黄果厚壳桂人工林幼树群体生存状态及谱分析
2022-01-21黄松殿向宇繁陈碧珍邓祥胜李明蔓
黄松殿,向宇繁,陈碧珍,邓祥胜,何 鹏,李明蔓,程 飞,田 湘
(1.广西壮族自治区南宁树木园,广西南宁 530032;2.广西大学 林学院,广西南宁 530004)
黄果厚壳桂(Cryptocarya concinna)是樟科(Lau⁃raceae)厚壳桂属常绿树种,主要分布于中国的广东、广西、江西和台湾等地,是南亚热带季风性气候典型的植被类群之一。黄果厚壳桂的木材纹理美观、木材颜色鲜明、干形饱满通直、材质坚硬且韧且分枝少[1-2],可作为建筑材料和家具原料,具有很高的栽种和研究价值。
由于分布区域的限制,目前对黄果厚壳桂的研究主要集中在广东和广西种群。张池等[3]采用He⁃gyi 指数模型分析广东省自然保护区黄果厚壳桂林分的种内竞争,发现黄果厚壳桂种群的竞争强度与林木径级、林木间距离有关。李林等[4]对黄果厚壳桂种群的死亡动态格局进行监测,发现死亡数量明显大于个体新增数量,需严防重度干扰对于该种群的影响。张咏梅等[5]用成对双样本均值分析的方法对锥栗(Castanea henryi)-荷木(Schima superba)-黄果厚壳桂群落进行对比,了解该群落在时间上的变化。彭少磷等[6]发现黄果厚壳桂是我国季风性气候地区常绿阔叶林的优势种群,也是部分南方地区地带性林分的建群种。这些研究均以天然林为研究对象,对黄果厚壳桂人工林的更新特征仍需进一步了解。
种群数量变化的规律和动态模型需要对该物种种群进行统计,主要体现方式是生命表。编制静态生命表要对物种种群年龄的分布频率在一段特定时间内取样,将数据进行统计并描述。静态生命表可反映种群出生率和死亡率随年龄变化的规律。从静态生命表里可解读出种群结构,以生命表为基础可对濒危植物采取措施,改变对目标种群的管理策略[7-9]。
谱分析可将一系列的数据从时域转换到频域进行分析,有效地研究这一系列数据是否存在周期性,从而清楚地观察并了解这段数据的动态特征。所有的植被都存在波动性,谱分析可揭示种群数量变动的周期性波动。通过谱分析来观察种群数量的动态变化,对濒危植被的保护有重要意义,还可为种群的群落演替、稳定性评价、物种多样性保护和开发利用提供理论依据。
