江苏宜兴龙池山自然保护区固定样地物种组成分析
2021-11-30伊贤贵谢春平彭智奇杨国栋董京京钟育谦翟飞飞王贤荣
伊贤贵,董 鹏,谢春平,彭智奇,杨国栋,董京京,钟育谦,翟飞飞,王贤荣
(1. 南京林业大学生物与环境学院,南方现代林业协同创新中心,亚热带森林生物多样性保护国家林业与草原局重点实验室,江苏 南京 210037;2.广东海洋大学滨海农学院,广东 湛江 524088;3.江苏省环境科学研究院,江苏 南京 210029;4.江苏省野生动植物保护站,江苏 南京 210036)
森林生物多样性的动态变化是了解生物与环境之间相互关系的纽带,建立固定大样地长期监测森林动态变化是生物多样性保护的有效途径,能真正准确了解生物进化过程、干扰机制及变化规律[1]。以大型固定样地为主的森林生物多样性监测受到越来越多的关注,为了解生物多样性的变化及其影响,理解物种共存机制等提供了翔实的数据[2]。国内外不同地区已经建立了许多不同类型的森林生态监测固定样地,以此掌握森林动态变化并评估人为、气候等干扰因素对森林的影响[3-5]。目前,我国已建成18个大型的固定监测样地,以及50多个辅助样地,覆盖了寒温带针叶林、温带针阔叶混交林、暖温带落叶阔叶林、亚热带常绿阔叶林、热带雨林等我国主要的地带性森林类型,形成了全球第一个具有完整纬度梯度的森林监测研究网络[6]。当前,已有学者对固定样地的森林群落特征、种间关联的空间分布格局、动态变化、物种共存机制等做了大量的基础研究工作,并获得了丰硕的成果[7-8],极大地促进了森林生物多样性的保护工作。
江苏省横跨纬度4°以上,处于暖温带和亚热带的过渡地带,植物分布上有明显纬度地带性分布和区系过渡性。由于受到全球气候变暖、人为干扰等因素的影响,江苏植被覆盖率呈下降趋势[9]。江苏宜溧低山丘陵区是我国常绿阔叶林分布的最北界,进行该区植被研究不仅对江苏省生物多样性的保护具有重要意义,而且可为我国不同地区常绿阔叶林的比较研究提供重要的基础材料。基于此,以江苏宜兴龙池山固定样地的调查结果为基础,分析该样地植被的物种组成、区系特征、优势种群、群落垂直结构和整体径级结构等,以期为当地生物多样性的保护、植被恢复策略的制定以及生态服务功能的评估等提供科学参考。
1 材料与方法
1.1 研究地概况
江苏宜兴龙池山自然保护区地处苏浙皖三省交汇处太湖西端的江苏宜兴市西南角(119°49′E, 31°22′N),面积为123 hm2,海拔50~480 m,为江苏省首批自然保护区。龙池山属低山丘陵,年平均气温15.7 ℃,1月平均气温2.9 ℃,7月平均气温28.3 ℃,无霜期平均达241 d,日均温在10 ℃以上的积温为 4 995.5 ℃;区域年降雨量1 200 mm左右,年均湿度为80%,属中亚热带湿润森林气候类型;土壤为石英砂岩母质上发育形成的黄棕壤[10]。该区以常绿阔叶林为主要保护对象,地带性植被保存较好。但受到人为干扰、旅游开发与管理体制等因素的影响,区内生物多样性保护受到了严重的威胁。