姜曲海猪FoxO3 基因组织表达和多态性与肉质关联分析
2021-12-03朱淑斌倪黎纲陈章言张君胜周春宝赵秀玲
朱淑斌,倪黎纲,陈章言,张君胜,周春宝,赵秀玲
(1. 江苏农牧科技职业学院,江苏泰州 225300;2. 姜曲海猪保种场,江苏泰州 225300)
叉头框(Forkhead box,Fox)蛋白家族有19 个亚族,FoxOs 是Fox 家族的重要成员,在哺乳动物中主要有FoxO1、FoxO3、FoxO4 和FoxO6 四种转录因子,均高度保守,分别表达于不同的组织器官[1]。FoxO转录因子作用复杂多样,包括氧化应激、细胞代谢、DNA 修复、细胞周期阻滞、参与细胞凋亡和自噬等,与动物机体的很多组织细胞发育及代谢密切相关,如对肌肉组织、脂肪组织和肝脏等都有非常重要调控机能[2‑3]。前人研究发现,FoxO1和FoxO3都是维持肌肉能量稳态的重要因素[4]。目前,对FoxO3基因研究多集中在人和小鼠方面,检测到韩国人FoxO3基因存在7 个多态位点[5]。WANG 等[6]和COSKUN 等[7]报道了FoxO3基因多态性与长寿相关,且与性别有一定的相关性。刘源等[8]报道了小鼠FoxO3基因对骨形态发生蛋白15 启动子区的转录活性有影响。在哺乳动物卵巢中,FoxO3基因可以通过促进卵巢颗粒细胞凋亡来调节卵泡的闭锁有关[9]。有关FoxO3基因在家畜方面的研究很少。猪的FoxO3基因定位于1 号染色体,含有2 个外显子和1个内含子。于晶[10]研究发现,FoxO3基因在第2 个外显子存在遗传多态性,并与胴体性状指标中的胴体体斜长、6~7 肋间背膘厚和大理石纹进行相关分析,结果显示,该位点的多态性与这几个胴体性状间均存在显著相关关系。王玲[11]研究发现,FoxO3基因在肉牛群体中分别存在8 个多态性位点,FoxO3基因单倍型组合与肌内脂肪(Intramuscular fat,IMF)含量、肌肉嫩度、肌纤维直径和肌纤维密度性状之间存在不同程度的相关性。陈亚楠[12]对FoxO3基因进行克隆及序列比对,共检测到3个多态位点与猪生长性状有关联。