本研究以广西良凤江国家森林公园内的黄果厚壳桂人工林天然更新的幼树群体为研究对象,统计幼树地径的径级数量分布,编制静态生命表,并进行谱分析,研究黄果厚壳桂人工林幼树群体的数量变化特征,为更好地了解黄果厚壳桂人工林的变化规律和林木生长特性提供参考,为黄果厚壳桂资源的开发利用、种质资源保护提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于广西南宁市南郊的良凤江国家森林公园内(108°15'~108°22'E,22°34'~22°46'N),地处北回归线以南,属亚热带季风性气候,冬季较短,夏季较长。全年日照充足,≥10 ℃年积温约7 600 ℃,年均气温21.6 ℃。干湿季节分明,雨季集中在4— 9月。地形以丘陵地貌为主,也有少量的阶地;土壤呈酸性,多为棕红色或砖红色,由砂页岩发育而成,有机质含量较少。
1.2 调查方法
该黄果厚壳桂人工林于1981年引种种植,面积约0.2 hm2,郁闭度在0.8 以上,林下有较多天然更新的幼苗和幼树。2020年8月下旬,对黄果厚壳桂人工林进行全面踏查。在能够代表林分特征和立地条件相似的地段设置3 个10 m×10 m 的样方,样方之间间隔5 m。采用卷尺(精确至0.01 m)和测高器测量样方内每株幼树的高度,用游标卡尺(精确至0.01 mm)测量幼树地径(距离地面高度10 cm 处)。幼树定义为树高大于30 cm 且胸径小于5 cm 的个体。
1.3 分析方法
1.3.1 生命表的编制
本研究生命表的编制用径级结构替代年龄结构[10]。本研究共获得713 株黄果厚壳桂幼树数据,以2.5 mm的地径为起测径级,每增加5 mm为1个径级,可将幼树地径分为6个径级,即Ⅰ(2.5 mm ≤D<7.5 mm)、Ⅱ(7.5 mm ≤D<12.5 mm)、Ⅲ(12.5 mm ≤D<17.5 mm)、Ⅳ(17.5 mm ≤D<22.5 mm)、Ⅴ(22.5 mm ≤D<27.5 mm)和Ⅵ(D ≥27.5 mm)。生命表中各项参数的计算公式如下[10-11]:
