楸木早/晚材水分吸着与湿胀行为*
2021-07-13欧阳白蒋佳荔
欧阳白 李 珠 蒋佳荔
(中国林业科学研究院木材工业研究所 国家林业和草原局木材科学与技术重点实验室 北京 100091)
楸(Catalpabungei)为紫葳科(Bignoniaceae)梓树属(Catalpa)落叶乔木,其树干高大、通直圆满、材质优良,主要用于生产家具、门窗、地板和乐器等。为加工出优质产品,学界已开展了宏观尺度下楸木干缩/湿胀性质的初步研究(刘盛全等, 2008; 麻文俊等, 2013; 吴玮, 2015),但研究结果局限于获取无疵试样的干缩率和湿胀率等参数上,尚缺乏在生长轮尺度上对楸木吸湿过程中湿胀特性的深入探讨。
木材的绝大多数细胞和组织平行于树干,沿轴向呈近似同心环状排列,因此可将木材视为具有3个对称轴(轴向L、径向R和弦向T,其中径向和弦向常统称为横向)的正交异向性材料(Bodigetal., 1993)。木材的湿胀行为也具有正交异向性,木材细胞的轴向湿胀率很小,比横向湿胀率约小一个数量级,如相同条件下欧洲云杉(Piceaabies)管胞的轴向湿胀应变为0.014 μm,而横向湿胀应变为0.170 μm(Joffreetal., 2016)。针对木材湿胀正交异向性,目前主要集中在木材横切面上径向与弦向之间湿胀的差异研究(Pangetal., 2005; Bonarskietal., 2015; Joffreetal., 2016),由于早材与晚材的密度、壁层构造、化学成分、微纤丝角等不同(Bertaudetal., 2004; Chauhanetal., 2004; Dongetal., 2010; Pateraetal., 2013; 2018; Joffreetal., 2016),且二者在径向和弦向上分别以串、并联的方式连接,因此当木材细胞壁中的水分增加时,早材与晚材的湿胀行为存在差异,并会发生彼此间的相互作用(Maetal., 2006; Deromeetal., 2011; Rafsanjanietal., 2012a; 2012b)。