姜曲海猪是江苏省的一个历史悠久的优良地方猪种,具有产仔较多、性温驯、早熟易肥、脂肪沉积能力强、肉质鲜美等特点,是“北腿”(如皋火腿)的主要原料猪之一,是我国地方猪种的宝贵资源。但目前对姜曲海猪肉质特性及其相关分子遗传基础了解甚少,不利于该珍贵遗传资源的保护和开发利用。为此,利用序列特异引物引导的聚合酶链式反应(Polymerase chain reaction‑sequence specific primer,PCR-SSP)方法检测地方猪种姜曲海猪的FoxO3基因多态性,分析其对猪肉质性状的遗传效应,并探索该基因的表达规律,旨在寻找与姜曲海猪肉质性状相关的遗传标记,为提高肉质的研究奠定基础,并为姜曲海猪种质资源特性及标记辅助选择(Marker assisted selection,MAS)的研究提供参考。
1 材料和方法
1.1 试验材料
112 头姜曲海猪、40 头苏姜猪和30 头杜长大猪由姜曲海种猪场提供,分别取约0.5 g 耳组织,于-20 ℃保存。试验猪群饲养至8 月龄,选择含有不同基因型的试验猪屠宰,取左侧胴体最后肋与第一、二腰椎间中心部位,带回江苏农牧科技职业学院动物科技学院猪肉质检测实验室进行肉质性状测定和肌肉组织学特性测定;选择姜曲海猪30 只,其中0(初生)、1、2、4、6、8 月龄各5 只,屠宰后取心脏、肝脏、脾脏、肾脏、腹脂、皮下脂肪、背最长肌、比目鱼肌、腓肠肌和半腱肌样品,放入1.5 mL 离心管中,迅速置于液氮中,带回实验室后,置于-70 ℃冰箱中保存,以备总RNA的提取。
1.2 主要试剂
Trizol 为美国Invitrogen 公司产品;DEPC 处理水、琼脂糖为大连宝生物技术有限公司产品;Tris碱、EDTA 为美国Sigma 公司产品;HiFi-MMLV cDNA 第一链合成试剂盒、SYBR PrimeScriptTMRTPCR Kit 和5×GoldstarTaqPCR Buffer 均为北京康为生物有限公司产品;氯仿、异丙醇和无水乙醇等分析纯均为国药集团化学试剂公司产品。
1.3 试验方法
1.3.1 DNA、RNA 的提取和检测 采用苯酚-氯仿法[10]提取猪耳组织基因组DNA,将其溶解在RNasefree ddH2O 中,-20 ℃保存备用。采用RNA 提取试剂盒对心脏、肝脏、脾脏、肾脏、腹脂、皮下脂肪、背最长肌、比目鱼肌、腓肠肌和半腱肌进行总RNA 提取。检测样本浓度和OD 值后,用1.5%的琼脂糖凝胶电泳对DNA、RNA质量进行检测。
