低温胁迫对诺丽幼苗叶片光合荧光特性的影响
2021-08-06杨朴丽徐荣杨焱李海泉
杨朴丽 徐荣 杨焱 李海泉

摘 要:为了解诺丽抗寒生理特性,比较其抗寒能力,以4个诺丽(Morinda citrifolia L.)种质幼苗为试材,研究不同低温(25、12、9、6、3、0 ℃)及不同时间(1、3、5 d)胁迫对各种质幼苗叶片光合及叶绿素荧光参数等指标的影响,利用隶属函数法对4个诺丽种质耐寒能力进行评价。结果表明:(1)相同时间下随着胁迫温度的降低,4个诺丽种质幼苗叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和电子传递速率(ETR)显著下降,而胞间二氧化碳浓度(Ci)、PSⅡ潜在活性(Fv/Fo)和PSⅡ最大光化学效率(Fv/Fm)则呈降-升-降趋势,不同处理间差异显著;(2)相同温度不同时间处理,除Ci外,各种质其余指标皆随胁迫时间的延长显著下降,6 ℃以下低温处理Ci含量随胁迫时间延长显著增加;(3)温度和时间产生交互作用,相同时间显示低温危害强度效应,相同低温显示低温危害累积效应,影响着各诺丽种质幼苗叶片的光合及荧光指标;(4)隶属函数综合评价结果表明,4个诺丽种质抗寒能力依次为‘NL004>‘NL003>‘NL001>‘NL002。综合分析表明,参試的4个诺丽种质中,‘NL004的抗寒适应性最强,‘NL002最弱,研究结果为诺丽抗寒栽培和育种提供参考。
关键词:诺丽;低温胁迫;光合作用;叶绿素荧光;抗寒能力评价
Abstract: The purpose of this study was to understand the physiological characteristics of cold resistance of Morinda citrifolia L., and compare the cold resistance ability of different germplasms. Four seedlings of noni germplasms were used as the test materials to study the effects of low temperature (25, 12, 9, 6, 3, 0 ℃) and (1, 3, 5 d) stress on the leaf photosynthesis and chlorophyll fluorescence parameters, and membership function method was used to evaluate the cold tolerance of the four seedlings of noni germplasms. With the decrease of stress temperature at the same time, the net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr) and electron transfer rate (ETR) of the leaves of 4 noni germplasms seedlings decreased significantly, while the intercellular carbon dioxide concentration (Ci), PSⅡ potential activity (Fv/Fo) and PSⅡ maximum photochemical efficiency (Fv/Fm) showed a downward-upward-downward trend, with significant differences among different treatments. With the treatments at the same temperature but different time, except Ci, other indexes of all germplasms decreased significantly with the extension of stress time. while Ci content increased significantly under 6 ℃ with the extension of stress time. The interaction between temperature and time showed the intensity effect of low temperature damage at the same time, and the accumulation effect of low temperature damage at the same low temperature, which affecting the photosynthesis and fluorescence indexes of seedling leaves of various noni germplasms. The comprehensive evaluation results of membership function showed that the cold resistance of 4 noni germplasms was ‘NL004 > ‘NL003 > ‘NL001 > ‘NL002. Comprehensive analysis showed that among the 4 tested noni germplasms, ‘NL004 had the strongest cold resistance adaptability and ‘NL002 was the weakest. The research results would provide references for cold-resistant cultivation and breeding of noni.
