高寒草甸主要组分种开花物候对氮素添加的响应
2021-08-03刘旭东章志龙杜国祯
刘旭东,章志龙,杜国祯
(1.兰州大学生命科学学院 / 兰州大学草地农业生态系统国家重点实验室,甘肃兰州 730000;2.甘肃民族师范学院化学与生命科学系,甘肃合作 747000)
物候是生物受环境因素驱动的周期性现象[1],植物物候对陆地生态系统变化具有重要的指示作用,是响应区域环境变化最敏感的性状之一[2-3]。物候是研究植物响应气候环境变化最简单的方式[4],植物物候与环境变化(温度、水分和土壤养分)存在高度的关联性[5],是植物应对选择压力的适应性策略[6]。在全球变化日趋明显的背景下,物候变化不仅关系到植物个体的生存和繁衍,还将对群落生物多样性的维持产生重要影响[7]。
全球氮沉降的加剧被视为影响陆地生态系统结构和功能的重要驱动因子[8-9]。氮是高寒生态系统最重要的限制性养分元素[10],外源性氮输入则能够减轻氮营养对植物生长的限制[11]。高寒生态系统对环境变化的响应具有高度的敏感性[12-13],并对这些变化的响应具有超前性[14]。氮营养作为重要的环境因子明显地影响着群落结构和功能[15-17],也对草地群落植物物候有着不同程度的影响[18-20]。同时,物候作为气候变化的“指纹”,表现出生态系统对环境变化的响应和适应,在一定程度上决定着群落水平的进化过程[21-23]。尽管如此,目前关于环境变化影响群落花期物候的机理尚不清楚[24-25]。
对青藏高原草地群落物候的研究不仅有助于深入认识草地群落构建机制,也有助于了解气候变化(例如氮沉降)对其生态系统服务功能的影响,为预测和评估青藏高原高寒草地生态系统提供数据支撑。本研究基于高寒草甸6年氮添加试验平台,通过对不同氮添加处理下高寒草甸群落花期物候的观测,研究不同功能群物种花期物候对氮添加的响应和主要组分种花期物候参数之间的关系,以及氮添加条件下高寒草甸花期物候和群落格局的动态变化,以期明确氮添加对高寒草甸植物花期物候和群落动态的影响。
1 材料与方法
1.1 研究区域概况
研究地位于兰州大学高寒草甸与湿地生态系统定位研究站(玛曲县阿孜站,33°40′N,101°52′E),海拔约3 550 m,年均温1.2℃(在生长季6月–8月最高温度达到11.7℃,在非生长季11月−翌年4月温度低至−10℃),年均降水量为620 mm(图1)。年均日照时数约为2 580 h,年均霜降天数 >270 d;植被属于高寒草甸类,主要优势种有禾草类植物垂穗披碱草(Elymus nutans)、草地早熟禾(Poa pratensis)、胡氏剪股颖(Agrostis hugoniana),杂草类植物条叶银莲花(Anemone trullifolia)、钝裂银莲花(Anemone obtusiloba)、丝叶毛茛(Ranunculus tanguticus)、小花草玉梅(Anemone rivularis)和瑞苓草(Saussurea nigrescens)等。研究样地自2008年起围封,仅冬季放牧。
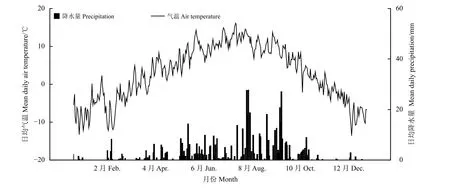
图1 2014年研究地日均气温与降水量变化Figure 1 Mean daily temperature and daily precipitation of thestudy site in 2014
