杉木无性系新品种‘洋020’和‘洋061’10年生幼龄材微观结构与力学性能的相关性*
2021-07-13孙海燕王玉荣赵荣军任海青
贾 茹 孙海燕 王玉荣 汪 睿 赵荣军 任海青
(1.中国林业科学研究院木材工业研究所 北京 100091; 2.中国林业科学研究院林业新技术研究所 北京 100091)
杉木(Cunninghamialanceolata)是我国人工林种植面积居于首位的针叶速生商品材树种,生长速度快、材质优良、木材纹理通直、结构均匀、不翘不裂,在建筑、家具制造和造船等领域有着较为广泛的应用(吴中伦, 1984)。通过杉木种内有性杂交得到变异群体,从中选拔优良个体育成的无性系品种,相较于种子混合繁殖来说,具有遗传增益较高、优良性状稳定、林相整齐等优点,在森林培育中被广泛采用(洪菊生等, 1993; 施季森等, 1994)。杉木种源-家系-家系内个体间在生长、材性的遗传变异及其性状相关性等方面已有较系统报道(施季森等, 1993),目前,杉木无性系研究主要集中在优良无性系选择、生长性状遗传评价等方面(胡德活等, 2001; 许忠坤, 2014; 段爱国等, 2014),而从木材加工利用角度对杉木无性系品种进行木材品质性状系统评价的报道较少。随着杉木无性系新品种的不断问世和大量造林应用,评价杉木无性系品种的木材品质性状,对于杉木木材的合理、高效利用以及优质高产杉木无性系品种选育和国家杉木大径材战略储备林基地建设等均具有重要意义。
木材力学性能是衡量木材品质性状的主要指标及评价木制品质量和加工性能的重要依据。杉木优良无性系生长与材质性状研究表明,木材力学性能也是筛选优良无性系的重要品质性状之一(朱安明等, 2015)。浙江富阳地区10年生杉木种源和无性系间在木材抗弯弹性模量、抗弯强度、顺纹抗压强度等木材力学性能上均存在显著遗传差异,总体趋势是杉木幼龄材的力学性能指标低于成熟材(骆秀琴等, 1994; 黄安民等, 2006)。成熟材与幼龄材相比,管胞次生壁的纵向弹性模量和硬度也明显不同(余雁等, 2006)。有研究指出,可通过幼龄材的特征参数预测成熟材材质(李坚等, 1999; 施季森等, 1993),利用杉木幼龄材与成熟材生长和材性等性状的相关关系可进行生长和材性性状的早期选择,从而缩短育种世代进程(施季森等, 1993; 韦立权, 2004)。不同针叶材树种成熟材和幼龄材的年龄界定不同,对于人工林杉木幼龄材和成熟材,其界定年龄是其生长期的第14~15年间(Mansfieldetal., 2009; 李坚等, 1999; 施季森等, 1993)。目前,虽有很多经过早期良种选育、生长性状良好的杉木无性系品种,但对杉木无性系幼龄期木材力学性能等材质性状的研究报道较少。
木材显微和超微构造特征以及化学组分是影响木材力学性能的主要因子,木材细胞壁厚度和壁腔比大小等显微构造可反映木材力学性能优劣(姜笑梅等, 1997)。细胞壁微纤丝角大小对木材硬度、弹性模量和强度性能指标的高低等影响显著,通常微纤丝角较小的木材具有较高的抗弯强度(刘一星等, 2004; 葛晓雯等, 2016; Tanabeetal., 2016)。木材纤维素结晶度也与木材弹性模量等力学强度性能密切相关(Wangetal., 2014; 范文俊等, 2015),一般来说结晶度越高,木材弹性模量、断裂强度、抗拉强度性能越好(Borregaetal., 2014; 丁涛等, 2012),反之亦然(范文俊等, 2015; 陈美玲等, 2016)。