脑脊液循环动力学机制研究进展
2021-09-02刘俊张国斌
刘俊 张国斌
经典脑脊液循环理论认为,脑脊液由脉络丛产生,侧脑室及三脑室中的脑脊液经正中孔及外侧孔到达小脑延髓池,其与脊髓腔脑脊液混合流入颅底各脑池,最终流向大脑凸面的蛛网膜下腔,由蛛网膜颗粒吸收入静脉窦完成循环[1]。然而,随着胶质淋巴循环理论的提出,脑脊液的产生来源、吸收途径、与脑组织间液的交换以及脑膜淋巴管等新发现,经典脑脊液循环理论受到了挑战。本文就中枢神经系统脑脊液循环动力学机制的争议和最新研究进展作一综述。
1 脑脊液来源与产生方式
1.1 脉络丛脑脊液由脉络丛产生是经典脑脊液循环理论核心之一。最早由CUSHING[2]和WEED[3]先后提出,之后在DANDY切除犬的脉络丛并堵塞室间孔的实验中得到证实[4]。WELCH[5]发现脉络膜静脉的血细胞压积要显著高于脉络膜前动脉,推测脉络膜血液生成了脑脊液。SPECTOR等[6]详细阐明脉络丛如何产生脑脊液的机制,即血浆在静水压作用下,由脉络丛内的毛细血管内皮细胞渗出到内皮间隙,再经过脉络丛上皮细胞上遍布的钠钾泵和水通道蛋白1(aquaporin 1,AQP1)释放到脑室中形成脑脊液。最新研究发现脉络丛可在脑代谢废物增加时或睡眠周期中的特殊时间点代偿性增加脑脊液的生成,以优化脑代谢废物的清除效率[7]。
然而,许多研究并不支持脉络丛为脑脊液唯一产生部位,例如BERING同样在犬身上实施DANDY的实验操作,却无法观察到切除脉络丛侧的脑室发生塌陷[8]。MILHORAT发现切除恒河猴的脉络丛后,其侧脑室内脑脊液生成量较未切除脉络丛的仅轻微减少。婴幼儿脑积水患者在接受脉络丛切除术后,仍有三分之二复发而不得不接受分流手术[9]。之后不断有研究质疑脑脊液只来源于脉络丛的这一经典理论,人们越发相信脑脊液产生可能存在其他来源。
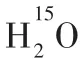
1.3 脑组织间液除了脉络丛及蛛网膜下腔毛细血管超滤液,脑组织间液也可能是脑脊液来源之一。早在2001年,PÉREZ-FÍGARES 等[16]即推测,20%脑脊液来源于脑组织间液。RENNELS等[17]发现脑脊液和脑间质液之间交换的最初证据,在犬和猫蛛网膜下腔注入标记物,发现其可沿着血管旁途径在10 min内进入大脑实质。然而,该研究是基于麻醉这种非生理状态。
直到2012年,由NEDERGAARD团队通过双光子激光扫描显微镜首次在活体鼠中发现蛛网膜下腔脑脊液和组织间液之间存在交换,并且是通过AQP4介导,这一机制被称之为胶质淋巴循环(glympic pathway)[18]。该理论认为,在血管周围间隙中,脑脊液和组织间液是对流的,组织间液可以通过星形细胞顶端的AQP4快速运输,在足突端被转运至血管周围间隙与脑脊液发生交换,进而随着脑脊液到达大脑表面被蛛网膜颗粒和静脉窦吸收,这一机制有力地证明脑组织间液也是脑脊液的重要来源[19-21]。近年来,人们利用核磁示踪及荧光显像技术已在活体动物或人体中发现了脑组织间液与脑脊液交换的证据。HAN等[22]利用核磁显像发现活体大鼠尾状核处的组织间液可逐渐流向大脑皮层背侧和腹侧并与其蛛网膜下腔脑脊液汇合。RINGSTAD等[23]在人体腰大池鞘内注射示踪剂并行强化核磁显像,发现示踪剂以一种向心的方式首先在大脑前、中、后动脉的前纵裂区、侧裂区、Wills环区富集,且6~9 h后达到高峰,然后示踪剂逐渐向基底节区、丘脑、中脑等脑深部区域转移,24 h后基本消失。最近SWEENEY等[20]利用荧光成像技术在活体动物上发现了脑脊液与脑组织间液交换的证据,另外,TAOKA及MESTRE等[24-25]在脑外伤、脑小血管病、梗塞性脑水肿及阿尔兹海默病等中枢神经系统疾病中也发现了脑脊液和脑组织间液交换的证据,只不过这种交换效率会显著受损。由此可见,胶质淋巴循环理论大大丰富了传统的脑脊液循环认知,但脑脊液和脑组织间液如何深度地交换融合从而参与脑的代谢和稳态仍旧需要进一步探索。