保护区及周边地区约有种子植物124科521属845种,在区系分区上属泛北极植物区中的中国-日本森林植物亚区的华东地区[11]。同时,该区域也是华东地区分布最北的常绿阔叶林自然植被,物种的多样性高,是江苏南部地区生物多样性关键地区,具有较高的生物多样性保护价值[12]。保护区内有金钱松(Pseudolarixamabilis)、天目木兰(Yulaniaamoena)、银缕梅(Parrotiasubaequalis)等数种国家重点保护植物分布[13];同时壳斗科(Fagaceae)、樟科(Lauraceae)、山茶科(Theaceae)等亚热带常绿阔叶林的优势科在此均可见,如江苏地区较少的米槠(Castanopsiscarlesii)、红楠(Machilusthunbergii)等在此均有分布。
1.2 样地调查
2019年7—10月,在江苏省宜兴市龙池山选取本区域较为典型的地带性植被,建立1 hm2的森林动态监测固定样地。样地基准点地理坐标119°43′52.389 1″E, 31°14′35.358 7″N;海拔为280~320 m。样地建设标准参考GB/T 38590—2020《森林资源连续清查技术规程》及CTFS(center for tropical forest science)森林动态样地监测标准[14],首先利用NTS-A11R全站仪(南方测绘)将100 m×100 m的固定样地划分为25个20 m×20 m的样方,并将水泥桩固定在相应点,明确样方位置;其次,利用网格法将每个样方划分为5 m×5 m的小样格。在此基础上,对每个样格内出现的所有胸径(DBH)≥1 cm的木本植物进行每木调查,同时进行编号和挂牌;记录物种名、胸径、冠幅、坐标、生长状况等基本信息。同时,对每个样方的坡度、土壤、郁闭度、枯落物厚度等亦做相应的记录。
1.3 数据分析
1)物种组成与科属区系分析。对样地内出现的所有木本植物参考《江苏植物》(1~5卷)等植物志书进行植物种名称确定;裸子植物以郑万钧系统进行统计,被子植物以哈钦松系统进行统计。同时参考文献[15-16]对该群落的区系类型进行分析。
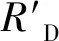
全文图表及数据统计均在Excel 2010中完成。
2 结果与分析
2.1 宜兴龙池山物种组成
对宜兴龙池山1 hm2固定样地植物群落的调查结果表明,该区域胸径(DBH)≥1 cm的木本植物共计有87种,隶属于40科66属[具体植物名录见本刊网络站(nldxb.njfu.edu.cn)或OSID附件)]。从属种组成来看(表1),含有4种以上的科有樟科、壳斗科、蔷薇科(Rosaceae)、大戟科(Euphorbiaceae)、漆树科(Anacardiaceae)和蝶形花科(Papilionaceae),其中樟科所含种最多达10种。余下科所含种数在1~3种,含有1种的科有23个,占所有科的67.50%;这说明物种组成较为分散,种类集中的优势科仅樟科、壳斗科等少数地带性科。