化学组分木质素的单体类型和含量对木材力学性能也有重要影响,如木材经过褐腐,细胞壁中的木质素降解后,木材硬度下降幅度较大(Wangetal., 2014); 杉木成熟晚材脱木质素后,会引起硬度和弹性模量降低(王传贵等, 2012)。研究还发现,杉木生物降解时木材化学组分、结晶度发生变化,其力学性能也会受到影响(Chenetal., 2017)。以往对杉木木材力学性能与微观结构的研究多集中于成熟材,而对于同一生境下不同杉木无性系品种幼龄材微观结构与力学性能的相关研究未见系统报道。
鉴于此,本研究以福建洋口国有林场杉木无性系品种比对示范试验林杉木无性系品种‘洋020’(C.lanceolata‘Yang 020’)和‘洋061’(C.lanceolata‘Yang 061’)10年生幼龄材为研究对象,应用光学显微图像分析系统、X射线衍射、傅里叶变换红外显微成像技术以及木材力学性能各项指标国标测试方法等对2个杉木无性系幼龄材的微观构造、木质素化学组分以及主要力学性能抗弯强度、抗弯弹性模量、顺纹抗压强度和硬度等进行研究,旨在表征和分析杉木无性系幼龄材的力学性能和微观结构特征,为杉木优良无性系选育、杉木木材加工利用和加工工艺研发提供科学依据。
1 材料与方法
1.1 试验材料
试材采自福建洋口国有林场(117°80′E,26°80′N)5块10年生杉木无性系品种比对示范试验林(100株块状小区,简比排列,重复3次)。原始选育群体源于1983年种内杂交试验林, 1991年从中初选优良单株,经扦插繁殖成系; 1992年建立无性系测定林(45个无性系,完全随机区组,4株小区,10次重复), 2006年复选15个无性系进行组培扩繁,先后在福建省内外建立示范林和区域测定林。其中,‘洋020’(编号: 国R-SC-CL-002-2018)和‘洋061’(编号: 国R-SC-CL-003-2018)于2019年通过国家林业和草原局林木新品种认定。‘洋020’具早期高生长迅速、胸径生长速度中上、侧枝较细短、树冠窄、自然整枝良好、树干通直圆满、耐轻度干旱和瘠薄立地条件、对杉梢螟(Diorictriaschuetzella)等虫害抗性较强等特征,宜适当密植并培育中小径材;‘洋061’的直径具持续生长优势,树高生长6~8年后加速持续至成熟期,树干通直圆满、尖削度小、出材量高,宜土层较深厚、肥沃湿润的立地造林并培育高规格大径材。
每个无性系各采伐8株标准木作为样木,共16株。其中,‘洋020’样木的平均树高14.58 m,胸径17.68 cm; ‘洋061’样木的平均树高11.88 m,胸径18.23 cm。各无性系取样木树高1~3.5 m间部分木段进行微观结构和力学性能测试。
1.2 试样制备
1.2.1 微观结构试样制备 截取样木树高1.8 m处70 mm厚圆盘,通过圆盘中心取2个垂直方向宽10 mm的中心条试样,分别编号A和B(图1)。中心条A和B沿径向从树皮至髓心分为内、中、外3部分。
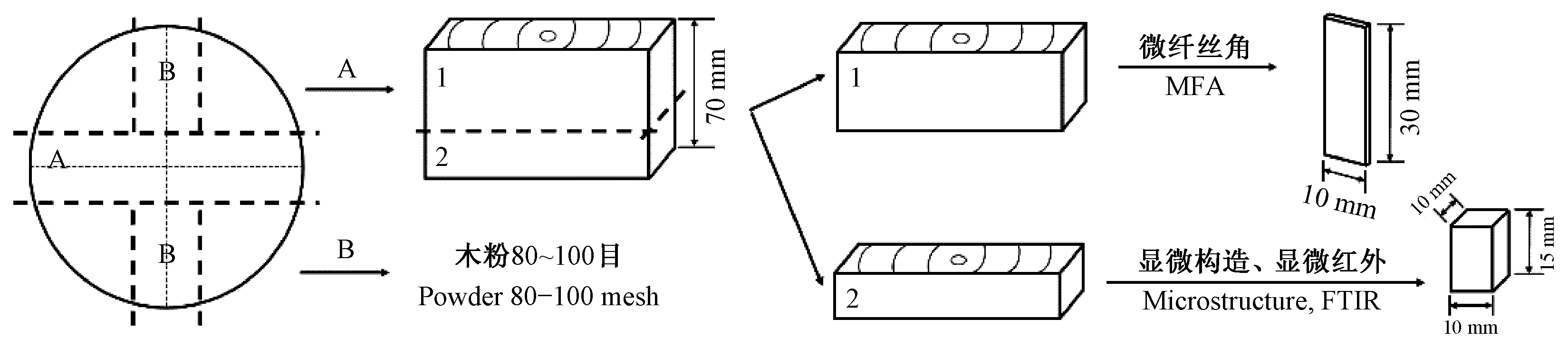
图1 木材微观结构试样制备示意Fig. 1 Sample preparation diagram of wood microstructure
中心条A截取为上(编号为1)和下(编号为2)2部分。上部中心条径向近树皮、中部和近髓心3部分分别锯成尺寸为30 mm×10 mm×1.5 mm(纵×径×弦)的木片,用于木材微纤丝角测量。下部中心条分别从径向3个不同部位取尺寸为15 mm×10 mm×10 mm(纵×径×弦)的小木块,用于显微结构观察和显微红外测试。中心条B 3个部位样品经粉碎研磨后取80~100目木粉,用于结晶度分析。
