吕梁山濒危植物珊瑚兰资源调查及形态解剖学研究
2021-09-11杨阳曾令天景梦岳
杨阳 曾令天 景梦岳


摘要:调查研究吕梁山野生珊瑚兰生存现状、资源分布及花芽分化的形态解剖学特征,为建立吕梁地区濒危植物珊瑚兰种植资源库和保育性开发提供依据。采用野外调查、路线踏查并辅以典型样地取样的方法对吕梁山野生珊瑚兰资源进行实地保护性调查;并用常规实体解剖石蜡切片法,显微观察珊瑚兰花芽分化及发育的过程。结果表明,野生珊瑚兰分布于111°13′10″~111°13′15″E,37°5′37″~37°5′31″N之间,海拔高度影响野生珊瑚兰分布。中性土壤和 9~20 cm土壤腐殖质能够为珊瑚兰植株生长提供一个稳定的生长环境。珊瑚兰的花芽形态特征与普通兰花保持一致,即由3片外轮花被片、2片内轮花被片、1个唇瓣、1个合蕊柱构成,在合蕊柱顶端有花药、蕊喙,底端为子房,特立中央胎座着生胚珠。研究发现,吕梁山有野生珊瑚兰资源分布,海拔高度、土壤腐殖质显著影响野生珊瑚兰居群分布。
关键词:吕梁山;濒危植物;珊瑚兰;资源调查;形态解剖;花芽分化
中图分类号: S682.310.1 文献标志码: A 文章编号:1002-1302(2021)16-0122-05
野生珊瑚兰(Corallorhiza trifida Chat.)是兰科珊瑚兰属植物,多年生腐生草本,非光合兰花,濒危(EN)级,保护等级Ⅱ级。在我国,珊瑚兰主要分布在西北、四川等地区[1-2]。近年来,在北京门头沟百花山发现有珊瑚兰生长,成为北京新记录的植物之一[3]。珊瑚兰属约有13个种,多数种原产于美洲,1个种原产于亚欧大陆[4]。珊瑚兰是腐生植物,无叶,没有叶绿素,无法像常见的绿色植物一样进行光合作用。它们依靠一些真菌进行共生,以此来获取土壤中的养分[5]。珊瑚兰对土壤要求较高,要求腐殖质丰富、没有污染的林下环境或灌木环境[6-8]。珊瑚兰作为一种腐生植物,生长方式较常见绿色植物大为不同,在增加植物多样性方面有至关重要的作用[9]。珊瑚兰在生长发育过程中,会与共生菌形成2个十分关键的时期,一是开花结果期,二是种子萌发及幼苗建成期,这2个时期共生菌极大地影响珊瑚兰的生长情况。虽然其种子数量巨大,然而种子胚发育不完全,没有胚乳,在自然条件下种子必须有特定真菌为其提供营养才能成功萌发,随后建立稳定的共生关系[10],经查阅文献可知,珊瑚兰对共生菌具有极高的特异性[2]。McKendrick等研究发现,珊瑚兰有外生菌根真菌从其他植物中获取营养并传输给珊瑚兰供其生长[11]。蒋玉玲等对珊瑚兰菌根真菌多样性做过研究[2],这为珊瑚兰根的研究提供了一些研究基础,但对于珊瑚兰花芽的研究,鲜有报道。为研究珊瑚兰的发育过程,须要对珊瑚兰的花芽进行解剖研究。采用石蜡切片法[12-13],研究花芽的解剖结构和花芽发育进程,为研究珊瑚兰提供形态解剖学依据。同时,珊瑚兰作为濒危植物,生存状况极不乐观,保护性研究刻不容缓。