从科的重要值分布格局来看,樟科、壳斗科和金缕梅科(Hamamelidaceae)的重要值最大,分别为16.77%、15.50%和14.99%;同时这3个科的相对多度也占有较大优势,分别为22.98%、15.45%和18.55%;这些数据说明它们在该固定样地中占有重要的地位。其次,重要值位列4至10的还有漆树科(Anacardiaceae)、安息香科(Styracaceae)、冬青科(Aquifoliaceae)、大戟科、榆科(Ulmaceae)、八角枫科(Alangiaceae)和山矾科(Symplocaceae)等;它们的重要值也均大于2.5%。上述10个科的重要值之和接近75%,同时它们的相对多度之和也达到了87.46%。因此,这10个优势科能够较好地反映样地的物种组成基本情况。
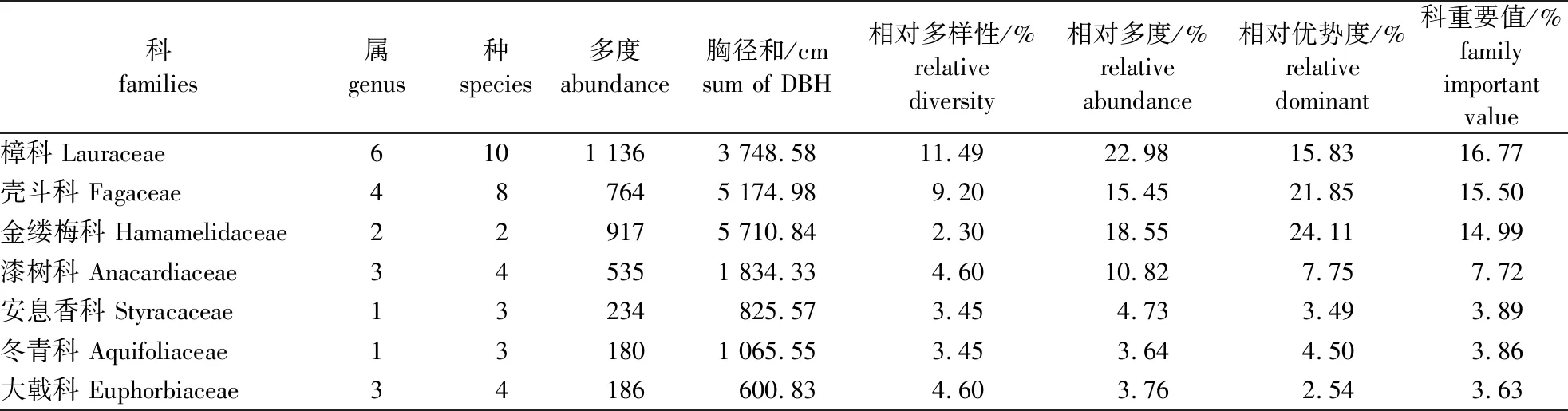
表1 宜兴龙池山1 hm2固定样地林木科属种组成及科重要值
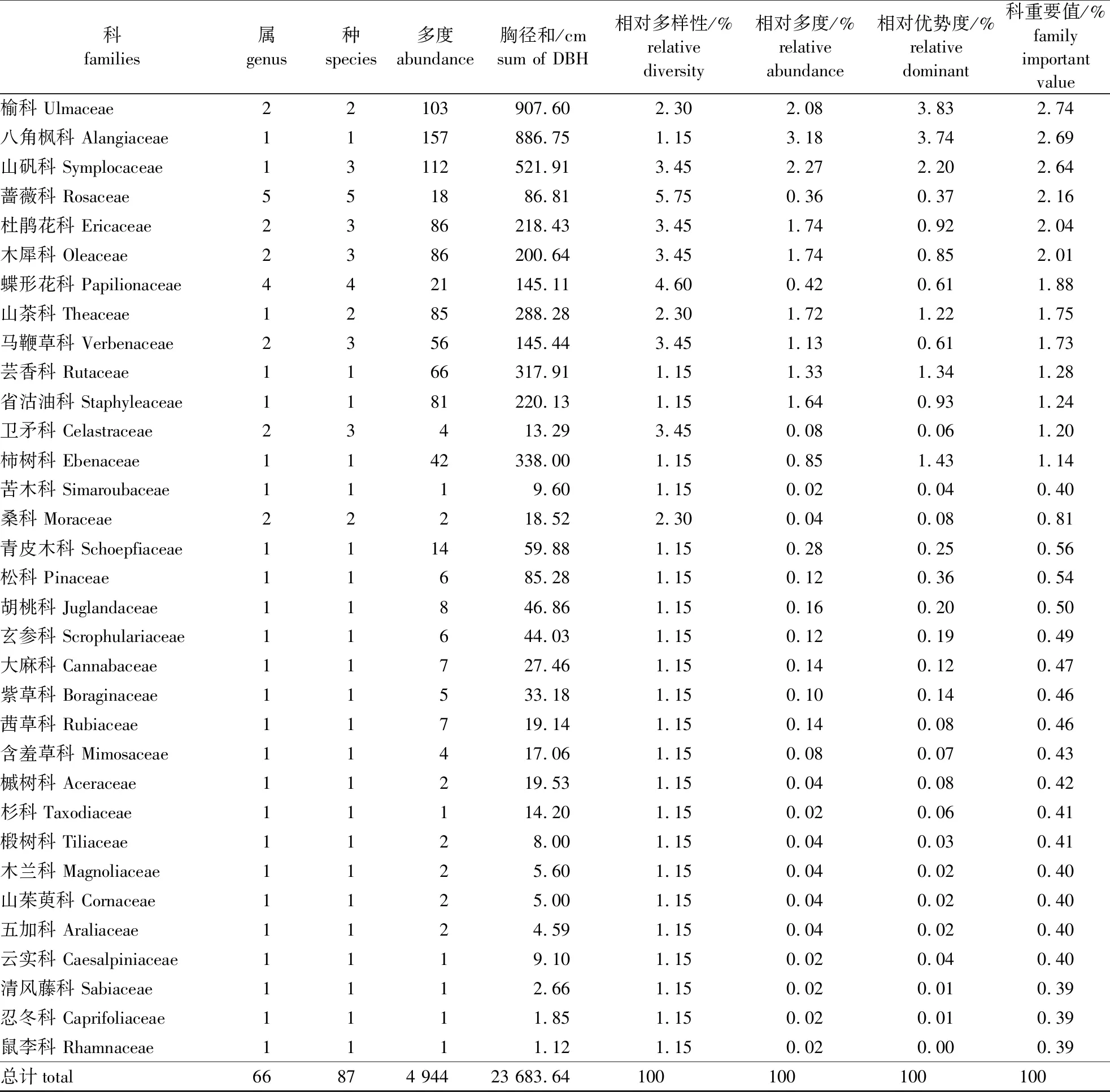
表1 (续)
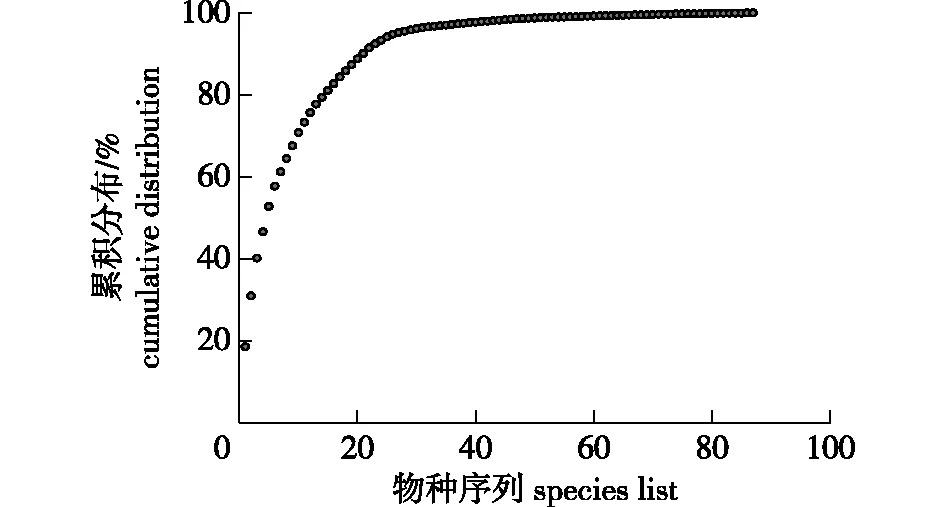
图1 宜兴龙池山1 hm2固定样地的物种多度累积分布图Fig.1 The cumulative distribution curve of species abundance in the 1 hm2 permanent plot of Yixing Longchi Mountain
在整个样地中,胸径≥1 cm的木本植物共计有4 944株,物种相对多度累计分布曲线如图1所示。从图1可知,在所有物种中个体数位列前10物种的累计相对多度值达70.73%;随着物种数量的进一步增加,个体数位列前15和20的物种的相对累计多度值分别达81.09%和88.77%。因此,可以看出样地内物种个体数的优势集中在前10个物种内;后续物种的个体数也越来越少,曲线趋于平缓。在这些植物种中,山鸡椒(Litseacubeba)、檵木(Loropetalumchinense)和野漆(Toxicodendronsuccedaneum)的个体数最为丰富,分别占群落内所有个体数的18.49%、12.38%和9.22%。根据Hubbell & Foster对稀有种、偶见种和常见种的物种数(S)定义,即稀有种为S≤1株/hm2,偶见种为1株/hm2
宜兴龙池山样地的种-面积曲线显示(图2A),物种数随着取样面积的增大而不断增加;当取样面积为0.1、0.2、0.3、0.4 hm2,物种数量分别为34、55、62和66种。当取样面积达到0.5 hm2时物种数累计出现已达80%(71种),而达到0.7 hm2时物种数已接近全部物种数的90%(75种);此时,曲线斜率已趋于平缓,物种数随取样面积增加无明显变化,说明该植物群落物种十分丰富。同时,后续增加的物种也多为稀有种或偶见种,他们在群落中的个体数也较为贫乏。