利用落射荧光显微镜、共聚焦激光扫描显微镜、环境扫描电子显微镜和X射线断层扫描显微镜等技术,可将木材的湿胀行为研究从宏观尺度(无疵试样)拓展到介观(生长轮)和微观(细胞壁)尺度(Pangetal., 2005; Maetal., 2006; Taguchietal., 2010; Joffreetal., 2016; Pateraetal., 2018)。那么,针对同一生长轮内早材和晚材的湿胀行为,研究对象有2类: 一是包含早材和晚材组织的生长轮; 二是同一生长轮内的独立早材和独立晚材。对欧洲云杉的研究结果表明,同一生长轮内晚材比早材的径向湿胀率大,早、晚材的弦向湿胀率无明显差异(Pateraetal., 2018); 早、晚材的差异湿胀(弦向/径向)分别为2.50和1.32(Lanvermannetal., 2014)。Pang等(2005)指出,辐射松(Pinusradiata)径向和弦向湿胀率均表现为独立早材小于独立晚材; Taguchi等(2010; 2011)认为,鱼鳞云杉(Piceajezoensisvar.microsperma)独立早材与独立晚材的弦向湿胀行为相似,但径向湿胀行为存在明显差异: 独立早材的湿胀率随含水率增加而减小,独立晚材的湿胀率则随含水率增加呈或增加、或不变、或减小的不规则现象,且这种现象随着细胞数量增加而愈加明显。
生长轮尺度上的早材和晚材湿胀行为是木材-水分关系研究的热点之一,但当前已开展的研究存在一定局限性,如研究对象多为针叶材,少有针对阔叶材的研究; 无法实时、同步、准确获取含水率与尺寸变化的对应数据,缺乏“早材和晚材湿胀应变”与“含水率变化”是否同步的研究。鉴于此,本研究以阔叶材楸木为研究对象,采用动态水分吸附分析仪联用白光显微镜成像技术,开展基于生长轮的早/晚材湿胀行为可视化研究,以期从生长轮尺度上揭示早/晚材的径向、弦向湿胀行为规律及其相互作用机理,阐明“湿胀行为”与“平衡含水率”之间是否存在滞后现象。
1 材料与方法
1.1 试验材料
以44年生楸木心材部位的第32生长轮为试验材料,试样锯解和制备方法如图1所示。利用X射线剖面密度仪(德国EWS,DENSE-LAB)测得第32生长轮早材和晚材的全干密度分别为0.33和0.48 g·cm-3。使用滑走切片机分别制备生长轮试样(growth ring,ELW)、早材试样(earlywood,EW)和晚材试样(latewood,LW),其中,生长轮试样尺寸为4 mm(L)×4 mm(T)× 8 mm(R)、早材试样尺寸为4 mm(L)× 4 mm(T)× 2.5 mm(R)、晚材试样尺寸为4 mm(L)× 4 mm(T)× 5 mm(R)。每种试样3个,质量均为(50±1)mg。在恒定室温(25 ℃)状态下,将制备好的试样置于装有五氧化二磷(P2O5)的干燥器中干燥,直至试样质量不再发生变化。