Keywords: Morinda citrifolia L.; low temperature stress; photosynthesis; chlorophyll fluorescence; cold tolerance evaluation
诺丽(Morinda citrifolia L.)属茜草科(Rubiaceae)巴戟天属(Morinda)植物,别名海巴戟、印桑葚、诺尼,是热带多年生常绿灌木或小乔木,其果实是典型的低钠、高钾食物,果汁可作为天然高能、无脂、低糖保健食品[1]。诺丽被波利尼西亚人作为民间药物已有2000多年的历史[2],Heinicke[3]在诺丽中发现丰富的塞洛宁原,它会在人体的肠道内转化成赛诺宁,当身体处于亚健康状态或病态时,身体对赛洛宁的需求量会大大增加。众多研究表明长期食用诺丽可促进消化、缓解疼痛、抗菌消炎,增强免疫力[4-5]。随着人们生活水平的提高,人群中肥胖症、高血压、糖尿病、癌症等疾病发生率越来越高,服用诺丽是人体补充赛洛宁、改善身体素质、预防疾病的最佳选择。因此,诺丽市场开发前景广阔,然其受环境制约严重,主要分布在海南岛和云南西双版纳等地,冬季低温严重影响诺丽生产及产业化发展。选育抗寒品种和提高栽培技术是亟待解决的难题。
目前国内对诺丽抗寒性的研究报道较少,尽管赵帅等[6]通过室内模拟低温胁迫对5个诺丽种质进行相关生理指标的测定评价了5个种质的抗寒性,结果表明相对电导率、丙二醛含量和过氧化物酶活性可作为诺丽抗寒性评价的参考指标,5个诺丽种质中‘WT1a种质的抗寒能力最强,‘WT4的抗寒能力较差。目前尚未见低温胁迫下诺丽光合荧光动力学特性的研究报道。采用光合作用系统结合叶绿素荧光技术可快速、灵敏、无损伤地探测低温对植株的胁迫效应,目前已应用于油棕[7]、果蔗[8]、南美胡荽[9]、降香黄檀[10]等多种热带、亚热带植物的低温胁迫鉴定。诸多研究表明,光合荧光参数可作为筛选耐寒品种的重要指标,低温逆境会导致净光合速率、PSⅡ最大光化学效率显著下降,抗寒性弱的品种降幅显著大于抗寒性强的品种[10-12]。本文以4个诺丽种质为试材,分析了不同时间、不同低温胁迫处理诺丽幼苗叶片净光合速率、气孔导度、胞间二氧化碳浓度、蒸腾速率及叶绿素荧光参数等指标的变化,探究诺丽光合荧光特性对低温胁迫的响应,并采用隶属函数法分析4个种质的耐寒能力,以期为扩大引种、耐寒种质筛选和抗寒性栽培提供理论依据。
1 材料与方法
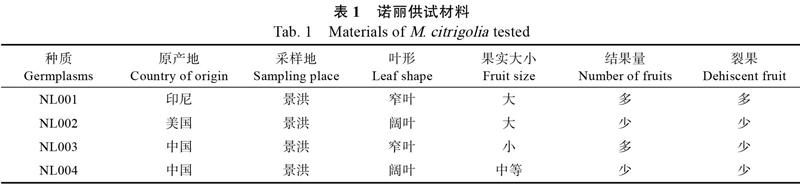
1.1 材料
試材所用的4个诺丽种质(表1)均引种栽培在西双版纳州云南省热带作物科学研究所诺丽种质保存基地,其中‘NL003和‘NL004均来自中国海南省。4个种质间叶形、果实等相关性状区别明显。2018年2月将成熟种子播种于沙床上,待长出2对真叶后移栽至14 cm×14 cm花盆中,温室大棚内培养,棚内年平均气温为28~ 30 ℃,湿度为60%~65%,待苗长至4~5对叶时进行低温胁迫处理。
1.2 方法
1.2.1 试验设计 选取高度、生长一致的健壮植株,移入不同温度(0、3、6、9、12 ℃)的人工气候箱,以常温条件(25 ℃)下的植株为对照,进行不同时间(1、3、5 d)处理,开展多因素试验,培养箱设置每天光照11 h,光照强度为6000 lx,相对湿度为80%;每处理5株苗,3次重复。胁迫处理后,选取诺丽心叶下第2对功能叶,进行光合及叶绿素荧光参数的测定。