此外,图2B所示的个体数-面积曲线的斜率基本保持一致,个体数随取样面积的增加而呈线性增长,这也说明了个体密度在样地中的分布较为均匀[18]。利用对数方程和一元线性方程对种-面积曲线和个体数-面积曲线进行拟合,拟合方程分别为:y=19.158 ln(x) + 84.19(R2= 0.969 9,P<0.01,F=3 154.008,df=1)和y=5 015.10x+ 299.53(R2=0.981 2,P<0.01,F=5 128.200,df=1)。
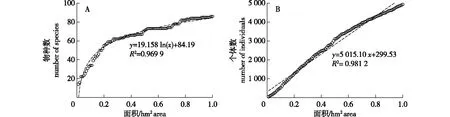
图2 宜兴龙池山1 hm2固定样地种-面积曲线(A)和个体数-面积曲线(B)Fig.2 The species-area curve (A) and individuals-area curve (B) in the 1 hm2 permanent plot of Yixing Longchi Mountain
整个样地中,重要值大于1%的物种共计25种,它们的重要值之和超过了90%;同时它们的相对多度和相对优势度也分别占94.09%和94.19%;因此这些物种能够代表及反映群落的整体情况(表2)。在该群落中重要值排在前3的分别为山鸡椒、檵木和枹栎(Quercusserrata),它们的重要值分别为12.65%、11.00%和7.89%。这3个物种在相对多度和相对优势度上均比其他物种占优,群落内接近40%的个体数均由它们组成。位列4至10的物种有野漆、牛鼻栓(Fortuneariasinensis)、苦槠(Castanopsissclerophylla)、冬青(Ilexchinensis)、瓜木(Alangiumplatanifolium)、野桐(Mallotustenuifolius)和野茉莉(Styraxjaponicus),它们的重要值分别为7.39%、5.83%、5.05%、4.08%、3.46%、3.32%和3.08%。同时上述10种植物的重要值也均大于3%,它们之和超过了60%,基本可以作为群落的典型代表种。但从上述这些物种可以看出,除苦槠、冬青为较典型的乔木外,其余物种多属小乔木或灌木的性质。重要值为2.00%~3.00%的物种有4个,其重要值之和为9.93%;重要值为1.00%~2.00%的物种有11个,其重要值之和为17.24%;余下物种的重要值均小于1.00%,重要值之和也仅为9.06%。因此,群落物种的重要值以山鸡椒和檵木最为突出。
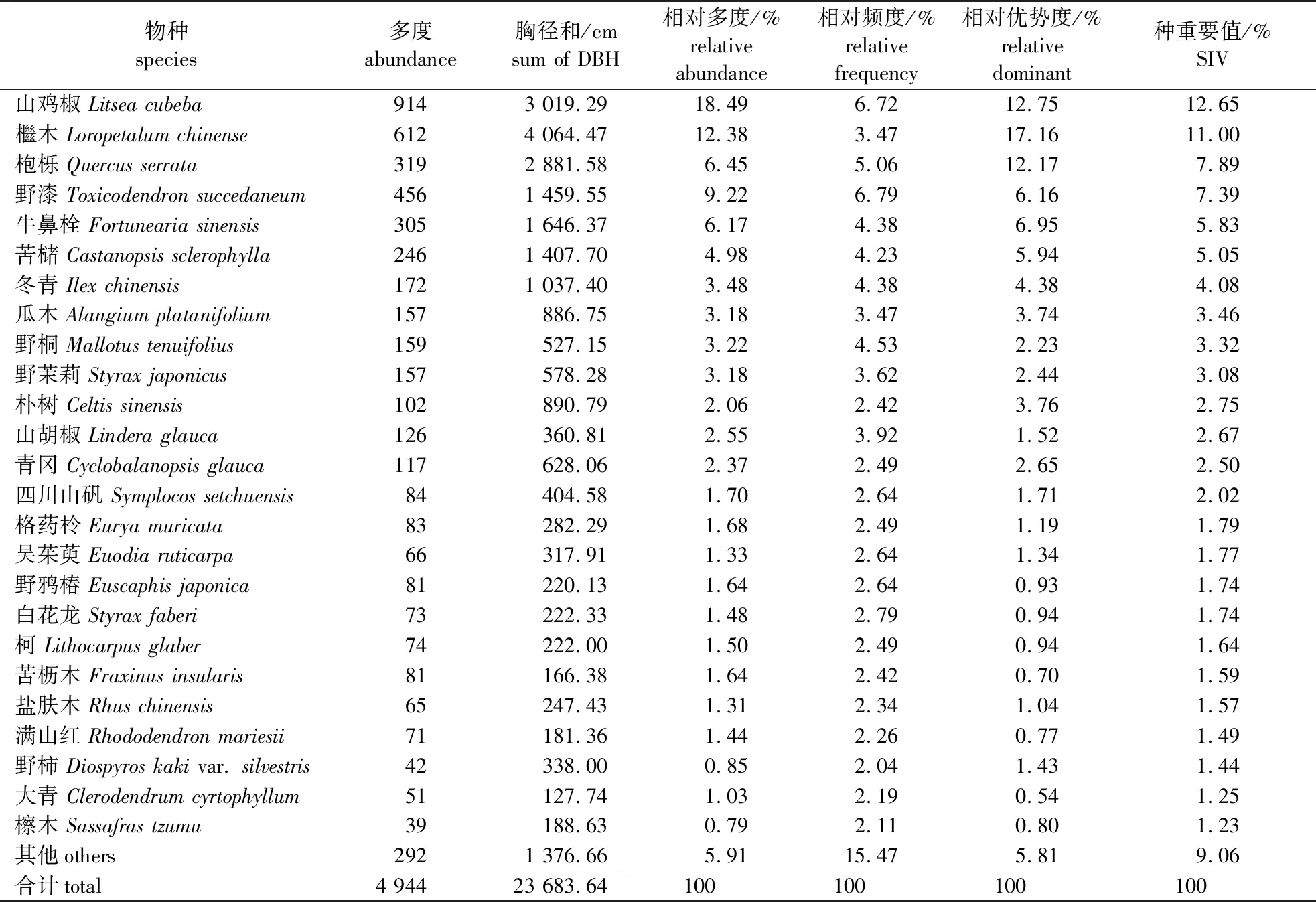
表2 宜兴龙池山1 hm2固定样地物种组成(种重要值大于1%)
2.2 群落结构特征
