0)表达趋势,其在临床分离菌株中的表达量高于标准菌株;模块2为30个下调蛋白质(0.2<差异倍数<0.5)的表达趋势,其在临床分离菌株的蛋白质表达量低于标准菌株中的表达量;模块3 为3 个上调蛋白质(6.0<差异倍数<10.0)的表达趋势,其在临床分离菌株中的表达量高于标准菌株;模块4 为11 个下调蛋白质(0.1<差异倍数<0.2)的表达趋势,这11个下调蛋白质在临床分离菌株中的表达量低于标准菌株。

图5 牛支原体标准菌株与临床分离菌株差异蛋白质模块表达趋势Fig.5 Expression trend of differential protein modules between the standard strains and clinical isolate of Mycoplasma bovis
3 结论与讨论
蛋白质组学研究是生命科学领域常用的工具,具有强大的数据挖掘功能,几乎能把样本中的所有蛋白质都鉴定出来[18‑19],是目前研究免疫原性蛋白质功能性蛋白、诊断试剂、疫苗候选抗原常用的技术手段[20‑23]。本研究应用差异蛋白质组学技术,对牛支原体标准菌株和临床分离菌株进行了差异蛋白质组学研究,共鉴定差异蛋白质66个。鉴定出的上调蛋白质主要有超家族蛋白的水解酶、延长因子等,下调蛋白质主要有亮氨酰氨基肽酶、乙醇脱氢酶、丙酮酸脱氢酶复合物E1-β亚基、膜脂蛋白P81、延长因子Ts、丙酮酸脱氢酶复合物E1-β 亚基等。牛支原体细胞膜表面含有大量的变量表面脂蛋白[24‑25]。表面脂蛋白C 端具有抗原决定簇和黏附结构。本研究鉴定出的表面脂蛋白在临床分离株中明显下调。
前人研究表明,牛支原体标准菌株PG45 共编码765个蛋白质[26‑30]。本研究结果显示,丙酮酸脱氢酶复合物E1-β亚基、延长因子、膜脂蛋白P81、延长因子Ts等,在临床分离菌株中明显下调。延长因子是原核细胞进行蛋白质翻译过程中所需的因子[31],本研究共鉴定2 种延长因子,即延长因子G 和延长因子Ts[32‑33]。前人研究表明,丙酮酸脱氢酶复合物E1-β 亚基是牛支原体重要的免疫原性蛋白质,可用于牛支原体的诊断和疫苗研究[34‑36]。
本研究通过差异蛋白质组学研究标准菌株PG45 与临床分离菌株HN12 之间的蛋白质差异。共鉴定2 402个蛋白质,存在显著差异的有66个,标准菌株相对于临床分离菌株上调蛋白质数量为25个,下调蛋白质数量为41个。其中鉴定出多个免疫原性蛋白质,为牛支原体抗原的筛选和诊断提供参考,为牛支原体病防控和基因工程亚单位疫苗的筛选提供了重要的蛋白质信息,也为诊断试剂、新药物及疫苗研发提供依据。