1.2.2 测定指标及方法 使用美国LI-COR公司便携式光合仪LI-6400XT测定植物的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),测定光强为1200 μmol/(m2·s),CO2浓度为400 μmol/mol,叶室温度为25 ℃,流速为500 μmol/s。选取诺丽心叶下第2对功能叶,并做标记,之后每次测定使用相同叶片,每枚叶片测定3次,每个处理3株重复。
叶绿素荧光参数采用德国WALZ-便携式调制叶绿素荧光仪PAM-2500进行测定,测定前对待测叶片暗适应20 min,避开叶脉,先测定叶片初始荧光(Fo),最大荧光(Fm),通过以上参数计算PSⅡ潜在活性(Fv/Fo),PSⅡ最大光化学效率Fv/Fm,电子传递速率ETR。
1.2.3 抗寒性综合评价 隶属函数法已广泛应用于果蔗[8]、降香黄檀[10]、油茶[13]、橡胶树[14]等植株的抗寒性鉴定,隶属函数平均值越大,抗寒性越强[15]。采用隶属函数法对所测得的各项指标进行转换,计算隶属函数值和平均隶属度,综合评价分析不同诺丽种质的抗寒性。如果分析指标与抗寒性呈正相关,用公式(1)计算;若呈负相关,则采用公式(2)计算:
1.3 数据处理
利用SPSS 23.0软件对试验数据进行统计分析。以一般线性模型的单变量的全因子分析同一种质不同温度、不同时间处理对光合及荧光指标的影响,采用ANOVA过程作方差分析,采用LSD法、Duncans法进行多重比较分析,采用CORR过程作线性相关性分析。采用隶属函数法对4个诺丽种质的耐寒性进行综合评价。
2 结果与分析
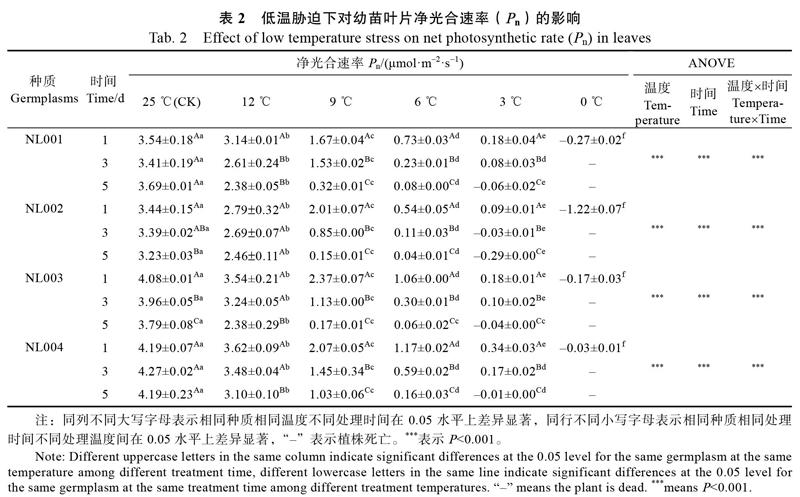
2.1 低温胁迫对诺丽幼苗净光合速率(Pn)的影响
不同低温胁迫对诺丽净光合速率(Pn)的影响见表2。随着处理温度的降低,不同种质的净光合速率均呈显著下降趋势,且温度越低则降幅越大;随低温胁迫天数的增加,不同种质的净光合速率显著下降,胁迫时间越长Pn值越小。12 ℃处理下,各种质相对于对照,Pn缓慢下降;9 ℃低温下Pn显著低于对照和12 ℃,9 ℃处理5 d的Pn大幅度下降,极显著低于处理1 d和3 d;6 ℃的Pn显著低于9 ℃,且随胁迫时间的延长Pn值越来越小;3 ℃处理1 d的Pn与6 ℃处理1 d相比下降了77.21%,随胁迫时间延长同一种质各处理间差异显著降低;低温3 ℃处理3 d时,‘NL002的净光合速率达到负值,3 ℃处理5 d与0 ℃胁迫1 d时,各种质的净光合速率均达到负值,随0 ℃胁迫时间延长植株死亡。相同温度相同处理时间下,4个诺丽幼苗叶片的Pn值由大到小依次为:‘NL004>‘NL003>‘NL001>‘NL002,说明‘NL004相对其他3个种质更耐低温。通过单变量全因子分析得出,各种质的主体间效应全部极显著,说明时间、温度对Pn都会造成极显著影响(P<0.001),相同时间显示低温危害强度效应,相同低温显示低温危害累积效应。
2.2 低温胁迫对诺丽幼苗气孔导度(Gs)的影响
如表3所示,同一种质在相同时间下,不同温度处理间存在显著差异,总体上温度下降气孔导度值变小;相同温度下不同时间各处理也差异明显,处理时间越长Gs值越小。胁迫温度从12 ℃降至9 ℃时,诺丽参试种质的Gs值大幅度下降,低于9 ℃的低温处理叶片的Gs值虽呈下降趋势,但降幅较小,0 ℃处理3 d植株死亡。相同温度处理下随胁迫时间的延长,各种质Gs值显著下降,与胁迫1 d、3 d处理相比,胁迫5 d各处理的Gs值下降到最低水平,不同天数间差异显著。从单变量全因子分析的结果来看,温度、时间对诺丽幼苗叶片气孔导度的变化均有影响,而且温度与处理天数有交互作用(P<0.001)。温度和时间产生的低温危害效应与Pn相似。