根据样地的实际情况将宜兴龙池山1 hm2固定样地群落的垂直结构大致划分为4层(图3A,3B)。乔木Ⅰ层为的树高≥10 m,主要有青冈(Cyclobalanopsisglauca)、苦槠、朴树(Celtissinensis)、黄檀(Dalbergiahupeana)等少数几个物种,他们的个体数所占比例大致接近2%。乔木Ⅱ层的高度为5~10m,它们不仅占整个群落50%左右的个体,而且构成了群落主体;这一层除上述几个物种继续出现外,还增加了野漆、黄连木、盐肤木(Rhuschinensis)、枹栎(Quercusserrata)、牛鼻栓、野柿(Diospyroskakivar.silvestris)等物种,该层次物种的多样性也较上一层高;同时由于该层的植物个体均已较为稳定地上升至群落中上层结构,其对资源的竞争和利用已经处于较为稳定的状态。乔木Ⅲ层的高度在5m以下,接近50%的物种均在该层(图3B);较为常见的物种有山鸡椒、山胡椒、檵木、白檀(Symplocospaniculata)、山矾(Symplocossumuntia)、野茉莉(Styraxjaponicus)、柯(Lithocarpusglaber)、满山红(Rhododendronmariesii)等物种。这一层不论是物种的个体数还是物种数均较上两层丰富;同时由于大量的个体均集中在这一层,植物的分枝交互重叠,这就导致种间的竞争也较前两层激烈。最下一层为林下更新层,该层除了乔木层的一些更新苗外,还有常见的灌木树种,如六月雪(Serissajaponica)、柃木(Euryajaponica)、茶(Camelliasinensis)等。草本层植物较为稀疏,常见类群主要以禾本科(Gramineae)、菊科(Compositae)及一些蕨类植物为主,如薹草(Carexsp.)、三脉紫菀(Asterageratoides)、沿阶草(Ophiopogonbodinieri)、黄精(Polygonatumsibiricum)、阔瓣鳞毛蕨(Dryopterischampionii)等。
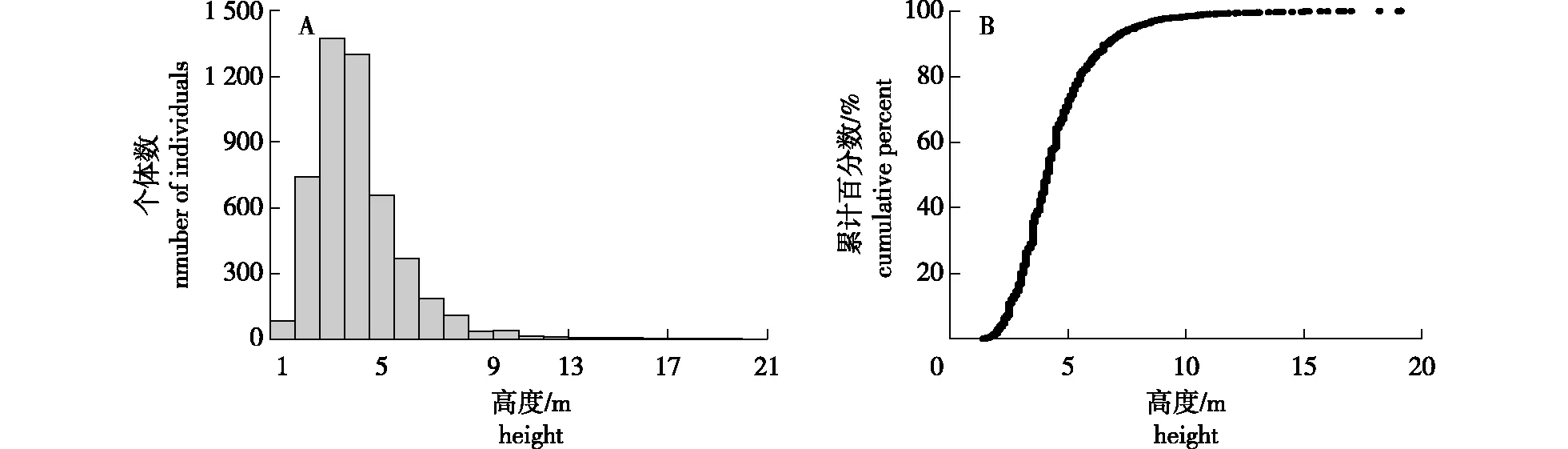
图3 宜兴龙池山1 hm2固定样地森林群落垂直结构分布(A)和高度累计分布图(B)Fig.3 The vertical distribution pattern (A) and cumulative distribution (B) of the community in the 1 hm2 permanent plot of Yixing Longchi Mountain
宜兴龙池山固定样地内所有DBH≥1 cm个体的胸径结构整体上接近倒“J”形(图4A),平均DBH为4.97 cm,最大DBH为40.4 cm;同时从图4B可以看出,曲线斜率较为陡直,超过70%的个体DBH在6 cm以下。将样地内所有木本植物的DBH划分为3个径级,即小径级木(1~6 cm)、中径级木(6~20 cm)和大径级木(≥20 cm),它们在群落中个体数所占比例分别为74.58%、24.13%和1.29%。因此,整体上随着径级的增加物种个体数量也逐渐减少,这说明了群落幼树个体较为丰富,群落的更新状态良好。
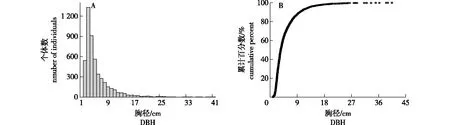