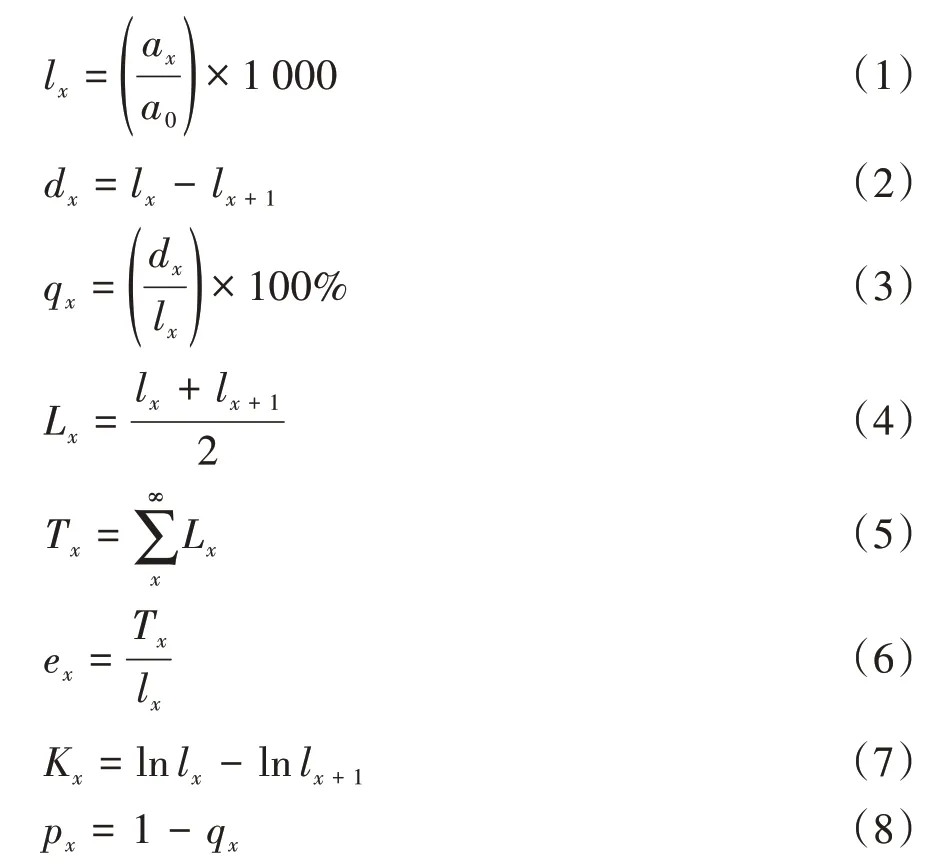
式中,x为径级;ax为在x径级内的幼树个体数;a0为第Ⅰ径级内的幼树个体数;lx为第x径级的标准化存活个体数;lnlx为标准化存活个体数lx的对数值;dx为从x到x+1 径级间隔期内的死亡数;qx为从x到x+1 径级间隔期内的死亡率;Lx为从x到x+1 径级间隔期内的存活数;Tx为从第x径级到超过第x径级的个体数;ex为第x径级个体的平均期望寿命;Kx为消失率;px为从x到x+1径级间隔期内的存活率[11]。
1.3.2 曲线绘制及分析
以径级(x)为横坐标、死亡率(qx)和消失率(Kx)为纵坐标绘制黄果厚壳桂幼树群体死亡率和消失率曲线;以径级为横坐标、标准化存活个体数(lx)的对数值(lnlx)为纵坐标绘制存活曲线。
存活曲线可以直观地表达出幼树群体的存活过程。Deevey 存活曲线分为3 种类型[12]。Deevey-Ⅰ型又称为A型,曲线整体呈凸型,表示该幼树群体在初期时,幼体死亡率非常低,绝大多数个体都能存活至平均寿命,当达到平均寿命时,短期内大量个体死亡;Deevey-Ⅱ型又称为B 型,曲线整体呈对角直线,表示该群体各径级的死亡率相等;Deevey-Ⅲ型又称为C型,曲线整体呈凹型,表示该群体在初期时死亡率高,随着径级的增长,死亡率逐渐降低且趋于稳定。
采用SPSS 统计软件对黄果厚壳桂幼树群体的存活曲线进行拟合,检验其是否符合Deevey-Ⅱ型或Deevey-Ⅲ型[13],建立的数学模型如下:
Deevey-Ⅱ型:

Deevey-Ⅲ型:

式中,Nx为第x径级内个体的存活数;N0为该群体形成初期的个体存活数;b为死亡率。
1.3.3 生存分析
为了更好地分析黄果厚壳桂幼树群体的生存规律,引入4种函数对其进行生存分析,生存率函数S(i)、积累死亡率函数F(i)、死亡密度函数f(i)和危险率函数λ(i),计算公式如下[14]:

式中,pi为存活率;qx为死亡率;hi为径级宽度。