2.3 低温胁迫对诺丽幼苗胞间CO2浓度(Ci)的影响
從表4可以看出,相同胁迫时间不同低温下各种质的Ci浓度差异显著,相同处理温度不同胁迫天数之间也存在显著差异。相同时间处理下,各种质诺丽幼苗叶片Ci浓度随处理温度的降低呈先降后升的变化趋势。总体上12 ℃和9 ℃的Ci浓度低于对照,而6 ℃和3 ℃条件下却比对照高,3 ℃下各种质的Ci浓度较高,0 ℃胁迫1 d时达最大值,随0 ℃胁迫时间延长寒害严重致植株死亡。‘NL001‘NL003和‘NL004在9~0 ℃低温处理下,胁迫1、3和5 d处理均随温度降低,诺丽叶片Ci浓度呈上升趋势,而‘NL002在12 ℃时,胁迫3 d和5 d处理则随温度降低Ci浓度呈上升趋势,说明‘NL002相较其他3个种质而言对低温更敏感。从单变量全因子方差分析的结果可知,温度、时间对诺丽幼苗叶片气孔导度的变化均有影响,且温度与处理天数有交互作用。处理天数(时间)除对‘NL001幼苗气孔导度呈显著影响(P<0.05)外,对其余种质的Ci浓度均呈极显著影响(P<0.001)。说明‘NL001对相同温度显示的低温危害效应略低于另外3个种质。
2.4 低温胁迫对诺丽幼苗叶片蒸腾速率(Tr)的影响
不同低温不同处理时间对4个诺丽种质幼苗叶片蒸腾速率的影响见表5。相同处理时间,各种质叶片的Tr值随胁迫温度的下降呈下降趋势,各处理间差异显著。相同低温条件下,随低温处理时间的延长,4个诺丽种质幼苗叶片的Tr值均显著下降。9 ℃下,‘NL004的Tr值随处理时间延长降幅显著低于其余3个种质,而‘NL002的Tr值下降速度最快。6 ℃低温下相对其他种质,‘NL002的Tr值最低,‘NL004最高。在3 ℃低温胁迫1 d时,‘NL004的Tr值有一个跃升,显著高于其他3个种质。上述可得出,‘NL004抗寒性较强,‘NL002抗寒相对较弱。9~3 ℃条件下,胁迫3 d和5 d的处理Tr值均出现缓慢下降,随胁迫天数的增加,4个种质Tr值的变化幅度相对较小。由表5还可看出,时间、温度主效应极显著,“温度时间”互作对Tr值具有极显著影响(P<0.01)。时间和温度对各种质产生的危害效应同于上述Pn和Gs。
2.5 低温处理对诺丽PSⅡ潜在活性(Fv/Fo)的影响
由表6可知,相同温度下4个种质均随着胁迫时间的延长Fv/Fo值逐渐降低,处理间差异显著,表明长时间低温胁迫导致诺丽幼苗抗寒性降低。相同时间下,各种质对照的Fv/Fo值显著高于各低温处理,随温度的降低变化程度各不相同,‘NL001在3 ℃处理下出现跃升,Fv/Fo值均显著高于相同时间的9 ℃和6 ℃处理,‘NL002和‘NL004在6 ℃时出现跃升,‘NL002只在6 ℃胁迫1 d时Fv/Fo值显著高于9 ℃和3 ℃处理,而‘NL004在6 ℃胁迫1 d和3 d下均显著高于9 ℃和3 ℃,综合整体数据可看出,‘NL002的Fv/Fo值显著低于其余3个种质,说明其受低温胁迫的影响大。从表6还可看出,温度、时间分别极显著影响诺丽幼苗叶片的PSⅡ潜在活性,温度与处理天数的交互作用对‘NL001种质的Fv/Fo值造成显著影响(P<0.05),其余种质温度与处理天数交互作用产生极显著影响(P<0.001)。说明相同时间显示低温危害强度效应,相同低温显示低温危害累积效应。