1.2.2 力学性能试样制备 将树高1~3.5 m样木分为3段,分段取样进行力学性能测试。近树皮(h1、h4)、中部(m1~m4)和近髓心(h2、h3)径向部位如图2所示。原木锯解后,按图2a先制备截面尺寸为30 mm×30 mm的毛坯条,再精加工成截面尺寸为20 mm×20 mm的试样,沿纵向制取2个试样,分别用于抗弯模量/强度和顺纹压缩强度测定,每个无性系8株样木制备抗弯和抗压无疵小试样各100个左右。

图2 抗弯、抗压和硬度试样制备示意Fig. 2 The diagram of wood bending, compression and hardness specimen preparation
硬度试样制备如图2b所示,先从原木试材中截取100 mm木段,制备截面尺寸为70 mm ×70 mm × 100 mm(径×弦×纵)的毛坯条,再精加工成截面尺寸为50 mm ×50 mm ×70 mm的试样,每个无性系8株样木制备硬度无疵小试样20个左右。
1.3 试验方法
1.3.1 显微构造观测 每个无性系选取3株样木,取样位置与宏观力学试样内、中、外3部分相对应。使用滑走切片机切取16 μm厚切片,番红染色,经脱水、透明制成永久切片。将切片置于ZEISS Imager A1显微镜下观察拍照,获得杉木不同无性系的显微结构。观测各试样横切面晚材管胞的双壁厚和腔径,获得其微观构造特征。
1.3.2 微纤丝角测定 选取上述30 mm×10 mm×1.5 mm的木片,使用荷兰PANalytical公司X-pert PRO多晶型X射线衍射仪测定每个无性系试样的微纤丝角,衍射角为22.4°,旋转范围为0~360°。测试时保证木片弦切面与X射线照射方向垂直,测试后得到002面衍射图,采用0.6T法计算各无性系木材试样的平均微纤丝角。
1.3.3 结晶度测定 使用荷兰PANalytical公司X-pert PRO多晶型X射线衍射仪测定每个无性系试样的结晶度。测试时将粉末样品置于样品盒中,压制平整,衍射仪扫描范围5°~55°,扫描速度6(°)·min-1。衍射图谱采用Segal经验方法计算结晶度,用公式C=(Iu-Ia)/Iu×100%(Iu为2θ=22°处的极大积分强度,Ia为2θ=18°处的极小积分强度)计算各无性系结晶度的平均值。
1.3.4 傅里叶变换红外显微原位分析 选用与显微构造观测相同的样品,使用滑走切片机切取14 μm厚切片,置于傅里叶变换红外显微成像系统透射模式下,分别对每个无性系近树皮、中部和近髓心3个部位样品进行原位检测,每个位置至少随机选取3个区域(面积为130 μm×130 μm)作为样品内观察重复。红外光谱数据分析时,每个区域至少选取15个点产生的红外光谱,并在红外光谱的1 800、1 540和785 cm-1处进行基线校准,图谱经归一化处理后用峰比值法比较木质素相对含量。
1.3.5 力学性能测试 按照GB/T 1928—2009规定,首先在恒温(20±2)℃、恒湿65%±3%的环境下平衡木材试样含水率,当试样含水率调至约12%时,利用万能力学试验机测定不同无性系木材的抗弯模量、抗弯强度、顺纹压缩强度和硬度,测定方法参照我国现行国家标准(GB/T 1936.2—2009; GB/T 1936.1—2009; GB/T 1935—2009; GB/T 1941—2009)和其他通用技术方法。