1.2 试验设计
于2009年4月在围栏封育的季节性放牧草地中设置15个10 m ×6 m 的样区,各样区间距为2 m。在每个样区内,随机设置3个氮添加处理小区,分别为对照(CK,0 g·m−2)、低氮处理(LN,5 g·m−2)和高氮处理(HN, 10 g·m−2),每个小区的面积为2.5 m ×6 m,缓冲带为1 m。在每个小区内设置1个0.5 m ×0.5 m的固定样方用于物候观测。所用含氮试剂为硝酸铵(NH4NO3),每年5月初,为了确保氮肥能够被植物有效吸收利用,并且保证氮肥随流水的淋失作用降到最小,在多云的阴天将氮肥施加到相应的梯度处理样方中[26]。
1.3 物候观测及花期计算
于2014年整个生长季(4月中旬− 10月初),每6~7 d 记录一次样方内出现的物种及物候期,每次记录独立于上次进行,共观测24次。本研究只关注始花期(first flowering date,FFD)和花期持续时间(flowering duration,FD)以及每次观测时群落中处于花期的物种丰富度,每个观测样方中,当杂草类植物出现可辨认的花瓣、花药和花丝时,记录为始花期;当禾草类植物的小穗只要不再被叶鞘包裹时,记录为始花期;当花瓣、花药、花丝或萼片开始枯萎时,记录为终花期[23,27],并以儒略历(Julian day,1月1日为1年中第1天)计数法计时。物种的始花期和终花期是处于该物候期的物种出现在开花样方中时间的平均值[28]。花期持续时间为对应样方中终花期时间与始花期时间的差值。
1.4 数据分析
在3种氮添加处理水平样方中均出现开花且在样本数量满足的情况下,以对照(CK)为基准,分别用低氮处理(LN)的数据和高氮处理(HN)的数据减去对照的数据,从而得到物种的花期提前与延迟的天数。用单因素方差分析(One-way ANOVA)检验氮添加对主要组分种和功能群(禾草类和杂草类)的始花期(FFD)和花期持续时间(FD)的影响。本研究中禾草类物种主要指禾本科物种,杂草类物种指除禾本科和豆科外的其他物种。用多重比较中的最小显著差数法(LSD method)比较始花期和花期持续时间在不同氮添加水平之间的差异(P<0.05)。始花期和花期持续时间之间的关系采用Pearson 相关分析进行验证。所有的分析和绘图分别在SPSS 25.0和OriginPro 2016中进行。
2 结果与分析
2.1 群落主要组分物种和功能群花期物候对氮添加的响应
相对于对照,施氮处理明显影响了群落中主要组分种的花期物候(图2和图3)。

图2 低氮添加处理对高寒草甸25个常见物种始花期和花期持续时间的影响Figure 2 Effect of low nitrogen addition treatment on the first flowering date and flowering duration of 25 common alpine species

图3 高氮添加处理对高寒草甸25个常见物种始花期和花期持续时间的影响Figure 3 Effect of high nitrogen addition treatment on the first flowering date and flowering duration of 25 common alpine species