1.3.2 PCR 扩增及其产物分析 参照GenBank 中FoxO3基因第二外显子序列,利用Primer Premier 5.0软件设计引物(FC:5′-GTCATTGAGATTCATGG‑TCC-3′;FT:5′-GTCATTGAGATTCATGGTCT-3′;R:5′-CGCCTGTCCCCCATCCTG-3′),引物由生工生物工程(上海)股份有限公司合成。PCR-SSP 扩增体系(20 μL):5×GoldstarTaqPCR Buffer 4 μL,GoldstarTaqPCR Polymerase 1 μL,dNTPs 2 μL,H2O 9 μL,上下游引物各1 μL,模板DNA 2 μL;PCR扩增程序:95 ℃预变性10 min;95 ℃变性30 s,59 ℃退火30 s,72 ℃延伸1 min,35 个循环;72 ℃延伸7 min,冷却至4 ℃。制备2%琼脂糖凝胶(加入溴化乙锭至终质量浓度0.5 mg/L),上样量5 μL,以TAE为电泳缓冲液,100 V 电泳10 min,于紫外灯下观察。
1.3.3 实时荧光定量PCR 根据NCBI 公布的猪FoxO3基因的cDNA 序列(NM_001135959.1),利用Primer Premier 5.0 软件设计目的基因引物(上游引物:5′-CGCACTACGGCAACCAGACA-3′;下游引物:5′-TAGGCTGGGCGGCAAAGGAC-3′),β-actin基因(上游引物:5′-ACACGGTGCCCATCTACGAG-3′;下游引物:5′-GCTTCTCCTTGATGTCCCGC-3′)作为内参,引物由生工生物工程(上海)股份有限公司合成。荧光定量PCR 反应体系20 μL:模板cDNA 2 μL,上下游引物各0.4 μL,UltraSYBR Mixture(2×)10 μL,ddH2O 7.2 μL。荧光定量PCR的反应条件:94 ℃预变性10 s;94 ℃15 s,60 ℃60 s,40个循环。
1.4 遗传多态性分析
运用SHEsis 在线软件(http://analysis.bio‑x.cn/myAnalysis.php)计算基因型频率、等位基因频率、遗传杂合度(He)、有效等位基因数(Ne)、多态信息含量(PIC)以及哈迪-温伯格平衡(χ2)等。
1.5 肉质性状及肌肉组织特性测定