图4 宜兴龙池山1 hm2固定样地木本植物总体径级分布图(A)和径级累计分布图(B)Fig.4 The DBH size-class abundance (A) and cumulative distribution (B) of overall woody plants in the 1 hm2permanent plot of Yixing Longchi Mountain
2.3 植物区系特征
龙池山1 hm2固定样地木本植物的科、属区系类型的统计见表3。从科的区系类型组成来看,泛热带性质的科最多(12个),如樟科、漆树科、大戟科、卫矛科、山茶科(Theaceae)、山矾科(Symplocaceae)等。其次是世界广布和北温带性质的科同样具有9个,其中世界广布科有蝶形花科、蔷薇科、茜草科(Rubiaceae)、桑科(Moraceae)、鼠李科(Rhamnaceae)等;温带性质的科有金缕梅科(Hamamelidaceae)、胡桃科(Juglandaceae)、木兰科(Magnoliaceae)、山茱萸科(Cornaceae)等。未见中国特有分布科。从科的温热比值(9/21)来看,群落的热带性要强于温带,这与群落所属的亚热带北部地区的位置相符合。
属的分布区类型对植被的区系性质更具有代表性。从表3属的分布区类型来看,没有世界广布属存在。热带性质的属共有29属,其中泛热带分布的属所占比例较高达19.70%(13属),如合欢属(Albizia)、大青属(Clerodendrum)、算盘子属(Glochidion)、山矾属(Symplocos)、叶下珠属(Phyllanthus)等;紧随其后的是东亚及热带南美间断分布、旧世界热带分布和热带亚洲分布,分别有8属、4属和4属。温带分布的属共有34属,其中北温带所占比例最高达21.54%(14属),如栎属(Quercus)、杜鹃属(Rhododendron)、榆属(Ulmus)、松属(Pinus)等;其后还有东亚及北美间断分布所占比也较高达18.46(12属),如石楠属(Photinia)、漆属(Toxicodendron)、玉兰属(Yulania)、柘属(Cudrania)等。其他分布区类型还有旧世界温带、东亚等,但它们所含的属数及所占比例均较低。从属的温热比例(34/30)来看,温带性要比热带性略强,这表明了该植被区系具有从热带向温带的过渡性质。
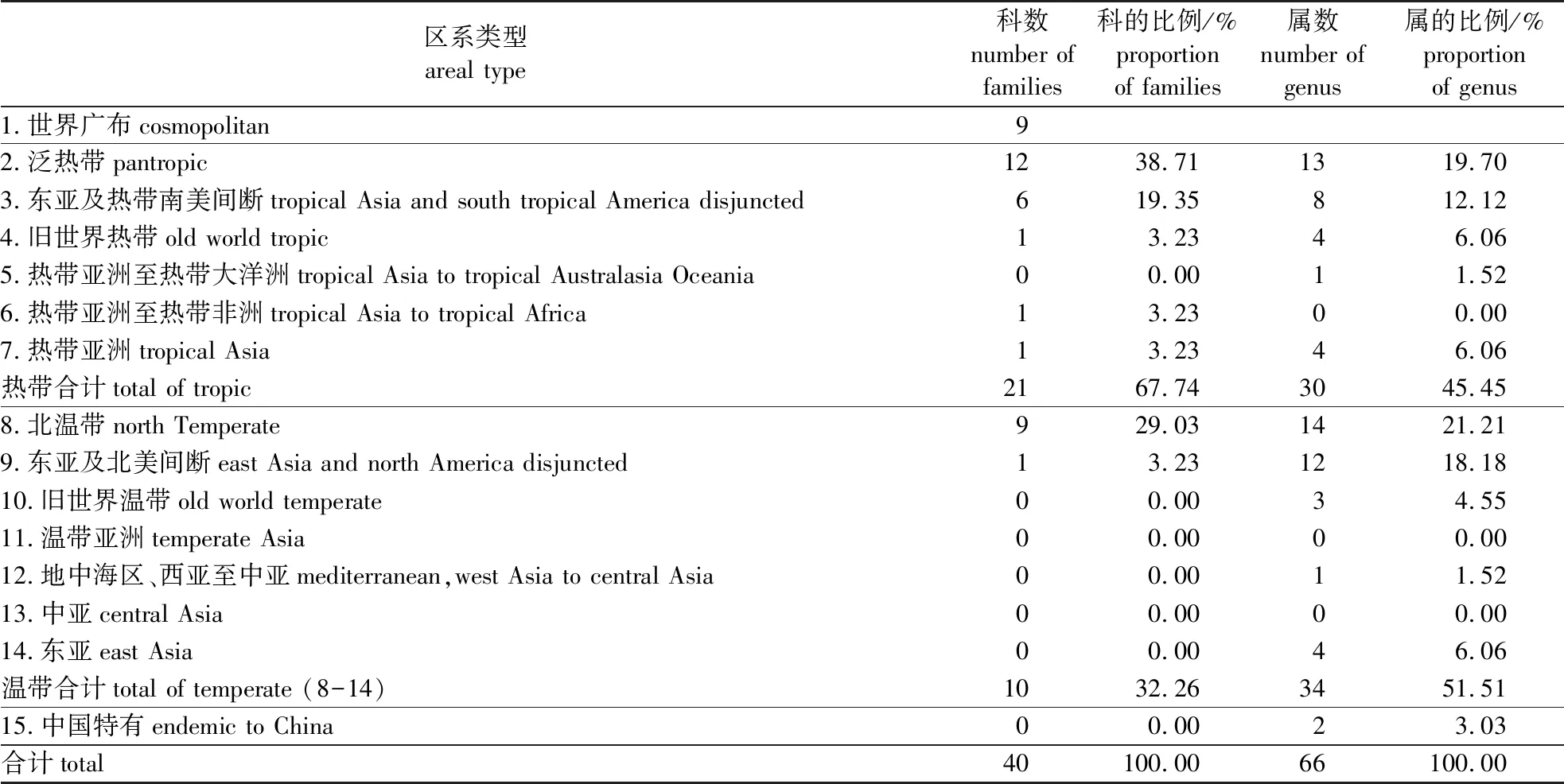
表3 宜兴龙池山1 hm2固定样地木本植物区系类型
3 讨 论
群落结构特征是植物在其生长过程中,由物种的生物学、遗传学特性、环境条件及物种相互关系等影响而形成的一种比较稳定的结构规律,可以为制定科学的保护策略与合理的经营措施提供有效依据[19]。短期森林群落调查是一种静态研究,仅能了解森林群落的现状;而固定样地的长期监测不仅为植物区系分析提供可靠依据,同时也为更好地了解种群特性与群落结构以及森林系统结构、功能和演化趋势提供了科学研究平台[1]。