2 脑脊液循环的流体动力学模式
2.1 从单向流动到双向流动经典的脑脊液循环模式指出脑脊液循环是沿着脑室-蛛网膜下腔-静脉窦静水压力梯度的单向流动。但是,OREŠKOVIC等[26]认为如果脑脊液循环是单向循环,那么处于平卧位时,由于脉络丛和静脉窦的压力梯度消失,脑脊液循环可能就停滞了,这显然是矛盾的。另外,一些临床研究也质疑了脉络丛是脑脊液生物泵的理论。TAKIZAWA等[27]利用四维相位对比和时空标记反转脉冲技术对11名健康志愿者侧脑室三角区脉络丛附近的脑脊液活动进行核磁扫描,发现此区域脑脊液流速显著低于脑干腹侧、三脑室和四脑室脑脊液活动,既然脉络丛推动着脑脊液向前流动,那么此处的脑脊液流速不应该是受到抑制的,这同样与脉络丛是脑脊液生物泵的理论相悖。
BULAT-OREŠKOVIC-KLARICA理论认为,不存在脉络丛的生物泵作用,脑脊液循环是双向的,即随着心脏的收缩舒张来回位移,可以从脑室到蛛网膜下腔,亦可以相反从蛛网膜下腔到脑室,脑脊液和脑组织液中任何物质的扩布距离取决于物质的半衰期,半衰期越长,最终将分布到更远的距离。早在1992年NILSSON等[28]即在核磁研究中观察到脑脊液循环是双向的,即导水管的脑脊液发生搏动性流动,流速随心动周期而变化,在心脏收缩期,脑脊液以大约8 mm/s速度流向后脑(尾端),然后在心脏舒张期间以5 mm/s速度缓慢地回到前脑(头颅)。因此脑脊液的净方向是朝向后脑。国内学者[29]同样验证了NILSSON发现,他们利用相位核磁技术观察了净中脑导水管每搏量,正常人多为头尾向,而常压性脑积水患者收缩期和舒张期则均为尾头向,且呈高动力状态。该理论可以较好的解释体位的变化脑脊液循环不会受到明显的影响,但是双向循环终归是一种理想的流动模式。因为双向循环理论不能很好解释脑实质内脑脊液与脑组织液的交换,也无法解释脑代谢废物如何排出。
2.2 脑脊液和脑组织间液通过胶质淋巴循环深度融合众所周知,流体的运动既有分子扩散,也有整体流,脑脊液亦是如此。胶质淋巴循环理论认为,蛛网膜下腔的脑脊液与脑组织间液在毛细血管的周围间隙即Virchow Robin空间(VRS)实现深度交换融合,但基于这种交换的流体动力学诸如驱动力和对流方向等研究目前仍存在争议。EIDE等[30]在常压脑积水患者重复了先前RINGSTAD的核磁实验,他发现脑积水患者较正常人向心性的富集效应显著减弱,认为这种有规律的富集很难用分子扩散解释,脑脊液循环可能是在胶质淋巴循环的作用下,以一种整体流的方式运行。
由于分辨率的限制,目前很难利用常规核磁成像观察VRS中脑脊液与脑组织液交换形式。最近DAVERSINCATTY等[31]利用计算机模拟成像技术发现VRS中脑脊液与脑组织液交换是一种振荡流和对流的综合,由脑动脉壁的扩张、刚性运动以及静态脑脊液压力梯度共同控制。同样KOUNDAL等[32]利用动态对比增强核磁显像技术及数学模型在大鼠中发现,溶质在颅底的大血管旁循环比在中脑和海马迅速的多,推测蛛网膜下腔和脑池中的脑脊液循环以对流占主导,而在脑实质中,组织液与脑脊液的交换以分子扩散为主,这就能够解释脑积水时由于脑室压升高会导致室管膜通透性增高,脑脊液到组织液的流量增加导致脑白质间质水肿,但是由于蛛网膜下腔脑脊液和组织液的引流效率更高,脑积水却不会导致大脑皮层灰质水肿。FAGHIH等[33]发现脑脊液-脑组织液的循环不仅通过简单的分子扩散,而且通过压力梯度和旋转重复加速和减速导致非常复杂的运动。由此可见,脑脊液运动不再认为是简单的循环流动,而是脑室和蛛网膜下腔中各种方向的流动组合,从而适应日常生命活动等复杂的生理过程。
3 脑脊液-组织间液的吸收
3.1 脑膜/脊膜淋巴管脑脊液的产生和吸收是动态平衡的,经典脑脊液循环模式中,脑脊液由硬脑膜上蛛网膜颗粒吸收入静脉窦。近些年随着脑膜淋巴管的发现,证实了相当比例的脑脊液是通过脑膜淋巴管引流吸收的。