根据4 个生存函数的计算结果绘制生存率曲线、累计死亡曲线、死亡密度曲线和危险率曲线。
1.3.4 群体数量预测
运用时间序列分析一次移动平均法预测黄果厚壳桂幼树群体的年龄结构,公式如下[15]:

式中,t为径级;n为需要预测的时间;Mt为经过n个径级时间后t径级种群存活数的预测值;Xk为第k径级的种群存活数。本研究将预测经过2、3、4、5和6个径级时间后各径级的存活数。
1.3.5 谱分析
谱分析可用于研究林木种群年龄结构更替过程的周期性及波动性,是Fourier 级数的展开,可以写成正弦波形式[16]:
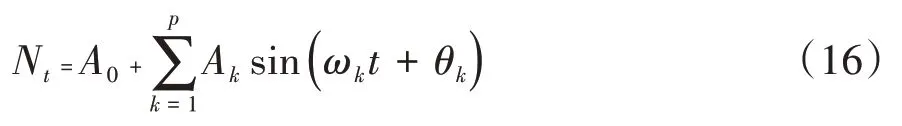
式中,A0为周期变化的均值;Ak为各谐波振幅值;ωk和θk分别为谐波频率和相角;Nt为t时刻种群大小。
谱分析正弦波形式中的各个参数由下列数学模型得出:
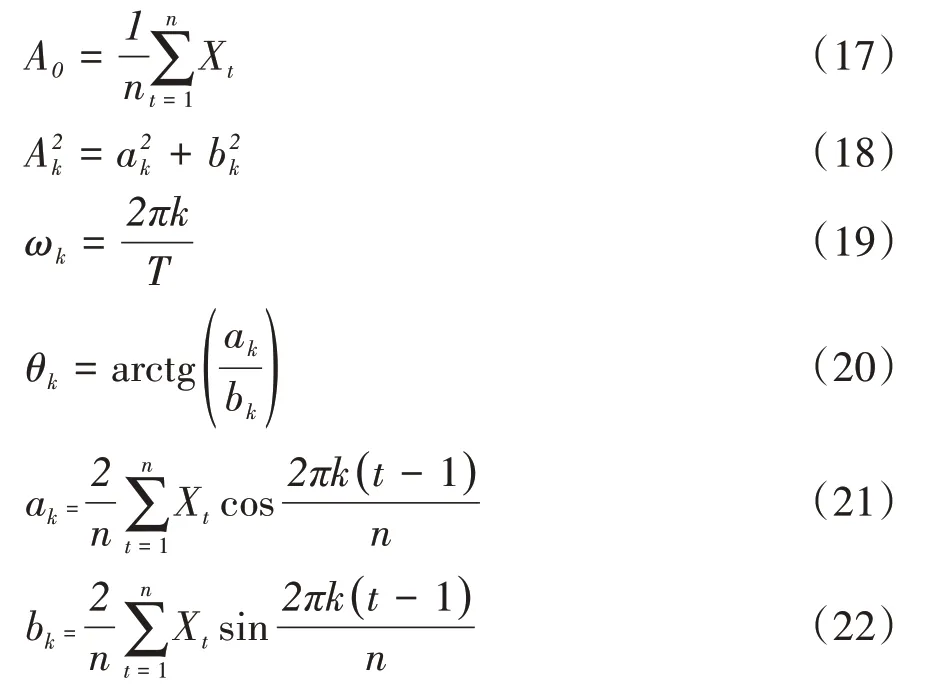
式中,ak和bk为参数估计值;将种群各径级个体数分布视为时间序列t,Xt表示t年龄序列时的个体数;n为系列总长度;P=n∕2为谐波的总个数;T为正弦波的基本周期,即时间序列t的最长周期,即资料总长度,即T=n为已知。
本研究中,黄果厚壳桂幼树群体共有6个径级,将群体各径级个体数分布视为时间序列t(t= 1,2,3,4,5,6);系列总长度n为6;谐波的总个数为P=n∕2=6∕2=3,需计算振幅值Ak(k=1,2,3);T=n=6。谐波对应径级的计算方法如下:

式中,D为地径;n为径级总数;hi为径级宽度;k= 1,2,3。通过D值所处的范围得出谐波对应的径级。
由于黄果厚壳桂幼树群体各径级个体存活数相差较大,在利用谱分析公式进行计算时采用对数处理,用Xt'代替公式中的Xt(即为表1 中第x径级内的幼树个体数ax)[17]。处理后对各波形的振幅值Ak(k=1,2,3)进行计算。
2 结果与分析
2.1 径级结构
黄果厚壳桂幼树群体的径级结构呈倒金字塔型。各径级个体存活数表现为Ⅰ>Ⅱ>Ⅲ>Ⅳ>Ⅴ>Ⅵ(图1)。第Ⅰ径级的个体数最多,占总数的38.43%;其次为第Ⅱ径级,占总数的31.98%;第Ⅲ~Ⅵ径级共占总数的29.59%。随径级增加,幼树个体数减少。