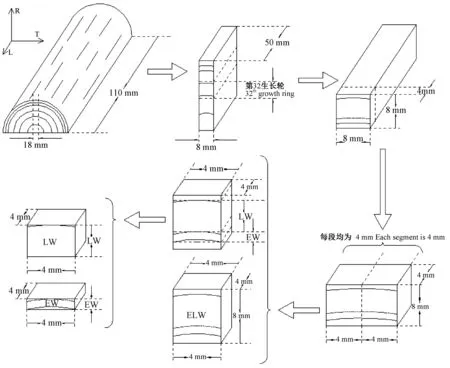
图1 试样锯解示意Fig. 1 Sketch map of sample sawing
1.2 试验方法
1.2.1 水分吸着等温线测量 采用动态水分吸附分析仪(英国Surface Measurement Systems,DVS Resolution)测量水分吸着等温线。温度设置为(25±0.1)℃,相对湿度(relative humidity,RH)范围为0%~90%,以10%相对湿度为梯度增量。每隔20 s测量1次试样质量。在每个恒定的相对湿度梯度,测试过程分为水分吸着阶段和平衡含水率恒定阶段: 在水分吸着阶段,当试样质量变化每10 min小于0.002%时,认为试样达到含水率平衡态,阶段结束点设为T1,该阶段测量生长轮、早材和晚材的径向和弦向湿胀应变量; 在平衡含水率恒定阶段,以试样达到含水率平衡态的时间点(T1)开始,相对湿度不变继续恒定360 min,阶段结束点设为T2,该阶段分别测量T1和T2时间点所对应的试样湿胀应变量。
1.2.2 湿胀应变量测量 在水分吸着等温线测量期间,采用动态水分吸附分析仪联用白光显微镜成像技术(台湾AnMo Electronics Corporation,Dino X Lite Digital Microscope)同步测量生长轮、早材和晚材的径、弦向湿胀应变量,每隔1 min拍摄1张图像,像素为1 280×960。通过比对图像中试样尺寸变化,计算得到相应的湿胀应变,同时建立试样含水率变化与尺寸变化之间的同步对应关系。
1.2.3 图像信息转换为湿胀应变量 早材试样定义为独立早材EW(图2a),晚材试样定义为独立晚材LW(图2b),生长轮试样定义为生长轮ELW(图2c),生长轮试样内的早材组织定义为生长轮内早材ELW-E(图2c),生长轮试样内的晚材组织定义为生长轮内晚材ELW-L(图2c)。
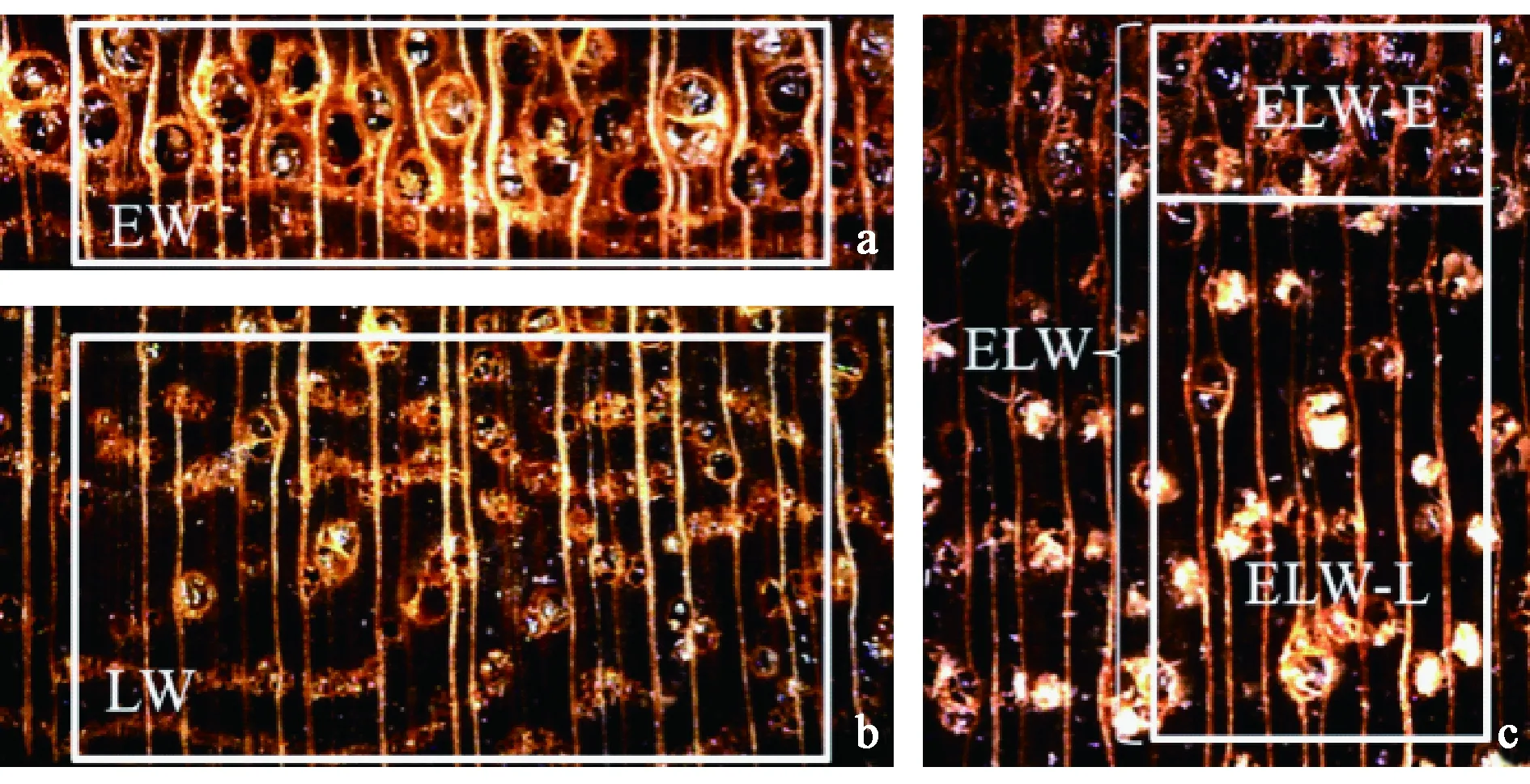
图2 试样湿胀应变的测量区域示意Fig. 2 Sketch map of measurement area of sample swelling