此外,兰科植物一般分布于温暖、湿度大、通风和具散射阳光的环境,兰科植物的生境特点决定了我国兰科植物地理分布不均匀,主要集中在长江流域及以南地区。比较而言,由于我国北方地区兰科植物多样性较低,科研人员开展调查和保护工作主要集中在兰科植物的多样性中心即西南地区,很少关注资源相对匮乏的北方地区。但对不同地区的兰科植物开展调查和进行多样性保护研究,可以保护不同的基因型和变异,为兰科植物种质基因库的建设提供更多选择。吕梁地区位于华北地区山西省的中部西侧,境内有森林覆盖率较高的山区,生物资源雄厚,符合兰科植物的生境特点。但记录该地区的野生兰花的文献资料较少,开展的相关工作匮乏,野生兰花种质资源总体分布状况不明,特别是野生珊瑚兰在吕梁地区的分布状况几无报道,所以尽快对吕梁地区濒危植物珊瑚兰种植资源进行调查分析和保育性开发显得尤为重要。
1 材料与方法
通过提前查阅资料确定吕梁地区交城县庞泉沟、中阳县上顶山和离石区吴城九凤山等5个有代表性的,具备兰科植物生境特点的样地,采用野外调查、样本采集与鉴定、整理、分类、编目相结合,选线与跑面相结合,专业踏察与当地群众了解访问相结合的方式,根据不同地区、山体的生态条件、植被特征及其自然环境等状况,采取路线踏查、抽样调查、典型样地调查等相组合的方法,进行吕梁地区野生珊瑚兰种质资源调查。调查工作从2016年4月上旬开始,2019年7月结束。调查内容包括居群分布情况、生长环境、生物学特征、生殖过程和种群发育情况等,并对调查结果进行分析。
采样分析生境、物候期特征和生殖生物学特征,包括伴生植物、坡度、土壤结构、物候期(萌动、抽茎、花期、果期、枯萎)。野外用甲醛-乙酸-乙醇(FAA)固定液采样,用4 ℃ 冰盒取回,通过固定—冲洗—脱水—透明—浸蜡—包埋—切片—粘片—烘片—染色封片,显微镜观察珊瑚兰的花芽分化及形态特征,其中切片厚度为7.5 μm。
2 结果与分析
2.1 野生珊瑚蘭居群分布情况分析
2016年4月至2019年7月经过多次踏查,拟踏察的庞泉沟自然保护区的八水沟、八道沟、福禄沟,离石九凤山、白马仙洞景区等5个调查地均未发现野生珊瑚兰种群。在地理位置为111°13′9″E、37°5′37″N的中阳县上顶山发现有野生珊瑚兰种群分布。
2017年4月开始对中阳县上顶山野生珊瑚兰居群分布情况进行详细调查。按照不同的海拔高度和经纬度,选取代表性区域作为典型样地,每个样地10 m2,重复3次记录数据。随海拔高度变化踏查,开始发现野生珊瑚兰群落的海拔高度为 1 820 m,经度为111°13′10″E,纬度为37°5′37″N。
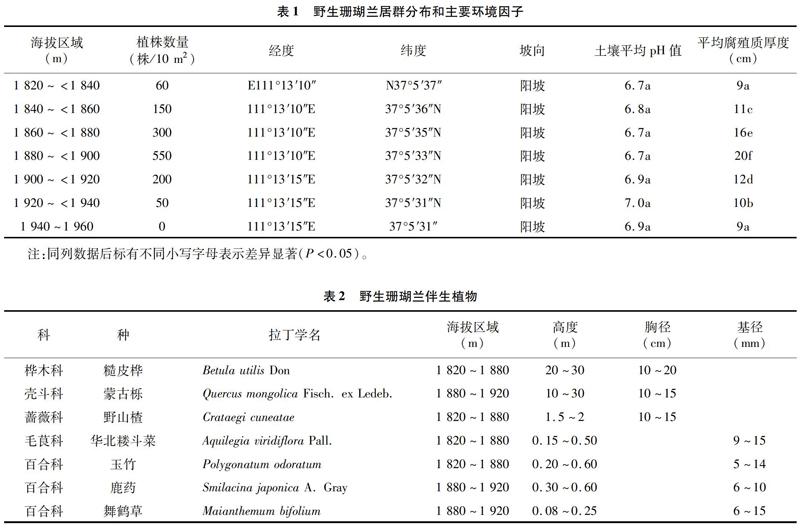
由表1可知,上顶山野生珊瑚兰群落分布于111°13′10″~111°13′15″E,37°5′31″~37°5′37″N,海拔高度为1 820~1 940 m,坡向为阳坡。样地野生珊瑚兰植株根际附近土壤平均pH值为6.7~7.0,平均腐殖质厚度为9~20 cm。土壤平均腐殖质厚度由9 cm升高至20 cm时,植株数量由平均 60株/10 m2 升高至550株/10 m2;当土壤平均腐殖质厚度降低至9 cm时,植株数量下降至0株/10 m2。可以看出,海拔高度、土壤平均腐殖质明显影响野生珊瑚兰居群分布。