早在18世纪,PAOLO MASCAGNI首先提到了脑膜淋巴管的存在。由于过于超前,该脑膜淋巴管的存在一直受到质疑。20世纪60年代,FÖLDI等[34]重新描述了中枢神经系统与周围神经系统之间存在淋巴连接。LI等[35]首次在电镜下观察到硬脑膜上存在类圆形孔样的淋巴管,称为“硬脑膜淋巴孔”,推测该孔可能为蛛网膜下腔脑脊液与颅外结缔组织及淋巴系统之间沟通的潜在直截通路。
脑膜淋巴管主要沿着硬脑膜静脉窦分布。ABSINTA等[36]对人类和非灵长类动物进行硬脑膜淋巴管核磁强化成像,利用两种不同的增强剂对比呈现了脑膜淋巴管的地形图,证实了硬脑膜淋巴管的存在,且发现其主要分布在矢状窦旁附近。NAGANAWA等[37]利用特殊三维实时反转恢复核磁成像评估内耳淋巴积水时,意外发现在大脑皮质静脉壁与软脑膜鞘之间间隙同样强化,且似乎沿着上矢状窦与硬脑膜淋巴管相连,推测这一间隙可能是脑脊液吸收入硬脑膜淋巴管的途径。RINGSTAD等[38]研究同样支持这种观点,18例不同脑脊液循环紊乱疾病患者进行鞘内注射脑脊液示踪剂后行核磁强化,发现对比剂集中在皮质静脉汇入矢状窦的窦旁硬脑膜附近,他认为的确存在类似于跨蛛网膜的分子通道,其位于矢状窦旁的硬脑膜,可能是沟通脑脊液和硬脑膜淋巴管的桥梁。
脑膜淋巴管可通过嗅神经淋巴途径参与脑脊液吸收。BRADBURY等[39]发现蛛网膜下腔内脑脊液可以穿过筛板沿着嗅神经进入鼻粘膜的淋巴管,最后汇入颈部淋巴结。ASPELUND等[40]和 LOUVEAU等[41]在啮齿类、鱼类、灵长类以及人类发现,脑脊液成分的分子片段会引流到脑膜淋巴管并沿着肩胛后静脉和乙状窦以及翼腭动脉的脑膜部分汇入到颈深淋巴结。如前所述,RINGSTAD等[38]研究同样发现18例患者有2例筛板附近出现了对比剂增强,推测筛板附近的嗅神经途径参与了脑脊液吸收。
除了硬脑膜及嗅神经,脊神经附近蛛网膜颗粒和硬脊膜淋巴管也可参与脑脊液的吸收。KIDO等[42]及MIURA等[43]发现了脊神经根处存在蛛网膜颗粒和淋巴管。BOZANOVICSOSIC等[44]发现猫有16%的脑脊液通过脊髓周围流出道排出。KOH等[45]发现脊髓蛛网膜下腔的脑脊液可以沿着脊神经汇入肋间淋巴结。
3.2 蛛网膜颗粒尽管蛛网膜颗粒是脑脊液吸收的重要部位,但最新研究表明,蛛网膜颗粒及静脉窦可能并非恒定参与脑脊液循环。AKAI等[46]在活体胎鼠上发现,不论在脑室还是在蛛网膜下腔注射荧光剂,均未发现矢状窦和蛛网膜颗粒发现荧光信号。EIDE在核磁上观察到示踪剂是随着脑脊液向心性的转移直至消失[30],示踪剂并不在硬脑膜及静脉窦旁富集,这进一步质疑了蛛网膜颗粒的吸收功能。ZAKHAROV等[47]研究与这种推论相吻合,他在绵羊小脑延髓池注射放射性示踪剂后发现,当颅内压正常时,脑脊液流出的主要途径是淋巴管,而只有颅内压升高时,才会触发向蛛网膜颗粒及颅内静脉系统的转运。MURTHA等[48]也在大鼠上发现,脑室注射的造影剂并没有出现在静脉窦中,而是大部分很快穿过筛板进入嗅神经周围鼻粘膜。
除了蛛网膜颗粒和静脉窦,MALOVESKA等[49]发现脑脊液的引流吸收途径可能部分是通过胶质淋巴循环进入脑膜淋巴管或者穿过筛板到嗅神经周围淋巴管或者直接被脑膜淋巴管吸收完成。当然,血管旁路的胶质淋巴循环和脑膜淋巴循环是否存在协同作用目前仍不清楚。关于在不同的生理和病理条件下脑脊液吸收如何有所侧重地发挥作用,目前仍缺少研究。另外,MA等[50]报道,寰枕筋膜处枕下小直肌与枕部硬脑膜之间的肌硬膜桥可能参与调节局部脑脊液循环,当刺激犬枕下肌肉收缩或者被动转头时局部脑脊液压力增加。最近发现对美洲短吻鳄枕下肌硬膜桥实施电刺激收缩也出现了局部脑脊液流速增加的现象[51]。
因此,脑脊液循环动力学异常复杂,其来源、循环模式以及吸收部位可能并非一成不变,而是由众多因素调控。不同的日常生理性活动及病理性疾病状态下脑脊液循环动力学可能会发生不同变化,后期仍需要深入研究。