采用Adobe Photoshop(Adobe Systems Incorporated)和AutoCAD(Autodesk)对水分吸着阶段和平衡含水率恒定阶段采集的系列图集进行降噪和测量处理,将图像信息转换为湿胀应变量数据: 首先,在轮界线附近找到4个管孔,放大至像素块可见; 然后,在像素块的明显分界处作标记点,记录4个标记点的位置(图3); 最后,测量4个标记点间的距离,即得出试样径向和弦向湿胀应变。通过下式计算试样径向和弦向湿胀率:

图3 图像处理示意Fig. 3 Sketch map of image processing
(1)
式中:εRH为试样在任一相对湿度环境中(径向/弦向)的湿胀率,%;lRH为试样在任一相对湿度环境中达到平衡含水率时(径向/弦向)的尺寸,μm;l0为绝干试样(径向/弦向)的尺寸,μm。
2 结果与讨论
2.1 早材和晚材的水分吸着等温线
图4所示为独立早材EW(图4a)、独立晚材LW(图4b)和生长轮ELW(图4c)的水分吸着等温线。从3个重复试样之间的数据重现性上看,EW之间存在一定差异,最大差值为3.33%; 而LW和ELW的数据重现性比较一致,最大差值仅分别为0.34%和0.45%。由于楸木早材导管直径约为晚材的10倍,早材导管内通常含有大量侵填体,侵填体的存在很大程度上降低了早材细胞壁的水分吸着能力,且早材导管内的侵填体含量与分布存在差异(Lietal., 2019),从而使得不同早材试样之间的平衡含水率也呈现差异(图4a)。对于楸木晚材,导管直径较小,仅有少数导管内含有侵填体(吴玮, 2015),且晚材率高达76.9%,因此与EW相比,LW和ELW的水分吸着等温线受侵填体的影响较小,数据重现性较好(图4b、c)。
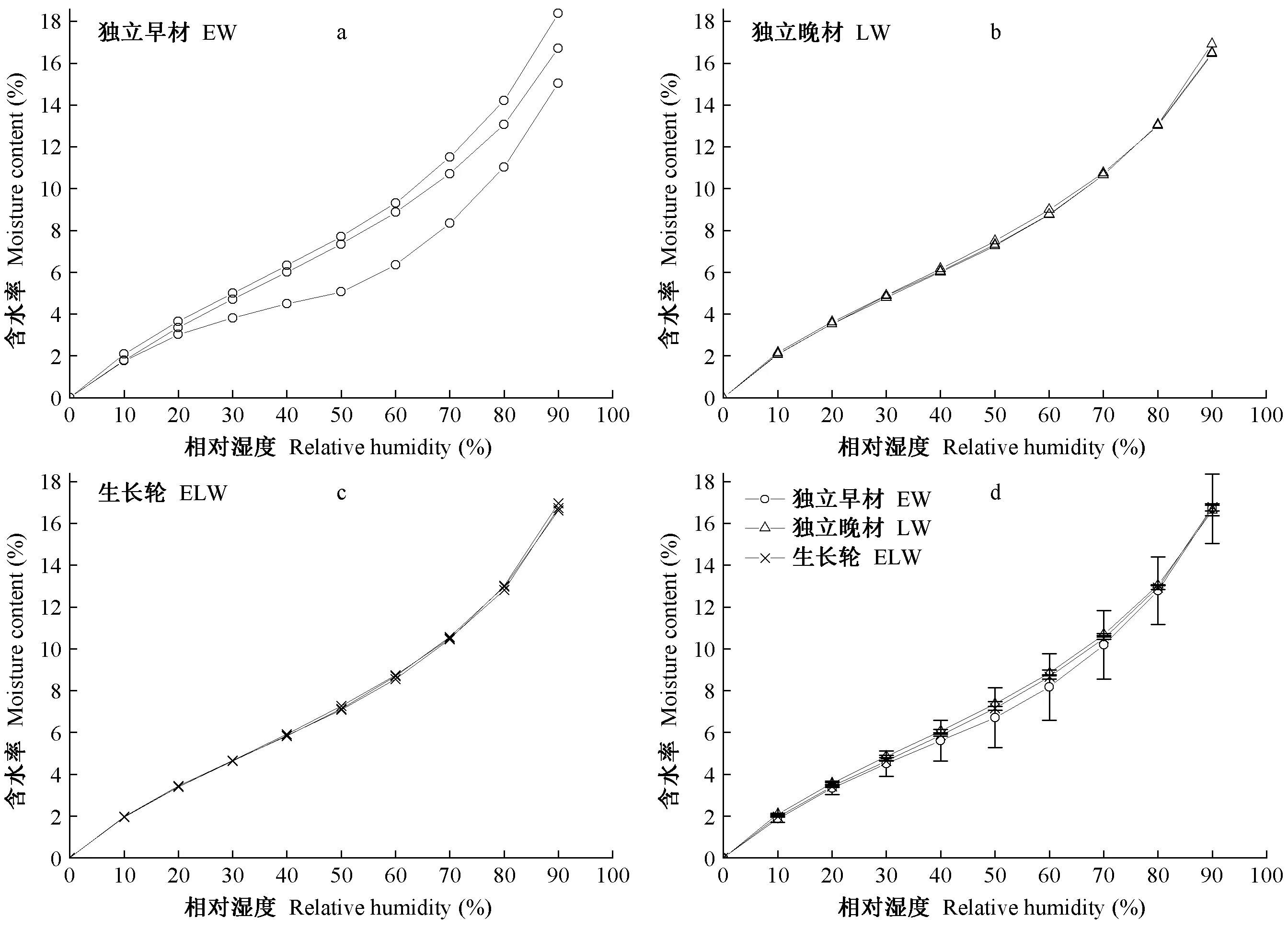
图4 独立早材、独立晚材和生长轮的水分吸着等温线(25±0.1 ℃)Fig. 4 The sorption isotherm of earlywood, latewood and the growth ring