2 结果与分析
2.1 2个杉木无性系幼龄材微观结构
2.1.1 显微构造特征比较 2个杉木无性系幼龄材近树皮处,即第8个年轮的显微构造如图3所示。10年生杉木木材主要由管胞和木射线等构成,年轮间界限明显; 早晚材管胞形态差异明显,早材管胞壁薄腔大,多为卵圆形,晚材管胞壁厚腔小,多为长方形。‘洋061’和‘洋020’早材管胞形态较为接近,晚材部分管胞形态有较大区别,相比于‘洋061’,‘洋020’晚材带管胞数量多,且管胞壁较厚,腔较小。

图3 无性系‘洋061’(a)和‘洋020’(b)木材横切面显微构造Fig. 3 Cross-sectional microstructure of wood between ‘Yang 061’(a) and‘Yang 020’ (b)
2个杉木无性系木材早、晚材带管胞平均双壁厚、腔径和壁腔比如表1所示。‘洋020’和‘洋061’管胞平均双壁厚分别为4.28和4.29 μm,略小于浙江安吉龙山林场26年生杉木无性系幼龄材(Songetal., 2011)。2个杉木无性系早材之间管胞形态差异较小,‘洋061’和‘洋020’早材管胞平均双壁厚分别为3.20和3.41 μm,壁腔比相差不大。对比2个无性系木材晚材管胞解剖参数发现,‘洋061’和‘洋020’晚材区管胞平均双壁厚分别为4.93和5.39 μm,腔径分别为21.20和18.55 μm,方差分析结果表明均达到0.05显著性差异水平。‘洋020’晚材区管胞较‘洋061’呈壁厚腔小的特点,‘洋020’晚材区管胞壁厚较‘洋061’约大9%,管胞腔约小13%,壁腔比约大25%。林木生长特性与组成细胞形态关系密切。对比‘洋061’和‘洋020’生长特性,10年生‘洋061’的胸径稍大,径向生长较快,年轮较宽,相较于‘洋020’,其管胞特别是晚材区管胞呈腔大壁薄的特点。这些显微构造特征不同有可能导致杉木无性系木材的力学性能差异。

表1 2个杉木无性系木材解剖构造参数①Tab.1 Anatomical structure parameters of woods between two Chinese fir clones
2.1.2 微纤丝角比较 2个杉木无性系木材薄片的X射线衍射图谱如图4所示。杉木无性系间木材薄片的X射线衍射图谱大体趋势一致,但出现峰值最高点位置和衍射峰半高宽不同,这表明杉木无性系间在细胞壁微纤丝排列角度上有一定差异。采用0.6T法对2个无性系木材薄片的衍射图谱进行分析发现,木材平均微纤丝角在12°~15°之间,其中‘洋020’细胞壁的微纤丝角比‘洋061’小18%,2个无性系的微纤丝角差异达显著性差异水平(P<0.05,表2)。总体上,杉木幼龄材的微纤丝角要大于36年生杉木无性系近成熟材的平均微纤丝角(Yinetal., 2011)。

表2 2个杉木无性系木材的微纤丝角统计分析Tab.2 Statistical results of microfibril angle of wood cell walls between two Chinese fir clones

图4 2个杉木无性系间木材薄片的X射线衍射图谱Fig. 4 XRD patterns between two Chinese fir clones by wood flake