肉色、大理石纹、pH 值、失水率、肌内脂肪含量和剪切力测定参考文献[13]中的方法进行。将各肌肉组织样品修成1 mm3大小,置于2.5%戊二醛溶液中固定,经PBS 冲洗后,用锇酸固定,梯度乙醇脱水,包埋聚合后进行超薄切片,醋酸铀和柠檬酸铅双染色,进行透射电镜观察,测定肌原纤维直径、肌节长度和肌纤维密度。
1.6 数据处理
实时荧光定量PCR 中,采用2-△△Ct方法计算FoxO3基因的相对表达水平中,采用SPSS 22.0 软件分析FoxO3基因不同基因型对肉质性状的影响。
2 结果与分析
2.1 FoxO3 基因PCR扩增结果
每个样品分别加入FC+R、FT+R 混合物进行PCR 扩增,通过电泳图谱观察基因分型结果,FoxO3基因型为TT、CC和TC(图1)。

图1 FoxO3基因PCR扩增结果Fig.1 PCR amplification results of FoxO3 gene
2.2 FoxO3 基因PCR-SSP遗传多态性分析
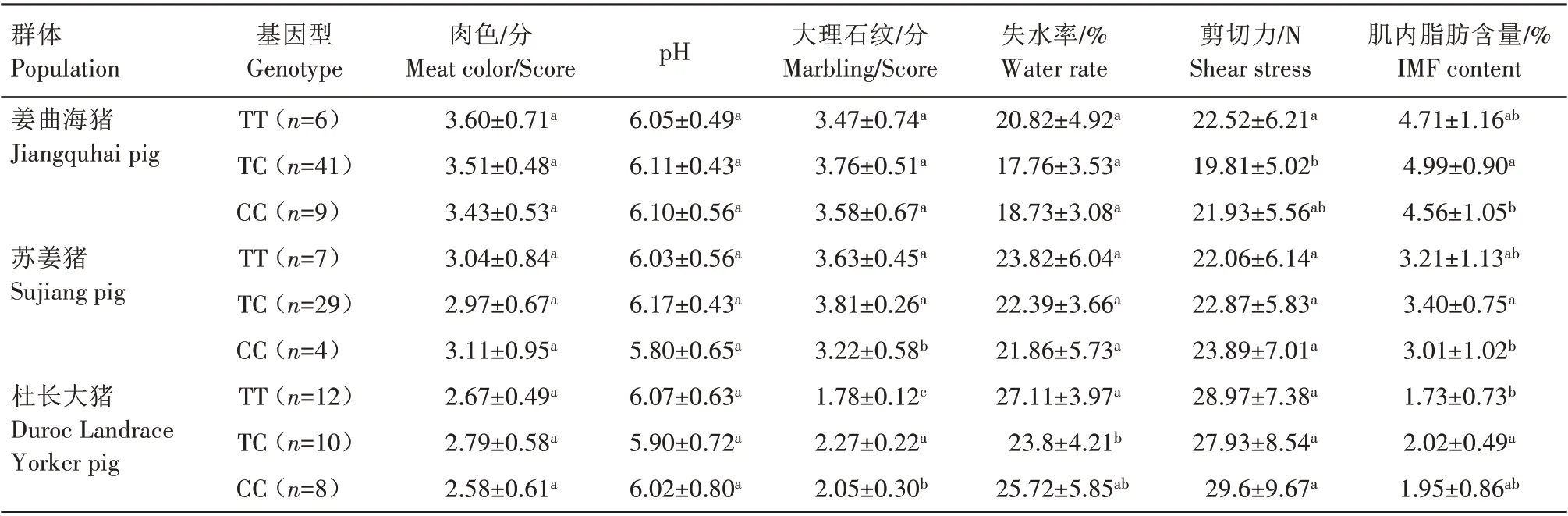
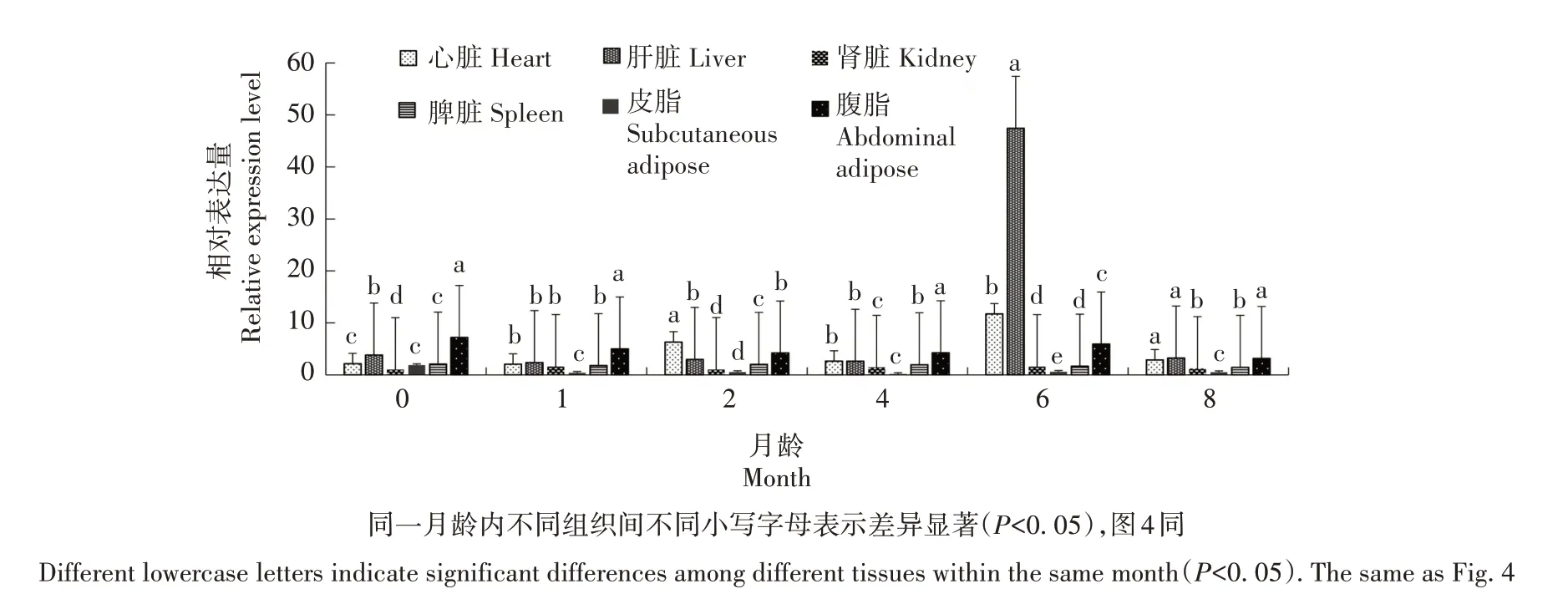
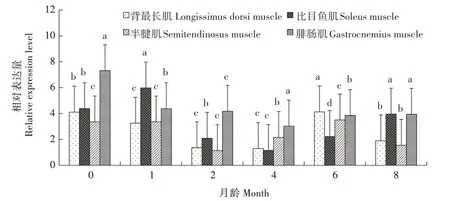
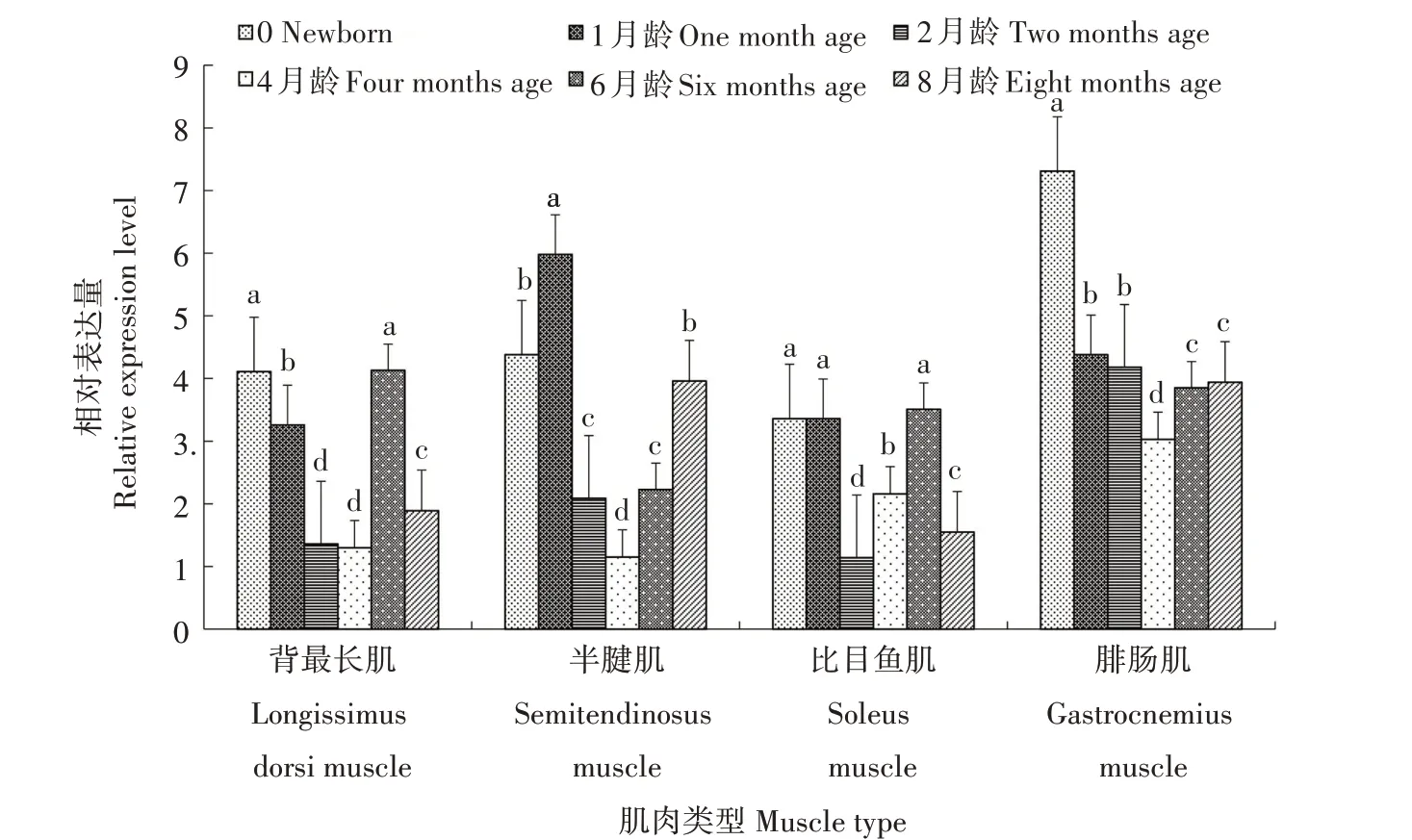
利用PCR-SSP 方法检测发现,FoxO3基因在姜曲海猪、苏姜猪和杜长大猪中均出现多态性(表1)。经χ2适合性检验,姜曲海猪和杜长大猪群χ2均小于5.99,表明群体基因频率和基因型频率均处于哈迪-温伯格平衡状态(P>0.05),苏姜猪未达到哈迪-温伯格平衡状态(P<0.05)。姜曲海猪、苏姜猪和杜长大猪群中T 基因的频率分别为0.424、0.538 和0.567,多态信息含量分别为0.369、0.374 和0.371,均属中度多态(0.25 表1 FoxO3基因的PCR-SSP多态性分析Tab.1 Polymorphism of FoxO3 gene by PCR-SSP 从表2 可以看出,FoxO3基因500 bp 位点多态性与姜曲海猪、苏姜猪和杜长大猪的大理石纹剪切力和肌内脂肪含量均有关联。姜曲海猪TC 基因型肌内脂肪含量显著高于CC 基因型,与TT 基因型差异不显著;TC基因型剪切力显著低于TT基因型,TT基因型与CC 基因型差异不显著。