低氮处理条件下,禾草类物种草地早熟禾、密花早熟禾(Poa poophagorum)、异针茅(Stipa aliena)、胡氏剪股颖、垂穗披碱草、羊茅(Festuca ovina)、芒剪股颖(Agrostis trinii)的始花期普遍性推迟(P<0.05);禾草类物种花期持续时间除垂穗披碱草显著延长(P<0.05)外,其余物种花期持续时间均有一定缩短,其中羊茅的花期持续时间显著缩短(P<0.05)。杂草类物种始花期和花期持续时间对施肥的响应因物种而异。菊科物种瑞苓草的始花期显著推迟(P< 0.05),花期持续时间缩短;毛茛科物种钝裂银莲花、高山唐松草(Thalictrum alpinum)、条叶银莲花、展毛翠雀花(Delphinium kamaonense)、丝叶毛茛的始花期显著提前(P<0.05),而钝裂银莲花和小花草玉梅的花期持续时间显著延长(P<0.05);龙胆科物种湿生扁蕾(Gentianopsis barbata)、华丽龙胆(Gentiana sino-ornata)和椭圆叶花锚(Halenia elliptica),玄参科物种短腺小米草(Euphrasia regelii)和毛果婆婆纳(Veronica eriogyne),伞形科物种细叶东俄芹(Tongoloa tenuifolia)和蔷薇科物种莓叶委陵菜(Potentilla fragarioides)的始花期显著提前(P< 0.05),其中细叶东俄芹、短腺小米草和毛果婆婆纳的花期持续时间显著延长(P<0.05)。
高氮处理条件下,禾草类物种除洽草(Koeleria cristata)和针茅(Stipa capillata)的始花期变化不显著(P> 0.05)外,其余物种始花期均显著推迟(P<0.05)。禾草类物种花期持续时间除垂穗披碱草外普遍缩短,其中异针茅和羊茅的花期持续时间显著缩短(P<0.05),垂穗披碱草的花期持续时间显著延长(P<0.05)。杂草类菊科物种瑞苓草的始花期显著推迟(P<0.05),花期持续时间缩短;毛茛科物种钝裂银莲花、丝叶毛茛、小花草玉梅、条叶银莲花和高山唐松草的始花期显著提前(P<0.05),花期持续时间显著延长(P<0.05);龙胆科物种湿生扁蕾、椭圆叶花锚和华丽龙胆的始花期显著提前(P<0.05),花期持续时间变化各异;玄参科物种毛果婆婆纳和短腺小米草,伞形科物种细叶东俄芹和蔷薇科物种莓叶委陵菜的始花期显著提前(P<0.05),花期持续时间显著延长(P<0.05)。
氮添加总体上导致禾草类物种的始花期显著推迟(LN和HN:P<0.01);杂草类物种始花期提前,其中在低氮处理水平下变化不显著(P>0.05),而在高氮条件下,杂草类物种始花期显著提前(P<0.05)(图4a)。禾草类物种花期持续时间在高氮处理下显著缩短(P<0.05);杂草类物种的花期持续时间在不同氮处理水平下均显著延长(LN:P<0.05;HN:P<0.01)(图4b)。
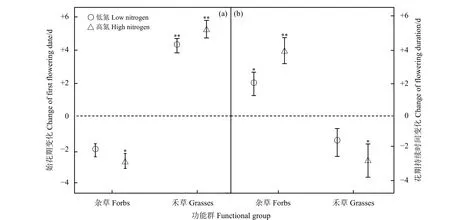
图4 不同氮添加处理下功能群始花期和花期持续时间的变化Figure 4 Changes in the first flowering date and flowering duration of the different functional groups under the different nitrogen addition levels