2.2 野生珊瑚兰伴生植物分析
伴生植物与野生珊瑚兰分布有着紧密联系,样地调查中,随着海拔高度的提升,野生珊瑚兰伴生植物也发生了变化(表2)。海拔为1 400~1 800 m的区域,多为华北落叶松、白皮松、油松、白桦、野山楂、山荆子、杜梨、暴马丁香、蔷薇、车前、舞鹤草、玉竹、华北乌头、藜芦、瓣蕊唐松草、芍药等。海拔为 1 820~<1 940 m的区域,乔灌木层可分为10~30 m 的大中型乔木,主要是糙皮桦、蒙古栎;还有1.0~3.0 m的灌木,主要是野山楂、蔷薇;草本层植物分布比较密集,主要包含了玉竹、鹿药、华北耧斗菜、舞鹤草等。海拔为1 940~1 960 m的区域,乔木层植物主要为山杨树、糙皮桦,蒙古栎几乎没有,草本层植物有高山野罂粟、东方草莓、水杨梅等。
2.3野生珊瑚兰生物学特征
野生珊瑚兰为总状花序,植株平均高度为10.43~13.41 cm,花葶高度为10~13 cm,每株小花数量平均4~7朵。海拔高度影响株高和小花数量。海拔为1 880~< 1900、1 900~<1 920 m的区域,野生珊瑚兰平均株高分别为12.07、13.41 cm,明显高于其他海拔的植株;平均小花数量为7朵,也明显高于其他海拔的植株(表3)。
野生珊瑚兰植株根状茎,直径为1.5~3.0 mm,呈珊瑚状,白色,多分枝[14]。海拔为1 860~<1 900 m的区域,野生珊瑚兰平均根状茎直径为3.0 mm,明显高于其他海拔的植株(图1)。
2.4 野生珊瑚兰花芽分化及解剖结构分析
野生珊瑚兰成年以后,在吕梁山特殊环境因素的影响下完成成花诱导,形成花器官原基,由营养生长期进入生殖生长期[15]。本试验研究其花芽分化及解剖结构,为兰科植物种质资源保护工作奠定基础。
试验发现,珊瑚兰的花器官形态符合典型兰花形态标准,由内到外依次为合蕊柱、内轮花被片、外轮花被片,内外轮花被片均为3片,其中一片内轮花被片特化为唇瓣,这也是蝴蝶兰[16]、寒兰、文心兰[17]等兰花的特有现象。珊瑚兰的花蕾呈梭形,外由花萼包被,长3~6 mm,宽1.0~1.5 mm。花被下方有绿色梭形鼓囊,外有长条形棱,是珊瑚兰花的子房,长2~6 mm,直径为1.5~2.5 mm。花被片长度与子房长度基本符合1 ∶ 1的长度比例。
珊瑚兰花芽分化开始于珊瑚状根状茎生长点,花序原基最先分化,生长点突起,花原基伸长,细胞质浓,细胞小(图2-1)。花序原基继续分化,进入顶花芽花被原基分化期。顶花芽在雄蕊原基分化期,分化出雄蕊原基,整体明显向外突出,由内到外依次为雄蕊、花被、苞片(图2-2、图2-3)。还分化出雌蕊原基,由内到外依次为雌蕊、花柱、雄蕊、2层花被、花苞(图2-4、图2-5)。
珊瑚兰花芽由外到内,依次是花被片、唇瓣、合蕊柱;其中花被片包括萼片和花瓣,蕊柱顶端着生1
个花药,1个花药里有2个花粉块,观察显微图(图2-5)可知,花粉块密度明显大于周围组织,花粉块下部有蕊喙、花柱、子房。花被片,包覆在花芽的最外侧,外轮花被片日后发育成为3片萼片,内轮花被片则发育成为2片花瓣。