2.6 低温胁迫对诺丽PSⅡ最大光化学效率(Fv/Fm)的影响
表7为不同低温不同处理时间对诺丽叶绿素荧光参数Fv/Fm的影响。4个诺丽种质在低温胁迫下Fv/Fm的变化趋势与表6中Fv/Fo值的变化情况相似,‘NL001在3 ℃下Fv/Fm值出现跃升,显著高于相同时间下的9 ℃和6 ℃处理,6 ℃条件下,不同处理时间之间差异不显著。‘NL002在6 ℃胁迫1 d时Fv/Fm值出现跃升且显著高于9 ℃和3 ℃处理,胁迫5 d时,6 ℃与3 ℃处理的Fv/Fm值差异不显著。‘NL003在相同时间下,Fv/Fm值随温度降低显著下降,3 ℃的低温处理,胁迫3 d与5 d间差异不显著。‘NL004在胁迫1 d时,12~3 ℃的低温处理间Fv/Fm值差异不显著,且‘NL004的Fv/Fm值在各个低温胁迫下均高于其余种质,说明‘NL004的耐寒能力相对较强。从表7还可得出,温度、时间分别极显著影响诺丽幼苗叶片的PSⅡ最大光化学效率(Fv/Fm),且温度与处理天数有交互作用(P<0.001),低温危害效应的表现与Fv/Fm相同。
2.7 低温处理对诺丽叶片表观量子传递速率(ETR)的影响
低温胁迫对ETR的影响如表8。各供试材料在各低温胁迫下,ETR均下降,明显低于正常水平,且随胁迫时间的延长,下降幅度均增大,同一种质相同胁迫时间下,不同低温胁迫处理间差异均达显著水平(P<0.05),温度越低下降幅度越大。由表8还可知,0 ℃处理3 d和5 d属于极重度低温胁迫,最终导致植株经受不住寒冷死亡,‘NL002在3 ℃胁迫3 d和5 d后量子传递速率出现0值,表明‘NL002受低温寒害影响较大。温度、时间分别极显著影响诺丽幼苗叶片的ETR,除‘NL004外,温度与处理天数对其余种质的ETR有交互作用(P<0.001)。
2.8 诺丽不同种质光合及荧光指标的相关性分析
对光合指标和叶绿素荧光参数进行相关性分析表明(表9),Pn与Gs、Tr、Fv/Fo、Fv/Fm和ETR呈极显著正相关(P<0.01),与Ci呈极显著负相关(P<0.01);ETR与Ci呈极显著负相关,与其余指标皆呈极显著正相关;Ci与Gs、Tr无显著相关性,但与其余指标存在着极显著负相关关系。以上分析说明,Ci与大部分指标呈负相关,起到负面效应,其值越高抗寒性越低。Pn、Gs、Tr、Fv/Fo、Fv/Fm和ETR等对诺丽抗寒起正向作用,其值越高,诺丽越抗寒。说明低温胁迫对诺丽的影响是多方面的,在诺丽抗寒鉴定中,以上指标均可作为衡量诺丽光合能力的参数。
2.9 利用隶属函数综合评价诺丽的抗寒性
本研究在低温胁迫下测定4个诺丽种质光合及荧光参数的基础上,采用隶属函数对不同诺丽种质进行抗寒性综合评价。由表10可知,4个诺丽种质的隶属度在0.33~0.46之间,从隶属函数综合评价值可得出4个参试诺丽种质的抗寒性依次为‘NL004>‘NL003>‘NL001>‘NL002。‘NL004的综合评价值高达0.445,抗寒性最强,‘NL003和‘NL001的综合评价值分别为0.41和0.37,抗寒性次之,‘NL002的综合评价值仅为0.33,抗寒性最差。
3 讨论