木材微纤丝角大小与树种间、树种内品种间等遗传因素有关,还可能与木材取样的树龄、部位以及经营措施等环境因素有关。美洲黑杨(Populusdeltoides)不同基因型的生长性状与木材微纤丝角相关研究表明,生长较快的杨树其木材也具有较大的微纤丝角(Wangetal., 2016)。对辐射松(Pinusradiata)不同无性系的微纤丝角研究发现,在相似立地条件下,微纤丝角大小受基因型控制(Donaldson, 1992)。‘洋061’与‘洋020’2个基因型具有不同的树高和胸径生长,在微观结构上微纤丝排列角度不同。微纤丝角大小通常与木材力学性能呈负相关关系。10年生时,径向生长稍慢的‘洋020’的微纤丝角较小,推测比‘洋061’具有更好的木材力学性能,但杉木无性系基因型间幼龄材微纤丝角的差异是否能够保持到成熟材阶段,有待进一步观察。
2.1.3 结晶度比较 2个杉木无性系木材粉末的X射线衍射图谱如图5所示。2个杉木无性系均在2θ为18°、22.5°和35°附近有3个衍射峰,分别对应(101)、(002)和(040)晶面。(002)衍射的极大峰值在2θ=22.5°附近,极小值在2θ=18°附近。2个杉木无性系木材粉末结晶度的X射线衍射图谱形状基本相同,但在2θ=22.5°处的峰强度明显不同,且半高宽差别较大,这说明无性系间纤维素结晶部分晶胞构造是相同的,但结晶程度不同。‘洋061’细胞壁纤维素结晶度较低,为35.88%±4.36%,‘洋020’ 细胞壁纤维素结晶度较高,为39.73%±4.58%,与浙江开化来源于不同地理种源的多个杉木无性系成熟材(其平均结晶度为43%~46%)(胡梦霄等, 2017)比较,‘洋020’和‘洋061’幼龄材的结晶度均较小,这可能与2个无性系试材尚处于幼龄期有关。无性系‘洋020’的结晶度虽然比‘洋061’高11%左右,但2个无性系间的结晶度测定值未达统计显著性差异标准。

图5 2个杉木无性系间木材粉末的X射线衍射图谱Fig.5 XRD patterns between two Chinese fir clones by wood powder
2.1.4 化学组分——木质素比较 采用傅里叶变换红外显微原位分析技术测定木质素含量。表3所示为1 800~800 cm-1的红外特征峰及其归属,其中1 600、1 508、1 264和1 232 cm-1处均为木质素的吸收峰(Faix, 1991; Åkerholmetal., 2003)。图6所示为杉木晚材管胞壁可见光图像和全光谱图像。

表3 针叶材细胞壁聚合物的傅里叶红外特征峰归属Tab.3 Fourier infrared characteristic peak of coniferous wood cell wall polymer

图6 杉木晚材管胞壁可见光图像(a)和全光谱图像(b)Fig. 6 Visible light image (a) and full spectrum image (b) of latewood tracheid walls in Chinese fir clones