唇瓣位于合蕊柱之上,花被片之下,呈芭蕉叶状扣在花药顶部。附着点在合蕊柱侧面,由2~3层细胞构成,细胞排列紧密均匀。
合蕊柱位于花芽正中,周围由花被片包覆,顶端是有2个花粉块的花药,往下依次是蕊喙、花柱、子房。其中,花粉块的细胞密度明显大于周围组织,十分容易分辨。其次花粉块下方有1个略向上伸的突起,此处为蕊喙,别称小喙,是兰科植物的蕊柱前面舌状突起的部分。在柱头的3个裂片中,2个形成柱头,不结实性的1个即变成这种蕊喙(图2-5)。
子房在整个花结构处于靠下部分,中空,子房内壁上附着很多圆卵形小体,称为胚珠,受精后发育成种子,胚珠附着在子房中的胎座上。
胚珠是一种受精后发育成种子的结构,被称为种子的前体。子房内,胚珠包被,通过珠柄着生于子房内壁的胎座上(图2-6、图2-7)。
3 结论与讨论
吕梁山野生珊瑚兰资源分布于111°13′10″~111°13′15″E,37°5′31″~37°5′37″N;4月上旬开始萌动,4月下旬至5月下旬为开花期,6月中下旬种子成熟;中性土壤和9~20 cm土壤腐殖质能够为珊瑚兰植株生长提供一个稳定的生长环境;海拔高度、土壤平均腐殖质明显影响野生珊瑚兰居群分布。海拔为1 820~1 940 m的区域,野生珊瑚兰主要的伴生植物乔木层为10~30 m的大中型乔木,主要是糙皮桦、蒙古栎;1.0~3.0 m的灌木,主要是野山楂、蔷薇;草本层植物主要半生植物为舞鹤草、鲜有玉竹、鹿药、华北耧斗菜。
通过观察珊瑚兰的花芽发现,珊瑚兰的花芽形态特征与普通兰花保持一致,即由3片外轮花被片、2片内轮花被片、1个唇瓣、1个合蕊柱构成,在合蕊柱顶端有花药、蕊喙,底端為子房,特立中央胎座着生胚珠。常规实体解剖石蜡切片法观察珊瑚兰花芽分化依次为花序原基分化期、花被原基分化期、雄蕊原基分化期、雌蕊原基分化期、胚珠形成期。
关于吕梁山濒危植物珊瑚兰资源调查的工作由2016年4月开始,历经3年,在前期充分查阅资料制定了交城县庞泉沟、中阳县上顶山和离石区吴城九凤山等5个有代表性的,具备兰科植物的生境特点的样地,但是在踏查的过程中文献记录庞泉沟自然保护区八水沟、八道沟、福禄沟有野生珊瑚兰居群分布,但在调查过程中,通过3次调查均未发现有珊瑚兰居群分布,分析原因可能因为庞泉沟自然保护区旅游和矿业开发破坏了珊瑚兰生境,具体原因还需要进一步调查才能确定。中阳县上顶山属吕梁山关帝山脉,自然生态保护较好,但是在发现野生珊瑚兰居群分布后,笔者所在课题组又扩大范围对该地区野生珊瑚兰资源进行了调查,但是遗憾的是除了本研究范围,其他地方均未发现珊瑚兰。珊瑚兰自身无法进行光合作用,只能由其共生的菌类供给营养。此前有学者研究,珊瑚兰的共生真菌有较强的专一性,要满足珊瑚兰及其共生菌能够同时良好地生长,对环境的要求必然是较为严格的。目前关于珊瑚兰的研究很少,野生珊瑚兰外形奇特,花期较长,有很大的开发价值,现今野生种群数量呈明显下降趋势。由于种群数量较少,繁殖生境要求严格,直接导致种群扩散缓慢,保护和恢复该植物种群的结构和数量显得格外重要[18-20]。
参考文献:
[1]侯天文. 四川黄龙沟优势兰科植物菌根真菌多样性研究[D]. 北京:北京林业大学,2010.