相关性分析结果显示(图5),自然群落(r=−0.41,P=0.006 7)、低氮处理群落(r=−0.49,P=0.004 6)和高氮处理群落(r=−0.47,P=0.013 2)的物种始花期与花期持续时间之间表现出显著的负相关性,表明物种开花时间越早,其花期持续时间越长,且在不同氮处理水平之间具有相同的趋势。
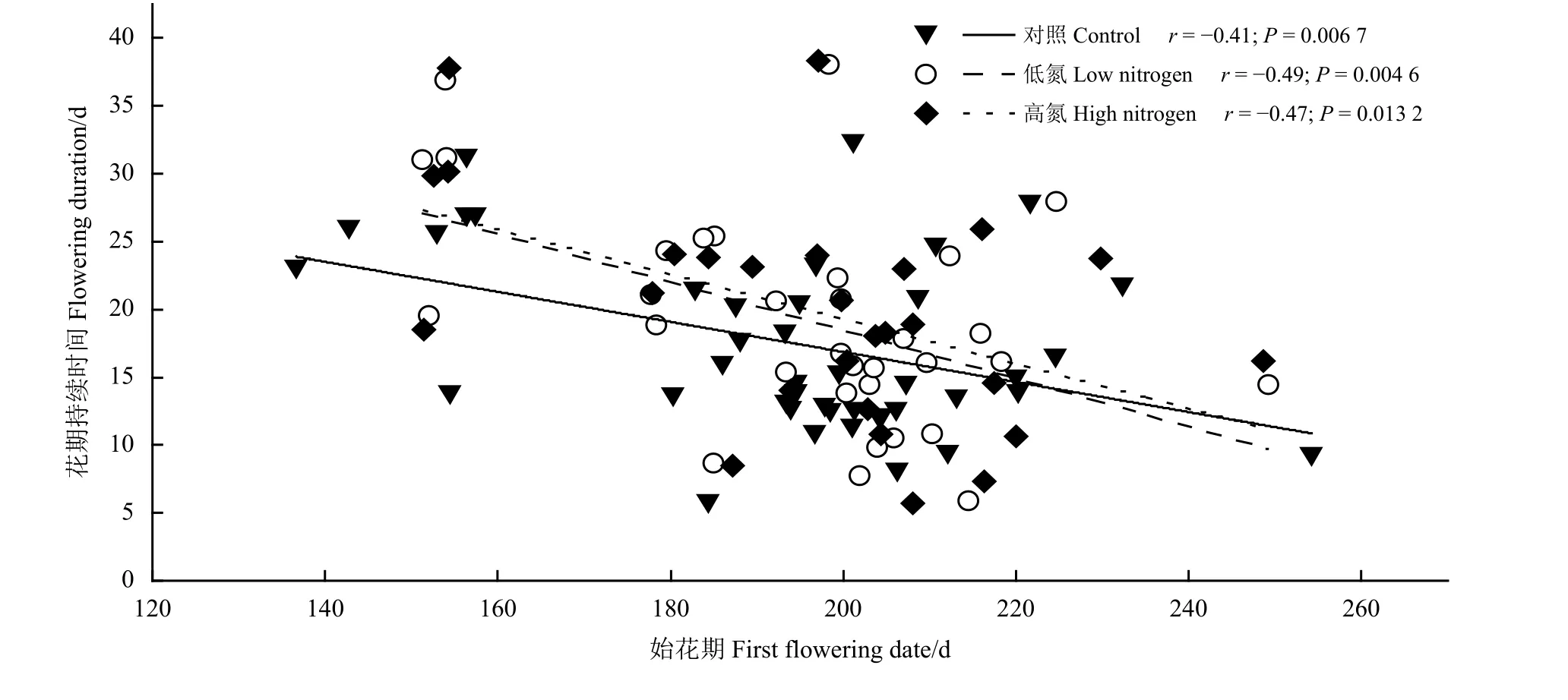
图5 氮添加条件下物种始花期和花期持续时间的相关性Figure5 Relationship between the first flowering date and flowering duration of thedifferent nitrogen addition levels
2.2 氮添加后群落花期物种丰富度的变化
对试验群落中开花物种物候观测发现,群落中花期物种遍及整个生长季,群落种间花期在生长季内呈连续交错趋势并持续5个月时间。大部分物种花期集中在7月− 8月,其中在自然群落中观测到49个物种开花,在低氮处理群落中观测到35个物种开花,在高氮处理群落中观测到25个物种开花。在3个处理中处于花期的物种多样性在整个生长季内的变化都是迅速的,从4月下旬群落返青伊始便进入花期,以较快的速度在6月末7月初达到顶峰后迅速下降,直到10月初生长季结束。群落中处于花期的物种多样性随时间呈峰型分布(图6)。在不同氮添加水平下,花期物种丰富度变化显著,且随着施氮量的增加群落中花期物种丰富度降低,每次观测时群落中开花物种组成的相似性随着施氮量的增加而降低,低氮群落和高氮群落中花期物种丰富度与自然群落相比分别降低了29%和49%。

图6 生长季群落中花期物种丰富度对氮添加的响应Figure 6 Species richness of flowering response to nitrogen addition during growth season
3 讨论
本研究表明,氮添加能够显著影响高寒草甸群落中25个主要组分种的花期物候,具体表现为禾草类物种始花期普遍推迟,开花持续时间缩短,而大部分杂草类物种始花期提前,花期持续时间延长。这一结果说明植物花期物候的变化除自身生活史特性决定外,环境因素变化也发挥着重要的作用。本研究中,物种花期物候的变化趋势类似于北美一年生草地群落[29]和内蒙古草原群落中所研究的花期物候变化结果[30],但是与高原苔原群落中所研究的结果相反,该地区研究发现氮添加导致禾草类植物始花期提前,而杂草类植物始花期推迟[18]。这说明氮添加对植物物候的影响在一定程度上依赖于植物所处的生境类型。
在不同生境条件下,同一功能类型的植物可能使用不同的生长策略。大量研究发现,相较于杂草类,氮添加更加有利于禾草类植物的生长[31]。植物从营养生长到生殖生长的转变是以生物量分配的变化和随后营养摄取减少为特征[32]。在营养丰富的生境中,植物通过摄取更多的营养和花费更长的时间用于营养生长[33],对于晚期开花物种来说更是如此。通过增加营养生长,禾草类物种对限制性光源和氮的竞争更具有优势,在施氮的样方中占据更加明显的优势地位,从而表明在高寒地区禾草类物种是良好的氮竞争者[34]。大多数杂草类物种开花较早,这种早开花策略有助于它们避免与禾草类植物进行激烈的光资源竞争[35]。因此,在施氮条件下,于生长季早期大多数杂草类物种花期提前可能有利于它们在后期生长中避免激烈的光竞争。