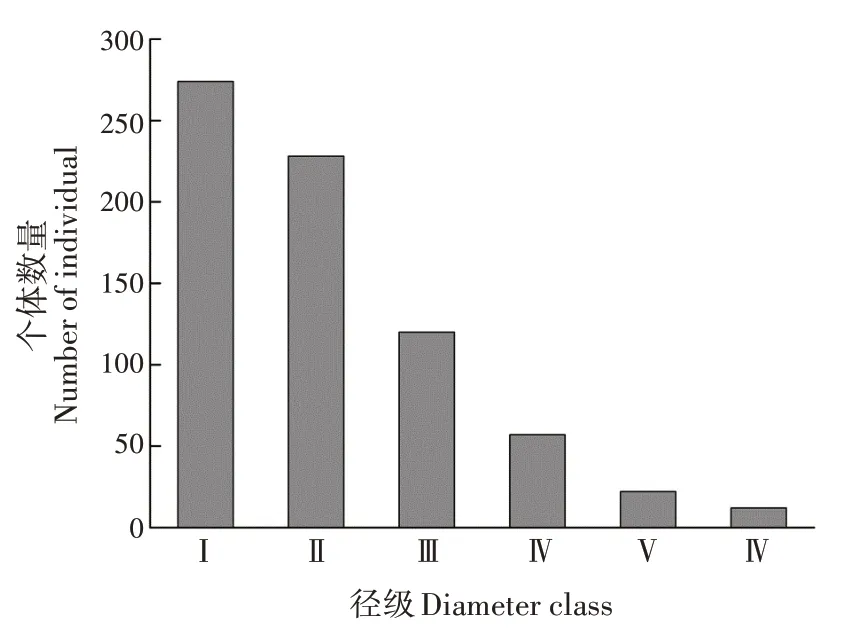
图1 黄果厚壳桂幼树群体径级结构Fig.1 Diameter class structure of C.concinna sapling popu⁃lation
2.2 静态生命表
黄果厚壳桂幼树群体中,第Ⅰ、Ⅱ径级的ax和lx较大,分别为274、1 000 和228、832;第Ⅵ径级的ax和lx最小,分别为12和44(表1)。ax和lx均随径级增大而减小,原因可能是黄果厚壳桂幼树群体中大部分个体年龄较小。
dx呈现明显的波动性,第Ⅱ径级最大(394),第Ⅴ径级最小(36),表明在第Ⅱ~Ⅲ径级间隔期内死亡数最多,在大径级的间隔期内死亡数最少(表1)。原因可能是随径级增加,幼树个体在生长过程中受种内竞争或环境等因素的影响而被淘汰,变成中、大径级的个体数量减少。

表1 黄果厚壳桂幼树群体静态生命表Tab.1 Static life table of C.concinna sapling population
ex反映各径级的平均生存能力。ex呈下降趋势,第Ⅰ径级最大(2.10),第Ⅵ径级最小(0.50)。在第Ⅰ至Ⅱ、第Ⅳ至第Ⅵ径级,ex明显降低。黄果厚壳桂幼树耐阴喜阴[18],林冠下的小径级个体更易存活,但由于种内竞争,群体生存能力下降;大径级个体因为生理原因生存能力变差,少数大径级个体较早分布在林分中,占据了更好的生长空间[19],没有被淘汰。
2.3 幼树群体曲线
2.3.1 死亡率和消失率曲线
qx和Kx的变化趋势基本一致,存在一定波动但整体呈增加的趋势,符合一般的种群规律(图2)。qx和Kx在第Ⅰ径级均最小,在第Ⅳ径级达到峰值后均下降,说明第Ⅰ~Ⅳ径级的幼树间竞争逐渐加剧或生存环境逐渐恶化,第Ⅳ径级后竞争减缓或生存环境改善。
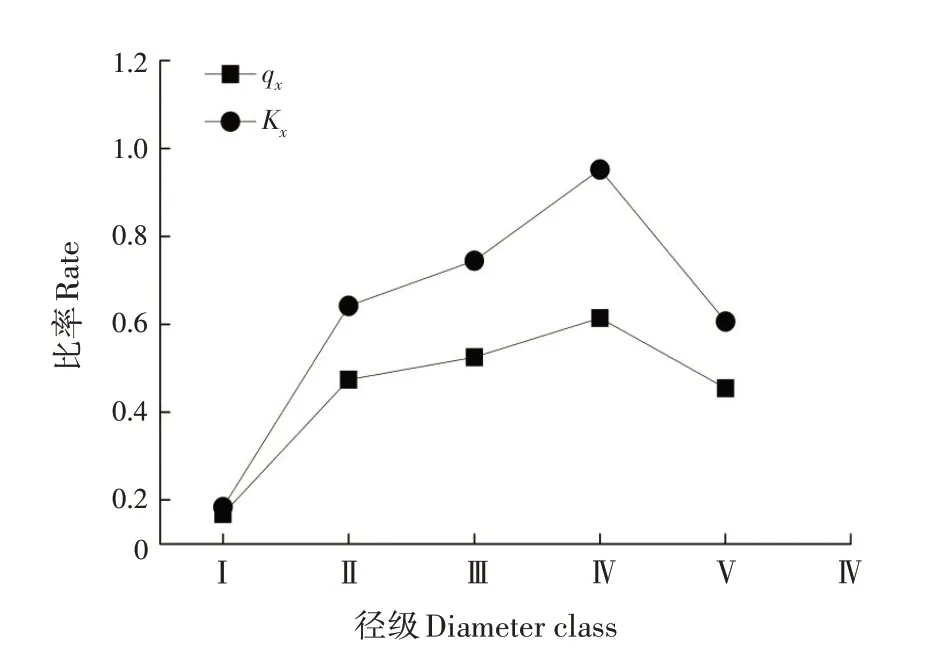