苏姜猪TC 基因型大理石纹得分和肌内脂肪含量均显著高于CC 基因型,TC 基因型与TT 基因型差异不显著;杜长大猪TC 基因型大理石纹得分显著高于CC 基因型,CC 基因型显著高于TT 基因型;TC 基因型肌内脂肪含量显著高于TT 基因型,TT 基因型与CC 基因型差异不显著。 表2 FoxO3 PCR-SSP基因型与肉质性状的关联性分析Tab.2 Association of FoxO3 genetypes by PCR-SSP in with meat quality FoxO3基因在姜曲海猪内脏和脂肪组织的差异表达见图2。初生时,FoxO3基因在腹脂的表达量最高,其次是肝脏,在肾脏的表达量最低;1月龄时,FoxO3基因仍是在腹脂的表达量最高,显著高于其他组织;2 月龄时,FoxO3基因在心脏的表达量最高,其次是腹脂和肝脏;4 月龄时,FoxO3基因在腹脂的表达量最高,其次是肝脏,在脾脏的表达量最低;6 月龄时,FoxO3基因在肝脏的表达量最高,其次是心脏,在脾脏的表达量最低;8 月龄时,FoxO3基因在肝脏的表达量最高,与在心脏和腹脂的表达量差异不显著,与在肾脏、脾脏和皮脂中的表达量差异显著。 图2 姜曲海猪FoxO3基因在内脏和脂肪中的表达差异Fig.2 Expression difference of FoxO3 gene in viscera and fat of Jiangquhai pig 姜曲海猪内脏和脂肪组织中FoxO3基因在不同发育时期表达差异见图3。在心脏中,FoxO3基因表达量在1 月龄时最低,6 月龄时达到最高;在肝脏中,FoxO3基因表达量1月龄时最低,除4月龄外,与其他各月龄均差异显著,6月龄时表达量达到最高值;在脾脏中,FoxO3基因1 月龄时表达量最高,与初生和2 月龄时表达量差异显著,6 月龄后表达量下降;在肾脏中,FoxO3基因1 月龄时表达量最高,与其他各月龄均差异显著。在皮脂中,FoxO3基因表达量在初生时最高,8 月龄时最低;在腹脂中,FoxO3基因表达量在初生时最高,8月龄最低。 