图7所示为2个杉木无性系木材管胞壁平均红外光谱,图谱经基线校正和归一化处理后用于后续分析。对红外图谱中的特征峰进行比较分析,采用I1 508/I1 452、I1 508/I1 424、I1 600/I1 452和I1 600/I1 424表征木质素的相对含量,2个杉木无性系木材的红外光谱特征峰高比列于表4。对比4组特征峰高比发现,‘洋020’的峰值高于‘洋061’,其I1 508/I1 452、I1 508/I1 424、I1 600/I1 452和I1 600/I1 424分别比‘洋061’高4%、5%、11%和12%。在林龄、立地条件和营林措施相同的条件下,2个无性系木质素含量不同,表明在木材次生生长过程中细胞壁木质素生物合成、沉积等动态发育模式显然不同,但无性系间木材细胞壁动态发育模式不同或遗传差异在木材细胞壁形成中的科学意义还有待进一步研究。在其他树种,如针叶材湿地松(Pinus elliottii)和阔叶材柳树(Salix L.)的研究中,也发现不同无性系间存在木质素含量差异(王晓荣等,2016;李景涛等,2017)。木质素作为细胞壁重要化学组分之一,填充于纤维素骨架中,具有增强植物体机械强度和维持细胞正常形态的重要作用,会影响木材力学性能。脱除杉木成熟晚材木质素后,其硬度和弹性模量显著降低(王传贵等,2012)。本研究2个无性系木材木质素含量不同有可能导致杉木无性系木材的力学性能差异。

图7 2个杉木无性系木材管胞壁平均红外光谱Fig. 7 Average IR spectra of wood tracheid walls between two Chinese fir clones
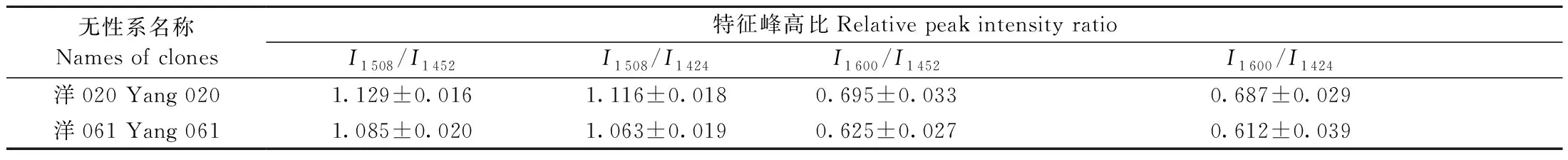
表4 2个杉木无性系木材的红外光谱特征峰高比Tab.4 Ratio of IR spectra characteristic peak values of woods between two Chinese fir clones
2.2 2个杉木无性系幼龄材力学性能
2.2.1 抗弯与抗压性能比较 由表5可知,2个杉木无性系木材的抗弯强度、抗弯弹性模量和顺纹抗压强度平均值分别为46.96 MPa、9.58 GPa和28.74 MPa。‘洋020’木材抗弯强度、抗弯弹性模量和抗压强度均较大,分别比‘洋061’高17%、12%和10%左右。2个无性系木材的力学性能接近于在福建采集的24年生杉木(余光等, 2014),但抗弯强度和顺纹抗压强度低于在江西大岗山采集的36年生杉木幼龄材(任海青等, 2006),‘洋020’抗弯弹性模量较其高。统计分析表明,2个杉木无性系幼龄材抗弯强度、抗弯弹性模量和抗压强度均存在显著性差异,说明依据杉木无性系幼龄材力学品质性状进行力学性能优良杉木品种的早期选择具有潜在可行性。

表5 2个杉木无性系木材主要力学性能统计分析Tab.5 Statistical results of wood main mechanical properties between two Chinese fir clones
2.2.2 硬度比较 由表5可知,2个杉木无性系木材硬度平均值为1 413 N,‘洋020’较‘洋061’高10%左右,无性系间硬度达到显著性差异水平。对比表6中各端面硬度发现,2个杉木无性系木材三面硬度中,均为端面最大、弦面其次、径面最小。

表6 2个杉木无性系木材不同切面硬度统计分析Tab.6 Statistical results of wood different section hardness between two Chinese fir clones
2个无性系木材端面硬度不存在显著性差异,弦面和径面硬度均具有显著性差异。目前,关于杉木木材硬度的研究报道较少,硬度样品要求尺寸较大,不易获取试样且加工测试费时较多可能是其中原因之一。木材硬度表征木材抵抗其他刚体压入木材的能力,也可反映其耐磨损能力。对比2个无性系木材硬度可见,硬度较高的‘洋020’木材应具有较好的耐磨能力。