本研究中,氮添加使得禾草类物种推迟开花,这意味着植物从营养生长到生殖生长转变的推迟,在一定程度上表明植物加强营养生长或克隆生长[29]。相比之下,杂草类物种在氮添加条件下提前开花并延长花期持续时间,而开花持续期的延长意味着植物在花期时间生态位上有更多的时间吸引传粉者或实现成功传粉,从而有利于有性繁殖。相关性分析结果显示,物种花期持续时间与始花期之间呈现显著的负相关关系,这表明氮添加后,在生长季早期开花的物种较生长季中后期开花的物种来说,花期持续时间更长。高寒草甸群落中条叶银莲花、钝裂银莲花以及高山唐松草等物种在生长季早期开花,即使它们能够进行繁殖后营养生长,但是相对于生长季中后期开花物种的营养生长来说,其高度仍处于劣势,所以早期开花物种为了避免在生长季后期资源竞争(如光、传粉者等)中处于不利地位,采用了有利于其生殖生长的开花对策。本研究结果表明,在生长季前期物种始花期普遍提前,且花期持续时间延长,使得其能够在繁殖阶段充分利用环境中有限的营养资源。同时,由于在高寒地区生长季较短,晚开花物种受环境变化因素影响的风险增大(如霜降等)。因此早开花物种受限于营养生长,而迟开花物种通常受限于生殖生长。高寒草甸植物群落中大多数物种花期物候对氮添加的响应较为敏感,表现出不同的变化。由于群落中物种受生物、非生物以及其内在因素的影响,因此开花时间频繁地处在多种选择压力之下[36-37]。
长期物候观测研究表明,在融雪提前和冬季温度升高条件下,大多数植物始花期显著提前,这说明表型可塑性在植物开花过程中扮演着重要作用[38]。但是,也有研究发现植物物候过程主要由遗传因素决定[33]。植物对环境因素(如光周期、降水、温度、春化作用、融雪时间和土壤养分)的响应,引起开花时间的变化,是植物应对环境时空异质性变化的重要策略之一[39]。虽然氮是影响高寒草甸植物物候的重要环境因子,但是要详细地阐明其对不同植物物种和功能群物候的影响,还需进一步观察研究。同时,高寒草甸群落物种在时间生态位上的权衡,对于物种共存来说至关重要[40]。
群落中植物花期物候分布受多种环境因素共同作用[37],对环境变化十分敏感,而花期物候的变化可能会对生态系统结构与功能产生深远的影响[41]。本研究通过对群落中物种整个生长季的物候观测发现,物种间开花期呈现连续交错趋势,氮添加后群落中花期物种丰富度显著降低,低氮群落和高氮群落的花期物种丰富度相较于自然群落分别降低了29%和49%。氮是影响草地群落物种生长的主要限制元素[42],氮添加导致环境异质性降低,从而影响植物的功能性状和繁殖策略[43-44],当环境中氮含量超过一定值时则会造成物种多样性降低,进而改变群落的组成结构和自然格局[45-46]。在高寒区域植物生长期短暂,群落物候的微小变化足以改变种间物候补偿效应[47]。氮添加后群落盖度和高度的增加可能会阻碍传粉者的活动[48]。同时,传粉者与其访问的物种花期出现时间错位时也会降低传粉功能[49]。群落物种花期物候变化和群落水平花期物候分布格局的偏移会降低群落物候补偿[50],形成的时间生态位分化会进一步影响群落的物种组成,进而导致群落结构和功能发生变化。