Pn是光合系统功能的直接体现,低温对光合作用的影响大多数植物表现出Pn明显下降[16-17]。Pn下降的主要原因,除去环境因子和气孔导度的影响,非气孔限制因素也发生作用[18]。气孔是植物与大气进行水汽和CO2交换的通道,影响着植物的光合与蒸腾[19]。气孔限制因素引起Pn下降表现为Gs、Tr下降,Ci也相应下降,非气孔限制因素引起的Pn下降表現为Gs、Tr下降的同时Ci升高[20-22]。本研究结果表明,相同胁迫时间下,25~9 ℃的温度处理Pn下降主要受气孔限制因素引起光合作用受限制所致,而9 ℃以下低温处理Pn下降很大程度来源于非气孔限制因素。
Fv/Fo是PSⅡ潜在活性,ETR表示实际光强下的表观电子传递效率,反映植物叶片总体光合能力[23]。Fv/Fm表示PSⅡ的最大光化学效率,常用作环境胁迫程度的指标和探针,对于由非气孔因素所导致的Pn下降,Fv/Fm降低表明植物受到光抑制[24-25]。本研究结果表明,‘NL001在9~6 ℃下存在光合作用非气孔限制因素产生了光抑制,而‘NL002和‘NL003由非气孔限制因素产生的光抑制还包括3 ℃低温处理,其中‘NL002的Fv/Fm下降幅度较大,可见‘NL002受低温胁迫的影响较大。这与前人在其他植物上的研究结果相似[9-10, 26-27]。此外本文研究还得出温度和时间的交互作用也对各种质的光合及荧光参数指标有一定的影响作用。
抗寒性是植物在对低温寒冷环境长期适应过程中,通过本身的遗传变异和自然选择获得的一种抗寒能力,单一抗寒指标难于判断植物对寒冷的综合适应能力[28]。本研究中低温胁迫对诺丽幼苗叶片的光合机构造成一定的伤害,但只是通过几个光合相关的生理指标对诺丽的抗寒性进行了初步比较,后续还需进一步测定其他生理生化指标,结合具体表型的田间观察,才能全面评价其抗寒能力。
4 结论
随胁迫温度降低、时间延长,4个诺丽种质幼苗叶片Pn、Gs、Tr和ETR呈显著下降趋势,而Ci、Fv/Fo和Fv/Fm值呈降-升-降变化。相同时间下,9 ℃以上的温度处理净光合速率下降主要受气孔限制因素所致,而9 ℃以下低温处理Pn下降很大程度来源于非气孔限制因素。相同温度下,6 ℃和3 ℃处理,胁迫时间延长,Ci显著增加,不同胁迫时间处理Pn下降大部分由非气孔限制因素引起。相关性分析结果表明所测指标均可作为衡量低温环境下诺丽光合能力的参数,由隶属函数综合分析可知,参试的4个诺丽种质中‘NL004的抗寒适应性最强,其次‘NL003,再次‘NL001,‘NL002最弱。
参考文献
杨 焱, 杨朴丽, 徐 荣, 等. 不同诺丽种质外观性状与营养成分的分析及评价[J]. 热带作物学报, 2017, 38(1): 53-58.
Sanni D M, Fatoki T H, Kolawole A O, et al. Xeronine structure and function: Computational comparative mastery of its mystery[J]. Silico Pharmacology, 2017, 5(1): 8.
Heinicke R M. The pharmacologically active ingredient of noni[J]. Bulletin Pacific Tropical Botanical Garden, 1985, 15: 10-14.
张伟敏, 符文英, 施瑞城, 等. 诺丽果实和叶中主要功能性物质的分布与营养评价[J]. 食品科学, 2008, 29(10): 575-577.
龚 敏, 苏 奎, 邓世明. 诺丽鲜果与诺丽发酵汁的抗氧化及抑菌活性的对比研究[J]. 食品科技, 2010, 35(10): 92-94.