[2]蒋玉玲. 辽宁省内九种兰科植物菌根真菌多样性研究[D]. 沈阳:沈阳农业大学,2018.
[3]沐先运,刘 晓,林秦文,等. 北京百花山自然保护区兰科植物多样性及保护评价[J]. 北京林业大学学报,2010,32(增刊1):14-17.
[4]石昌魁. 甘肅省兰科植物系统分类与区系地理[D]. 兰州:甘肃农业大学,2008.
[5]侯天文. 四川黄龙沟优势兰科植物菌根真菌多样性研究[D]. 北京:北京林业大学,2010.
[6]冉景丞,鲁成巍. 茂兰自然保护区兰科植物资源现状及保护利用途径[J]. 安徽农业科学,2009,37(11):5209-5211.
[7]孙 悦,李 标,郭顺星. 腐生型兰科植物研究进展[J]. 广西植物,2017,37(2):191-203.
[8]董建新,朱毓永,李铁军,等. 小五台山野生兰科植物研究[J]. 承德民族师专学报,2009,29(2):54-55.
[9]孙晓颖. 五种野生兜兰植物菌根真菌多样性研究[D]. 北京:北京林业大学,2014.
[10]孙晓颖,张武凡,刘红霞. 带叶兜兰种子原地共生萌发及有效菌根真菌的分离与鉴定[J]. 热带亚热带植物学报,2015,23(1):59-64.
[11]McKendrick S L,Leake J R,Taylor D L,et al. Symbiotic germination and development of myco-heterotrophic plants in nature:ontogeny of Corallorhiza trifida and characterization of its mycorrhizal fungi[J]. The New Phytologist,2000,145(3):523-537.
[12]刘桂芝. 石蜡切片标本的制作过程[J]. 通化师范学院学报,2004(11):90-91.
[13]李淑娴. 墨兰成花机理及花期调控技术研究[D]. 福州:福建农林大学,2016.
[14]王长宝,徐增奇,王 鹤,等. 濒危植物珊瑚兰(Cypripedium macranthos SW.)地下器官的形态解剖学研究[J]. 东北师大学报(自然科学版),2015,47(3):111-115.
[15]龚 湉. 寒兰成花机理及花期调控研究[D]. 福州:福建农林大学,2015.
[16]韦 莉. 蝴蝶兰花芽分化的解剖学观察及成花机理研究[D]. 南京:南京林业大学,2010.
[17]彭 芳. 文心兰花芽形态分化及其生理生化的研究[D]. 南宁:广西大学,2011.
[18]董林水. 晋西基岩山地与黄土丘陵区生态过渡带植被特征及生态恢复机理研究[D]. 北京:中国林业科学研究院,2005.
[19]李忠超,王武源. 濒危药用植物八角莲生态生物学特征[J]. 热带亚热带植物学报,2006,14(3):190-195.
[20]张 毓,张启翔,赵世伟,等. 濒危植物大花杓兰胚与珠被发育的研究[J]. 园艺学报,2010,37(1):72-76.