1)群落物种多样性。植物组成和地理成分作为自然地理和历史条件在一定区域内共同作用的结果, 物种的构成中不仅能间接、稳定地反映一定区域内土壤、气候等自然条件特点[20],在物种丰富度、种群特性、物种更新和群落形成及其稳定性与演替规律的阐明中也具有重要作用。宜兴龙池山固定样地内物种多样性组成与其面积相近的其他1 hm2左右的固定样地比较,其物种数要多于江西大岗山低海拔常绿阔叶林(22科34属41种)[21]和宝天曼短柄枹林(27科47属71种)[22],而少于福建武夷山常绿阔叶林(34科65属139种)[23]。从水热条件来看,福建武夷山和江西大岗山要优于宜兴龙池山,但大岗山物种数要明显低于龙池山固定样地,这可能和群落演替的阶段有关。研究表明群落在演替早期,由于无明显优势种,物种的优势度较低,物种多样性较高;但随着演替的进行和优势度的增加,多样性变小;而随着群落结构的复杂化,多样性又将随着演替发展而增加[19]。因此,物种组成较好地反映了群落的不同演替时期和群落结构的复杂性。此外,从稀有种的情况看,龙池山固定样地的稀有种所占比例(17.24%)高于武夷山(16.55%)[23]而低于宝天曼(26.68%)[22];这有可能是龙池山和宝天曼群落均属于演替早期,物种进入群落的随机性较强。稀有种比例高的原因可能与物种本身特性、区系交汇、人为干扰等因素有关;同时,稀有种虽然在样地中个体数所占比例不大,但在维持生物多样性及森林生态系统功能的稳定中起着重要作用[24]。
2)群落区系过渡性强。植物区系特征对于了解一个区域植被及植物群落的分布、演替、环境变迁以及植物的进化均有着重要的意义[25]。宜兴龙池山固定样地科的热带性类型分布和温带性类型分布分别占比例为67.74%和32.26%;但同时世界广布科有9个科,占所有科的22.50%。虽然世界广布科无法体现某一地区的具体植被类型,但世界广布科所占比例较大则说明了该植被具有一定的次生性。但是,从属的分布区类型来看,温带性所占比例为51.51%要略强于热带性的45.45%。因此,综合科属区系分布类型,可以判断该群落区系具有热带区系与温带区系交汇融合的特点。武夷山固定样地不论在科或属的分布类型上均是热带强于温带,而宝天曼样地区系则表现为温带强于热带的特点。因此,宜兴龙池山固定样地的区系特征较好地体现了其所在的地理位置。
3)地带性优势种明显。从重要值大于1%的物种来看,多数物种都是北亚热带森林常见物种,如枹栎、野漆、牛鼻栓、瓜木野桐等;而山鸡椒、檵木、苦槠、冬青、青冈等物种更倾向于中亚热带分布。因此,从优势物种的组成结构也可以看出该区域植被的过渡性与复杂性。同时,山鸡椒、檵木、枹栎等物种的重要值占据较大的比重,但其实是以量取胜,其相对多度较为丰富;而且这些物种从本质上更倾向于小乔木或灌木的性质,无法形成乔木上层的主体。值得关注的是,当前群落乔木层优势树种中存在落叶和常绿树种混生的情况,如常绿树种苦槠、青冈、冬青等,落叶树种枹栎、黄连木、野漆等;一方面说明了当前群落为落叶常绿混交林的属性,另一方面也说明了群落常绿阔叶林的演替方向。