图3 姜曲海猪内脏和脂肪组织中FoxO3基因在不同发育时期的表达差异Fig.3 FoxO3 gene expression difference in viscera and adipose tissue of Jiangquhai pig at different developmental stages FoxO3基因在不同骨骼肌中的表达差异见图4。初生时,FoxO3基因在腓肠肌中的表达量最高,其次是比目鱼肌,在半腱肌中的表达量最低;1 月龄时,FoxO3基因在比目鱼肌中的表达量最高,其次是腓肠肌;2 月龄时,FoxO3基因在腓肠肌的表达量最高,其次是比目鱼肌,在半腱肌中的表达量最低;4月龄时,FoxO3基因在腓肠肌中的表达量最高,其次是半腱肌,在比目鱼肌中的表达量最低;6 月龄时,FoxO3基因在背最长肌中的表达量最高,其次是腓肠肌,在比目鱼肌中的表达量最低;8 月龄时,在腓肠肌中的表达量最高,与背最长肌、半腱肌差异显著,与比目鱼肌差异不显著。 图4 姜曲海猪FoxO3基因在肌肉中的表达差异Fig.4 Expression difference of FoxO3 gene in muscles of Jiangquhai pig 姜曲海猪肌肉组织中FoxO3基因在不同发育时期表达差异见图5。背最长肌中,FoxO3基因表达量在6 月龄时最高,4 月龄时最低;半腱肌中,FoxO3基因表达量在1 月龄时达到最高值,与其他各月龄相比均差异显著,然后开始下降,4月龄时表达量达到最低值;比目鱼肌中,FoxO3基因表达量在2月龄时最低,与其他各月龄相比均差异显著;腓肠肌中,FoxO3基因表达量初生时最高,与其他各月龄均差异显著,随着月龄增加,表达量持续下降,4 月龄时表达量最低。 图5 姜曲海猪肌肉组织中FoxO3基因在不同发育时期的表达差异Fig.5 FoxO3 gene expression difference in muscle tissue of Jiangquhai pig at different developmental stages 8 月龄时,姜曲海猪背最长肌中FoxO3基因表达量与肌纤维直径、肌纤维密度、肌内脂肪含量和剪切力的相关系数分别是-0.518、0.525、-0.148 和0.084,FoxO3基因表达量与肌纤维直径和肌内脂肪含量呈负相关,与肌纤维密度和剪切力呈正相关,但相关性均不显著(P>0.05)。 