2.2.3 综合强度与分级比较 木材综合强度常用木材顺纹抗压和抗弯强度的总和表征。2个杉木无性系平均综合强度为75.70 MPa,‘洋020’ 和‘洋061’的综合强度分别为81.63和69.76 MPa,‘洋020’平均综合强度高于‘洋061’15%左右。
为了确定2个优良杉木无性系的力学性能相对于我国人工林树种所处水平,根据我国现有木材强度分级标准(成俊卿等, 1992; 尹思慈, 1990; 李坚等, 1999),2个杉木无性系10年生木材的抗弯、抗压性能及综合强度等级均为低等,硬度为甚软(表7)。综合对比2个无性系,‘洋020’的木材力学性能高于‘洋061’,且‘洋020’各力学性能指标的变异系数小于‘洋061’,其幼龄材力学性质相对较优。

表7 2个杉木无性系微观结构与力学性能指标Tab.7 Index values of microstructures and mechanical properties between two Chinese fir clones
3 讨论
据最新全国森林资源清查报告,杉木人工林面积、立木蓄积总量均居用材人工林首位,加速优质、高产杉木品种培育,提高木材产量和品质,是国家应对木材供应安全的迫切需求。杉木无性系‘洋020’和‘洋061’为不同遗传背景的基因型,是按照不同选育目标入选的。‘洋020’主要特性是幼龄期高生长显著,树冠较窄,适于密植和中小径材培育; 而‘洋061’直径速生性启动早于树高生长,主干粗壮,尖削度小,树冠浓密,呈尖塔形,树高生长6~8年后开始加速并维持到主伐,单株立木蓄积超过‘洋020’20%以上,适合培育高规格大径材。杉木木材解剖构造参数、微纤丝角和结晶度等参数,可能随不同遗传背景品种的木材形成发育模式不同、林龄和采样地异质性改变而有所变化。本研究在杉木无性系品种对比示范试验林中采集试样,较好地控制了立地条件和林龄的异质性,虽然还不能确切预测出成熟材阶段的木材品质性状,但‘洋020’和‘洋061’幼龄材显微构造参数、微纤丝角、结晶度、木质素含量等微观结构和化学组分与力学性能的相关性及其品种间的显著差异性,为适应不同营林目标需求优良杉木新品种的培育提供了新视野。
3.1 显微构造参数与力学性能的相关关系
对比2个杉木无性系幼龄材显微构造发现,2个品种早材管胞形态特征区别不大,明显区别在于晚材管胞形态特征。‘洋020’晚材带较宽、管胞呈腔小壁厚的特点,壁腔比较‘洋061’高25%左右,抗弯强度、抗压强度、抗弯弹性模量、综合强度和硬度均高于‘洋061’。2个杉木无性系幼龄材管胞壁厚度以及壁腔比等解剖构造参数与杉木力学性能指标呈正相关,特别是晚材的显微构造特征显著影响杉木无性系幼龄材力学性能指标高低。黄艳辉(2007)关于杉木材性和姜笑梅等(2002)对湿地松的研究中也发现了相似规律。因此本研究认为,在开展杉木无性系早期选择时,作为承载木材各种力学性质载体的细胞壁特征,尤其是晚材管胞壁厚和胞腔大小,是材性育种早期选择需要密切关注的重要性状。
3.2 微纤丝角与力学性能的相关关系