此外,从样地优势物种的构成可以看出,目前群落内落叶树种的比例要高于常绿树种。这是因为,当前落叶物种具有更高的资源养分利用效率,能够利用夏季良好的水热条件以迅速获得碳水化合物完成高生长,在与常绿树种的竞争过程中居于优势地位[26]。随着群落的进一步演替,群落郁闭度增加,喜阳的落叶树种在林下层与耐阴的常绿树种的竞争将失去优势地位,群落将逐渐演替成为以常绿树种为主的群落特征。
4)群落垂直与径级结构。森林群落的垂直结构体现了不同树种的高度,反映了不同植物在群落中的分层结构和生态位[27];物种高度结构在森林群落的变化使得光、热、水等均发生相应的改变,从而使得物种在各层交互错开。本研究将群落划分为4层,依次为乔木Ⅰ层(≥10 m)、乔木Ⅱ层(5 m~10 m)、乔木Ⅲ层(<5 m)和灌木层,而群落物种的平均高度为4.5 m;因此乔木Ⅰ层和Ⅱ层构成了群落的上层结构,这些物种在上层光资源的竞争中占有较大的优势。在乔木Ⅰ层和Ⅱ层的树种中,青冈、苦槠、朴树、黄檀、野漆、黄连木、盐肤木、枹栎等所占比例较高。不难看出,野漆、黄连木、盐肤木、枹栎等均为北亚热带和南暖温带地区常见的落叶树种,而且它们也是在次生林早期演替中最容易入侵群落的。但是,结合群落林下层更新苗情况发现,大量的常绿阔叶树种的幼树幼苗存在。虽然这些落叶树种目前仍占有较大的优势,但随着群落的演替,其当前的优势地位将逐渐被常绿树种所取代,径级结构是植物群落稳定性和生长发育状况的重要指标[8],本研究群落所有个体的径级结构呈倒“J”形,这说明了中小径级个体数量要多于大径级的个体数量,同时反映出群落的稳定性及更新情况良好。径级结构与树高结构近似,均是随着量级的增加个体数逐渐减少,当达到一定量级后个体数趋于平稳,这反映出群落物种的高度与胸径呈现一定的相关性。同时,小径级个体数较多可能是南方森林或者演替中森林的一个普遍特点;这可能是种群、种间竞争激烈,物种更替频繁,而群落处于受到一定程度干扰后的恢复演替阶段[28]。常绿阔叶树种对生态条件要求较高,不管是种子出苗或萌蘖再生,其更新能力都远比落叶阔叶树种弱;一旦落叶常绿阔叶混交林遭受严重破坏,原来林内的常绿阔叶树种恢复困难,特别是生态环境剧变后,其很长时间内都难以恢复[29]。
本研究综合群落的物种组成、区系特征与径级结构等发现,宜兴龙池山固定样地的植被由落叶与常绿阔叶混交林朝着常绿阔叶林的方向演替。本研究仅就当前调查的数据进行了统计分析,对群落物种多样性及结构组成影响的因素还有很多,如地形、土壤、小气候、种间关系、不同物种的生理生态适应性等;在社会经济发达地区减少人为干扰和生境破坏,是保护当前群落健康发展及物种多样性的关键。以固定样地为研究平台有待后续研究。在社会经济发达地区减少人为干扰和生境破坏,是保护当前群落健康发展及物种多样性的关键。深入探讨不同生态机制对森林群落维持的作用,将更有利于科学合理地制定有效的生物多样性经营、管理与保护策略。