8 月龄时,姜曲海猪FoxO3基因表达量与背最长肌、比目鱼肌、半腱肌和腓肠肌的肌原纤维直径呈负相关,相关系数分别是-0.574、-0.408、-0.406和-0.937,其中与腓肠肌肌原纤维直径呈显著负相关(P<0.05);FoxO3基因与背最长肌、比目鱼肌、半腱肌和腓肠肌肌节长度呈负相关,相关系数分别是-0.874、-0.445、-0.386和-0.628,其中与背最长肌肌节长度呈显著负相关(P<0.05)。 本研究首次利用PCR-SSP 检测了姜曲海猪、苏姜猪和杜长大猪FoxO3基因多态性,先将肉质差异较大的样品进行基因克隆测序,初步筛选SNP 位点,根据位点的实际情况,再选用适宜方法检测单核苷酸多态性,姜曲海猪、苏姜猪和杜长大猪中FoxO3基因均出现多态性,除了苏姜猪的FoxO3基因的各种基因型频率和基因频率经χ2检验未达到哈迪-温伯格平衡状态,其他各猪群的FoxO3基因的χ2均小于5.99,表明该群体基因频率和基因型频率都处于哈迪-温伯格平衡状态(P>0.05)。姜曲海猪FoxO3基因多态信息含量在0.25~0.50,属中度多态。一般认为,多态性越高的群体,其遗传多样性越丰富。由此可知,姜曲海猪的遗传多样性不及苏姜猪丰富,这可能与群体本身的遗传结构和近年保种有关。群体的遗传结构受群体大小、交配系统、祖先群体的多态性以及迁移、突变和选择等许多因素的影响。 FoxO 转录因子功能极其广泛,主要是由于FoxO 转录因子能够调节PI3K/AKT或PI3K/PKB信号通路中的下游基因,而这些基因与细胞生理过程,如细胞凋亡、DNA 损伤/修复、应激等有关。此外,FoxO 转录因子对脂肪分化、肌肉生成等过程中也发挥着关键性的作用[14‑15]。 NAKAO[16]和RATHBONE[17]报道了FoxO3基因通过上调p27基因抑制细胞的增殖,对巨核细胞与骨骼肌细胞的再生起负调节作用。本研究结果显示,FoxO3基因500 bp 位点多态性对姜曲海猪、苏姜猪和杜长大猪的大理石纹等分、剪切力和肌内脂肪含量均有影响,且FoxO3基因不同基因型间肉质性状存在较大差异。于晶[10]利用PCR-RFLP 检测了4 个地方猪种FoxO3基因第二外显子77 bp位点的多态性,结果表明,第二外显子77 bp 位点与胴体体斜长、6~7 肋间背膘厚、大理石纹存在显著关联,与失水率存在极显著相关。