2个杉木无性系幼龄材平均微纤丝角在12°~15°之间,‘洋020’的平均微纤丝角较‘洋061’小18%。对比2个无性系幼龄材的微纤丝角与力学性能发现,杉木无性系幼龄材微纤丝角大小与木材力学性能呈负相关关系。安徽产25年生和江西产36年生人工林杉木也表现出微纤丝角大小与MOR、MOE抗弯性能呈负相关关系(黄艳辉, 2007; Yinetal., 2011),且在对不同针叶材树种如辐射松、日本落叶松(Larixkaempferi)等的研究中也发现了相同规律(Barriosetal., 2017; Sunetal., 2017)。针叶材鱼鳞云杉(Piceajezoensis)的微纤丝角与抗弯性能径向变异研究结果表明,幼龄材部分微纤丝角与弹性模量的相关性大于成熟材(Tanabeetal., 2016),这预示着针叶材幼龄材微纤丝角与木材力学性能有较强相关性。因此本研究认为,杉木幼龄材细胞壁微纤丝角大小也是决定杉木力学性能优劣的重要影响因子之一。
3.3 结晶度与力学性质的相关关系
2个杉木无性系幼龄材纤维素结晶度在35%~40%之间。经比较发现,力学性能较优的‘洋020’的结晶度较‘洋061’约高11%,但2个无性系的结晶度未达到统计上的显著性差异。以往研究表明,结晶度大小与纤维定向性优劣以及木材抵御外力的能力相关(Barriosetal., 2017; 陈美玲等, 2016),结晶度与木材的力学性质呈正相关(Borregaetal., 2014)。相对于杉木成熟材,10年生杉木无性系幼龄材纤维素结晶度较小,这是因为幼龄期的杉木还处于木材形成的生长发育阶段,纤维素的定向性还未完全稳定,因而幼龄材纤维素结晶度高低不是影响杉木幼龄材力学性能最主要的结构因子。
3.4 木质素含量与力学性质的相关关系
无性系‘洋020’木质素4组特征峰高比平均值相较于‘洋061’高8%左右。作为细胞壁的结壳物质,木质素赋予了木材硬度和刚性。尽管本研究杉木无性系试样取自幼龄材阶段,木质素还处于不断沉积过程中,但木质素含量高低也与木材力学性能呈正相关关系,‘洋020’幼龄材具相对较高的抗弯强度、抗弯弹性模量、顺纹抗压强度和硬度。以往对杉木成熟材的力学性能研究表明,木质素含量与木材抗弯强度、弹性模量和硬度呈显著正相关关系(Chenetal., 2017; 王传贵等, 2012)。本研究发现,在杉木幼龄材阶段,不同杉木无性系木质素含量差异与抗弯性能、抗压性能和硬度呈正相关关系。因此本研究认为,细胞壁木质素含量与微纤丝角和解剖构造等可能协同影响杉木幼龄材的力学性能。
4 结论
本研究在福建洋口国有林场生境一致的杉木无性系品种比对试验示范林中采集‘洋020’和‘洋061’无性系标准株样木,测定其10年生幼龄材的微观结构和力学性能,并对微观结构与力学性能的相关性进行分析,主要结论如下。
1) 与‘洋061’相比,‘洋020’具有晚材区宽、晚材管胞壁厚、壁腔比大、细胞壁微纤丝角小、细胞壁结晶度和木质素含量高等特点,2个无性系幼龄材解剖构造参数与微纤丝角微观结构因子指标达到统计上的显著性差异。
2) 参照我国主要树种木材强度等级标准,杉木幼龄材的抗弯和抗压性能等级为低等,硬度为甚软; 但‘洋061’和‘洋020’杉木无性系间幼龄材抗弯强度、抗弯弹性模量、顺纹抗压强度和硬度等力学性能指标存在统计上的显著差异,‘洋020’幼龄材力学性能高于‘洋061’。
3)‘洋061’和‘洋020’杉木无性系幼龄材管胞壁厚度、壁腔比、结晶度和木质素含量与力学性能呈正相关关系,微纤丝角与力学性质呈负相关关系。
综上可知,幼龄材解剖构造参数、微纤丝角等差异是影响杉木无性系品种间幼龄材力学性能的主要因子,‘洋061’和‘洋020’幼龄材解剖构造参数、微纤丝角指标可反映幼龄期品种间木材性状发育的遗传差异,研究结果可为杉木无性系品种木材品质早期选择育种和杉木木材加工利用提供科学依据。