图2 黄果厚壳桂幼树群体死亡率(qx)和消失率(Kx)曲线Fig.2 Curves for mortality rate and vanishing rate of C.con⁃cinna sapling population
2.3.2 存活曲线
存活曲线呈近乎线性下降趋势(图3)。Deevey-Ⅱ指数函数模型的R2= 0.97,Deevey III 幂函数模型的R2= 0.84,说明指数函数模型的拟合效果好,优于幂函数模型(表2)。黄果厚壳桂幼树群体的存活曲线属于Deevey-Ⅱ型,表明该群体死亡率相对稳定。

表2 黄果厚壳桂幼树群体存活曲线的检验模型Tab.2 Test models of survival curves of C.concinna sapling population

图3 黄果厚壳桂幼树群体存活曲线Fig.3 Survival curve of C.concinna sapling population
2.3.3 生存函数曲线
黄果厚壳桂幼树群体的F(i)呈单调上升,S(i)呈单调下降,两者互补(表3)。在接近第Ⅱ径级时,F(i)等于S(i),说明在第Ⅱ径级之前群体达到平衡状态,随后F(i)超过S(i),表明群体进入衰减期。群体在第Ⅰ~Ⅲ径级有相对较高的ex,但个体密度大,生长竞争激烈,导致大批个体在这3个径级被淘汰,使f(i)较高;第Ⅳ径级后,f(i)趋于平缓,源于个体间竞争趋于缓和,淘汰减少。λ(i)在第Ⅴ径级出现顶峰,说明第Ⅴ径级幼树死亡风险最高,这与生理性死亡等原因有关。从幼树群体数量上看,小、中径级数量多但减少快,大径级数量少但减少趋于平缓,幼树群体整体呈衰减趋势。
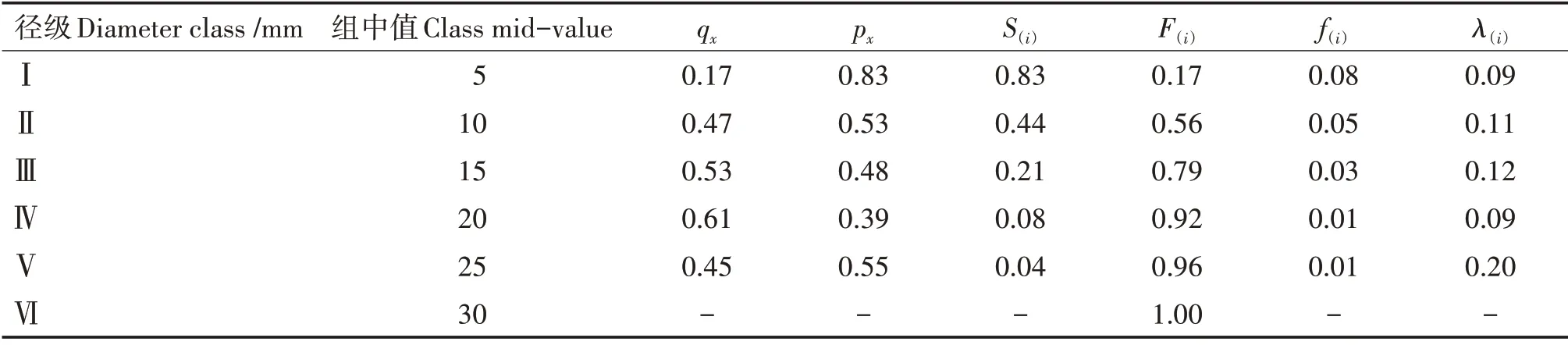
表3 黄果厚壳桂幼树群体的生存函数估算值Tab.3 Estimated values of survival functions of C.concinna sapling population
2.3.4 群体数量的时间序列分析