图4d所示为EW、LW和ELW之间的水分吸着等温线平均值。在相对湿度0%~90%的水分吸着过程中,EW、LW和ELW之间的平衡含水率差值呈先增大后减小的变化趋势; 在任一相对湿度水平下,三者之间的平衡含水率差值小于0.66%,LW的平衡含水率最高,EW的平衡含水率最低。推测是因为在同一生长轮内,晚材的纤维素含量往往略高于早材,而木质素含量略低于早材(Bertaudetal., 2004),木材中无定形纤维素的吸湿性显著高于木质素(Kollmannetal., 1968; 周海珍等, 2017)。当相对湿度从50%升至90%时,EW与LW之间的平衡含水率差值从0.65%下降至0.03%。推测是由于早材微纳米孔数量较多,在高湿条件下形成了更多的毛细管凝结水。Pang等(2005)对辐射松早材和晚材的吸着等温线研究也发现了类似情况。
2.2 早材和晚材的径向湿胀行为
图5所示为独立早材(EW)、独立晚材(LW)、生长轮内早材(ELW-E)、生长轮内晚材(ELW-L)和生长轮(ELW)径向湿胀应变量随相对湿度的变化情况。可以看出,无论独立试样亦或生长轮试样的内部组织,晚材的湿胀应变量均大于早材,生长轮的湿胀应变量介于二者之间,且早材与晚材之间径向湿胀应变量的差值随相对湿度增加而增大。在90%相对湿度条件下,LW的径向湿胀应变量约为EW的1.6倍,ELW-L的径向湿胀应变量约为ELW-E的1.7倍。晚材的径向湿胀应变量明显高于早材主要有2方面原因:一方面是因为相同条件下晚材的平衡含水率高于早材,即吸着点多,细胞壁无定形区能容纳更多的吸着水分子进入,从而引起晚材的湿胀应变量更大(Bertaudetal., 2004); 另一方面可归因于木射线细胞对早材细胞的径向湿胀具有抑制作用(Maetal., 2006; Pateraetal., 2013),由于早材细胞壁薄、刚度低,早材和晚材中木射线细胞的组织比率相近,因此相比于晚材,横向排列的木射线细胞在早材径向湿胀中呈现更显著的抑制作用(Murataetal., 2001; Crameretal., 2005; Pateraetal., 2018)。
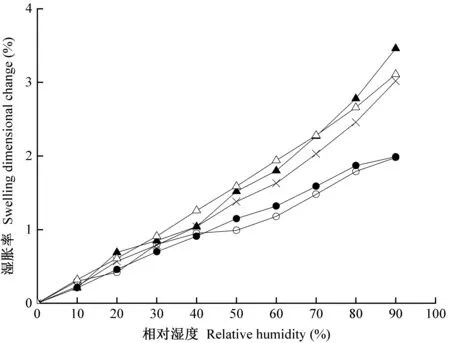
图5 独立早材、独立晚材和生长轮的径向湿胀率Fig. 5 The radial swelling dimensional change of EW, LW and ELW独立早材EW; △独立晚材LW; ●生长轮内早材ELW-E; ▲生长轮内晚材ELW-L; ×生长轮ELW.下同The same below.