赵 帅, 阙艳鹏, 夏栋楠, 等. 5个海巴戟种质低温胁迫下的抗寒性评价[J]. 热带生物学报, 2018, 9(1): 69-75.
杨华庚, 林位夫. 低温胁迫对油棕幼苗光合作用及叶绿素荧光特性的影响[J]. 中国农学通报, 2009, 25(24): 506-509.
陈明辉, 程世平, 张志录, 等. 低温胁迫下不同果蔗品种光合及荧光特性的变化及耐寒性评价[J]. 热带作物学报, 2018, 39(3): 465-471.
杨柳青, 张 柳, 廖飞勇, 等. 低温胁迫对南美天胡荽光合作用和叶绿素荧光的影响[J]. 经济林研究, 2014, 32(4): 99-102.
张毅龙, 张卫强, 甘先华. 低温胁迫对6种珍贵树种苗木光合荧光特性的影响[J]. 生态环境学报, 2014, 23(5): 777-784.
周福平, 柳青山, 张一中, 等. 低溫胁迫对高粱幼苗叶绿素荧光参数的影响[J]. 种子, 2018, 37(9): 36-40.
刘 建, 项东云, 陈健波, 等. 低温胁迫对桉树光合和叶绿素荧光参数的影响[J]. 桉树科技, 2009, 26(1): 1-6.
石林曌, 倪天虹, 裴云霞, 等. 湖北省主栽油茶品种苗期抗寒性评价[J]. 经济林研究, 2019, 37(4) : 10-19.
张凤良, 李小琴, 杨 湉, 等. 橡胶树老幼态无性系对低温胁迫的生理响应[J]. 西部林业科学, 2019, 48(6): 127-132.
许桂芳, 张朝阳, 向佐湘. 利用隶属函数法对4种珍珠菜属植物的抗寒性综合评价[J]. 西北林学院学报, 2009, 24(3): 24-26.
周 建, 杨立峰, 郝峰鸽, 等. 低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J]. 西北植物学报, 2009, 29(1): 136-142.
崔 波, 程邵丽, 袁秀云, 等. 低温胁迫对白及光合作用及叶绿素荧光参数的影响[J]. 热带作物学报, 2019, 40(5): 891-897.
李法营, 高进波, 蓝增全, 等. 西双版纳引种的四个诺丽品系光合日变化研究[J]. 北方园艺, 2010(6): 18-22.
邵怡若, 许建新, 薛 立, 等. 低温胁迫时间对4种幼苗生理生化及光合特性的影响[J]. 生态学报, 2013, 33(14): 4237-4247.
Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 1982, 33: 317-346.
侯 伟, 孙爱花, 杨福孙, 等. 低温胁迫对西瓜幼苗光合作用与叶绿素荧光特性的影响[J]. 广东农业科学, 2014, 41 (13) : 35-39.
眭晓蕾, 毛胜利, 王立浩, 等. 低温对弱光影响甜椒光合作用的胁迫效应[J]. 核农学报, 2008, 22(6): 880-886.
Baker N R. Chlorophyll fluorescence: A probe of photosynthesis in vivo[J]. Annual Review of Plant Biology, 2008, 59(1): 89-113.
Constant S, Perewoska I, Alfonso M, et al. Expression of the psba gene during photoinhibition and recovery in Synechocystis PCC 6714: Inhibition and damage of transcriptional and translational machinery prevent the restoration of photosystem II activity[J]. Plant Molecular Biology, 1997, 34(1): 1-13.
张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报, 1999, 16 (4) : 444-448.
李清雪, 兰 岚, 贾志清, 等. 4种锦鸡儿属植物幼苗叶绿素荧光参数对重复低温胁迫的响应[J]. 林业科学, 2016, 52(10): 31-37.
辛洪河, 杨俊明, 曹书敏, 等. 早实核桃田间光合荧光特性的研究[J]. 经济林研究, 2011, 29(2): 72-77.
赵志平, 张阳梅, 原慧芳, 等. 西双版纳莲雾抗寒性比较试验[J]. 热带农业科技, 2013, 36(1): 29-33.
责任编辑:谢龙莲