以黄果厚壳桂幼树群体各径级个体数量为基础,采用时间序列分析的方法预测该群体第Ⅱ~Ⅵ径级后的数量(表4)。除了第Ⅱ径级在两个径级后数量下降近一半外,其他各径级均呈增长趋势,第Ⅲ径级增长最明显,说明该群体的小、中径级个体数量充足,短期内占群体主导地位。第Ⅲ径级后,主要为大径级,整体上仍处于数量增长期,与小径级数量充足有关。整体上看,各个径级个体数量在之后的各径级阶段均上升,但在经过6个径级后,群体存活数由713 个下降至119 个,说明该黄果厚壳桂幼树群体属于衰减型群体。
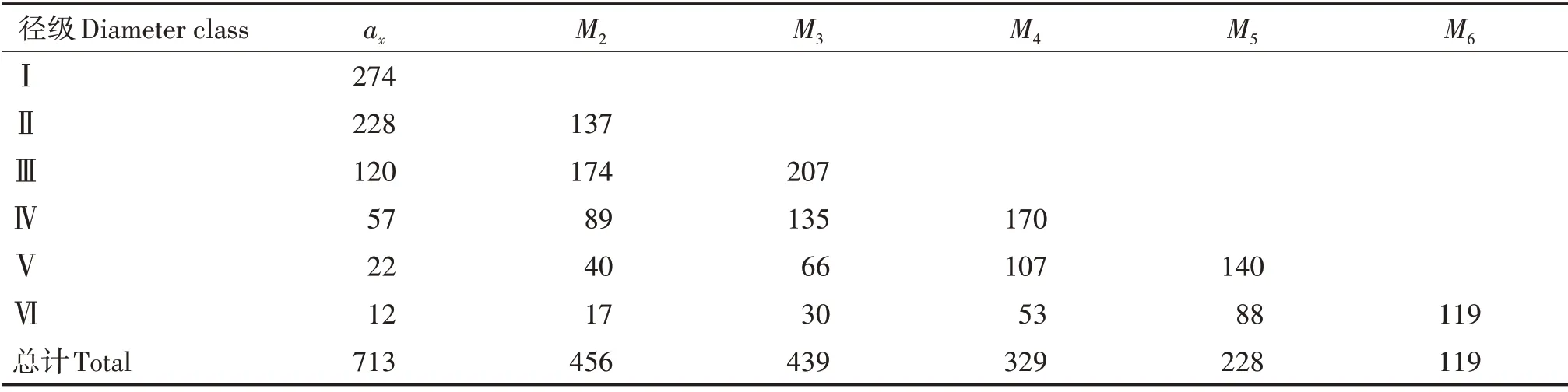
表4 黄果厚壳桂幼树群体动态变化的时间序列分析Tab.4 Analysis of time series of C.concinna sapling population
2.4 谱分析
A1至A3的振幅呈单调递减的趋势,各谐波的振幅相差较大,表明该群体数量变化的波动并非单一周期,即大周期内包含小周期,且各周期的作用强度不同(表5)。谐波A2和A3对应的周期分别是基波A1的1∕2 和1∕3,整体上随着周期的增大而增大[20]。基波的周期取决于黄果厚壳桂幼树群体的特性,能够表现出该群体数量变化的波动性。除去周期变化的平均值(A0)后,基波A1振幅值最大,表明群体的数量变化受基波的影响十分显著。

表5 黄果厚壳桂幼树群体的周期性波动Tab.5 Periodic fluctuation of C.concinna sapling population
黄果厚壳桂幼树群体的数量动态除受基波的影响外,还表现出小周期波动。第1 次小周期为谐波A2,反应在6 × 5∕2 = 15 mm,位于空间序列径级12.5~17.5 mm 处,对应第Ⅲ径级,这一小周期波动与幼树的生理特性或林分的环境有关。第2次小周期也是理论上的最小周期(A3),反映在6×5∕3=10 mm,位于空间序列径级7.5~12.5 mm 处,对应第Ⅱ径级,这一小周期波动与幼树间的激烈竞争导致部分个体死亡有关,这种数量调节更有利于群体的稳定发展。
3 讨论