从图5还可看出,在相对湿度0%~90%范围内,ELW-E的径向湿胀应变量略高于EW,说明生长轮内晚材发生径向湿胀时带动着生长轮内早材发生径向湿胀。比较ELW-L与LW的径向湿胀应变量,当相对湿度小于70%时,前者略小于后者,说明生长轮内晚材的径向湿胀受生长轮内早材的抑制; 当相对湿度大于70%时,前者反超后者,说明高湿条件下生长轮内早材对生长轮内晚材的径向湿胀有促进作用。此外,ELW的湿胀应变量略低于ELW-L,但明显高于ELW-E,说明楸木生长轮的径向湿胀行为是生长轮内早材和生长轮内晚材发生共同湿胀的净效果,晚材的湿胀行为占主导(Maetal., 2006; Pateraetal., 2018)。
2.3 早材和晚材的弦向湿胀行为
图6所示为独立早材(EW)、独立晚材(LW)、生长轮内早材(ELW-E)、生长轮内晚材(ELW-L)和生长轮(ELW)弦向湿胀应变量随相对湿度的变化情况。可以看出,LW的弦向湿胀应变量大于EW,ELW的弦向湿胀应变量介于二者之间,且LW与EW之间弦向湿胀应变量的差值随相对湿度增加而增大。在90%相对湿度条件下,LW的弦向湿胀应变量约为EW的1.3倍,ELW-L的弦向湿胀应变量与ELW-E相近。这是因为在ELW-E与ELW-L的交界处,存在弦向的拉伸/压缩应力区域,湿胀应变量高的ELW-L带动着湿胀应变量低的ELW-E一起发生湿胀,从而使得二者湿胀应变量基本一致(Maetal., 2006; Pateraetal., 2018)。
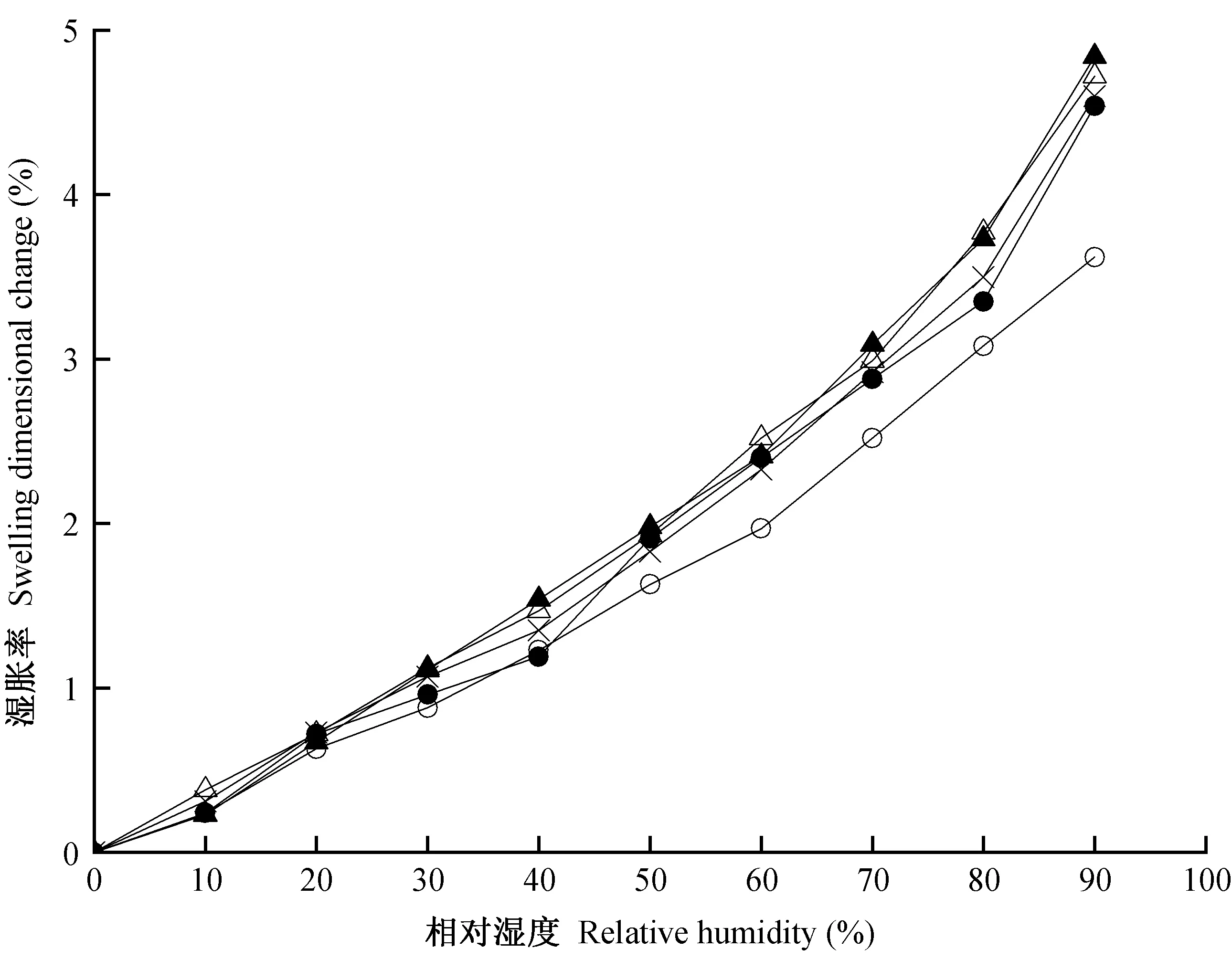
图6 独立早材、独立晚材和生长轮的弦向湿胀率Fig. 6 The tangential swelling dimensional change of EW, LW and ELW