由此可见,FoxO3基因多态性与猪肉质有一定关联。 本研究发现,FoxO3基因在姜曲海猪初生、1 月龄、2月龄、4月龄、6月龄和8月龄时的皮下脂肪、腹脂、心脏、肝脏、脾脏、肾脏、背最长肌、比目鱼肌、半键肌和腓肠肌10 种组织中均有表达。姜曲海猪FoxO3基因在同一组织不同发育阶段的差异表达,可能与FoxO3基因在不同组织不同发育阶段的调控作用密切相关。MAMMUCARI 等[18]研究发现,FoxO3基因调控骨骼肌的蛋白质分解主要通过2 个途径,即泛素-蛋白酶体和自噬/溶酶体途径。综上,FoxO3基因表达对动物体骨骼肌的发育有着重要的调控作用。 本研究发现,FoxO3基因与肌纤维直径和肌内脂肪呈负相关,与肌纤维密度和剪切力呈正相关,但相关系数未达到显著水平;FoxO3基因与4 个部位肌肉肌原纤维直径和肌节长度均呈负相关,与腓肠肌肌原纤维直径、背最长肌肌节长度均呈显著负相关。王玲[11]研究发现,牛FoxO3基因表达量与肌纤维密度、面积、直径显著相关,其研究结果与本研究结果不完全一致,原因可能与畜种及月龄有关。 鉴于猪FoxO3基因表达、遗传多态性与肉质性状存在相关性,进而推断,在骨骼肌生长发育中,FoxO扮演了重要的角色。其主要原因是由于FoxO转录因子广泛参与生理代谢[19]。EELEN 等[20]的研究结果显示,FoxO3基因主要表达于骨骼肌中。祁样正[21]报道了在肌肉分化过程中,FoxO3基因表达量一直保持较高水平,表达量变化不明显,这可能与FoxO3基因对骨骼肌细胞的再生起负性调节作用有关。 受猪群规模限制,本研究的样本数目有限,使得遗传效应分析效果受到影响。后续研究中应继续扩大样本量,并在更多的猪群中开展深入分析研究,进一步探索FoxO家族基因对肌肉和脂肪性状所起的作用,寻找有效的分子遗传标记,加快优良猪种的培育。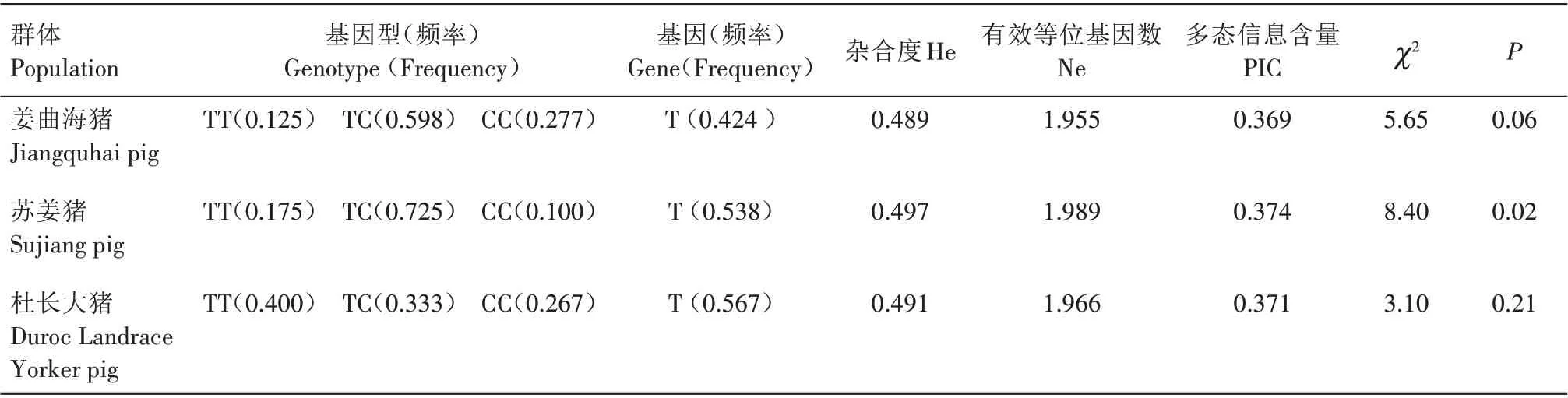
2.3 FoxO3 基因的遗传变异对肉质的影响
2.4 FoxO3基因在内脏和脂肪组织的表达差异分析
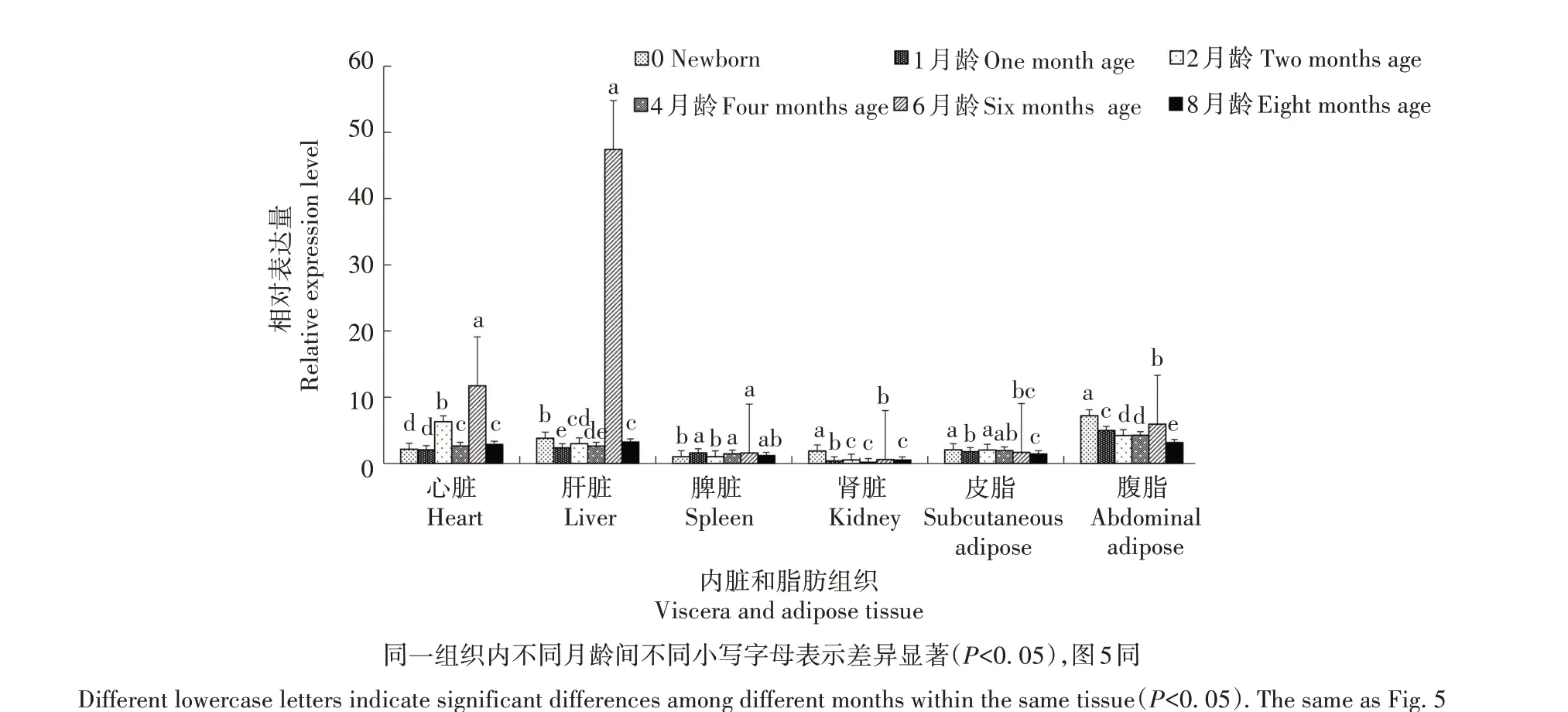
2.5 FoxO3基因在骨骼肌中的表达差异分析
2.6 FoxO3基因表达量与肉质性状相关性分析
2.7 FoxO3基因表达量与肌肉超微结构相关性分析
3 结论与讨论