种群是构成生物群落和生态系统的基本单位,幼树的群体结构可反映种群的发展趋势。研究林分特定种群的幼树结构特征和数量变化规律,对于林分的可持续经营有重要意义。了解幼树群体的存活率和死亡率等变化规律,是研究种群的基本任务之一[21-23]。
径级结构可体现种群的结构特征和稳定性,静态生命表和存活曲线从宏观角度观测群体数量的变化规律。黄果厚壳桂人工林幼树群体的径级结构呈倒金字塔型,各年龄结构较为完整,由于黄果厚壳桂的耐荫特性,小径级个体在林冠下更易存活,这与小径级ex较大相一致;由于个体密度大,随径级增加,个体间竞争加剧,群体生存能力逐渐下降,被淘汰个体也逐渐增加[24],少数大径级个体占据更好的生长空间而生存下来。该幼树群体在第Ⅳ径级时qx和Kx出现峰值,这是由于在第Ⅳ径级,种内竞争和环境筛选导致幼树在生长发育至高径级过程中死亡[25]。存活曲线呈下降趋势,说明林冠下的荫蔽环境有利于小径级个体存活;吴敏等[26]发现光照因素可能导致幼苗难以生长发育为幼树,对黄果厚壳桂人工林的管理应充分考虑该树种的光适应性,为其提供适当的遮荫环境,保证小径级苗木的生长发育。存活曲线属于Deevey-Ⅱ型,随径级增大,个体存活数量逐渐减少,说明该幼树群体拥有较多的小径级个体和少量的中、大径级个体,群体构成较为稳定[27]。在小径级阶段,种内竞争激烈,生存空间或资源不足,自疏或它疏作用造成淘汰,仅保留少部分发育至大径级。从生存函数来看,幼树群体在小、中径级时期的生存压力主要来自于种内竞争,大径级时期的生存压力主要来自于环境筛选。通过时间序列预测可知,该幼树群体小径级个体数多,未来有较多幼树可发育成大径级。对于该林分的经营重点应放在小径级幼树的管理上,比如改善小径级个体的种内竞争。随着种群个体数的增加,相邻的个体间会相互影响(密度效应),通过合理的措施调控密度效应,可以改善幼树群体的生存状况。
谱分析表明,该幼树群体的数量变化明显受生命周期生物特性因素的影响[28],且在数量变化的大周期内有一部分小周期。从整体来看,该群体仍以小径级黄果厚壳桂幼树个体为主,未进入成熟、衰老的阶段。谱分析结果只能体现当前幼树群体生长情况的变化规律,不能体现黄果厚壳桂种群整体在整个基本周期的数量变化规律。