从图6还可看出,在90%相对湿度条件下,ELW-E的弦向湿胀应变量约为EW的1.3倍,EWL-L的弦向湿胀应变量与LW基本一致。这一现象说明ELW-E发生弦向湿胀时,被迫与ELW-L同步,达到与之相适应的湿胀应变量(Rafsanjanietal., 2012a; 2012b)。ELW的弦向湿胀应变量也约为EW的1.3倍,但与ELW-E、ELW-L、LW的弦向湿胀应变量无明显差异。综上可知,楸木生长轮的弦向湿胀行为主要取决于晚材(Lanvermannetal., 2014)。
2.4 早材和晚材的径、弦向差异湿胀
图7所示为独立早材(EW)、独立晚材(LW)、生长轮内早材(ELW-E)、生长轮内晚材(ELW-L)和生长轮(ELW)径、弦向差异湿胀随相对湿度的变化情况。可以看出,一方面,ELW、ELW-L、LW均表现出相似的差异湿胀(约为1.30),且随相对湿度增加稍有增大,这主要是因为随着含水率增加,弦向湿胀的变化速率比径向湿胀更大(Chauhanetal., 2004); 另一方面,ELW-E与EW的差异湿胀取决于相对湿度水平,当相对湿度大于40%时,二者的差异湿胀均明显增大,推测是因为此时细胞壁中开始形成多分子层的吸着水分子,微纤丝之间水层变厚引起细胞壁的径、弦向差异湿胀进一步增大(Pateraetal., 2013); 当相对湿度为90%时,EW的差异湿胀为1.83,ELW-E的差异湿胀为2.28。无论是独立早材还是生长轮内早材,均表现出比晚材更高的径、弦向差异湿胀,分析其原因: 1) 早材细胞壁S1和S3层的厚度百分比高于晚材,且早材细胞壁的径、弦向差异湿胀由S1和S3层主导,而晚材细胞壁则S2层起主导作用(Fengeletal., 1973; Sahlbergetal., 1997),Rafsanjani等(2014)研究发现当S1和S3层比S2层的刚度大时,其径、弦向差异湿胀大; 2) 木射线可抑制壁薄、腔大的早材细胞径向湿胀,而壁厚、腔小的晚材细胞径向湿胀则几乎不受木射线影响(Rafsanjanietal., 2012a); 此外,细胞壁厚薄、排列和形状等也均会对木材径、弦向差异湿胀产生影响(Salmén, 2015)。
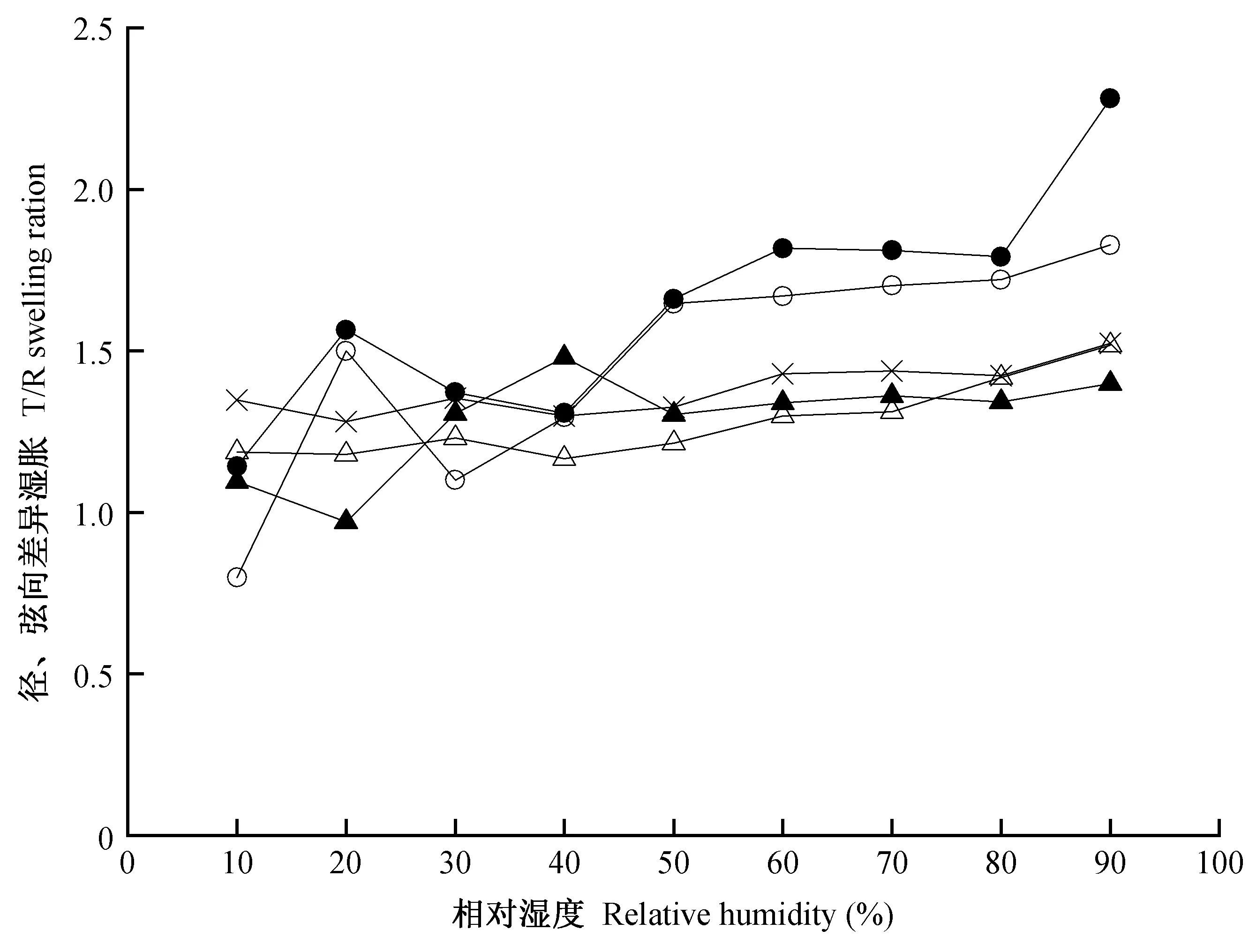
图7 独立早材、独立晚材和生长轮的径、弦向湿胀差异Fig. 7 The ratio of tangential to radial swelling for EW, LW and ELW
相比于独立早材,生长轮内早材的径、弦向差异湿胀更大,推测是因为生长轮内早材的弦向湿胀受晚材影响,被迫达到与生长轮内晚材弦向湿胀应变量一致,使得生长轮内早材的弦向湿胀大于不受外力影响的独立早材,从而表现出更高的径、弦向差异湿胀(Crameretal., 2005)。
2.5 早材和晚材的“含水率平衡态”与“径、弦向湿胀应变平衡态”
表1所示为独立早材(EW)、独立晚材(LW)和生长轮(ELW)在水分吸着阶段结束时(T1)和平衡含水率恒定阶段结束时(T2)的湿胀应变量。可以看出,“保持含水率平衡态360 min”产生的径、弦向湿胀应变量均比“刚达到含水率平衡态”低4个数量级,可忽略不计。这一结果表明,EW、LW与ELW达到“含水率平衡态”的时间点与达到“径、弦向湿胀应变平衡态”的时间点是一致的,没有发现滞后现象,也就是说“细胞壁的尺寸变化”对于“水分进入细胞壁无定形区的响应”是及时的(Thithanhetal., 2015; Joffreetal., 2016)。
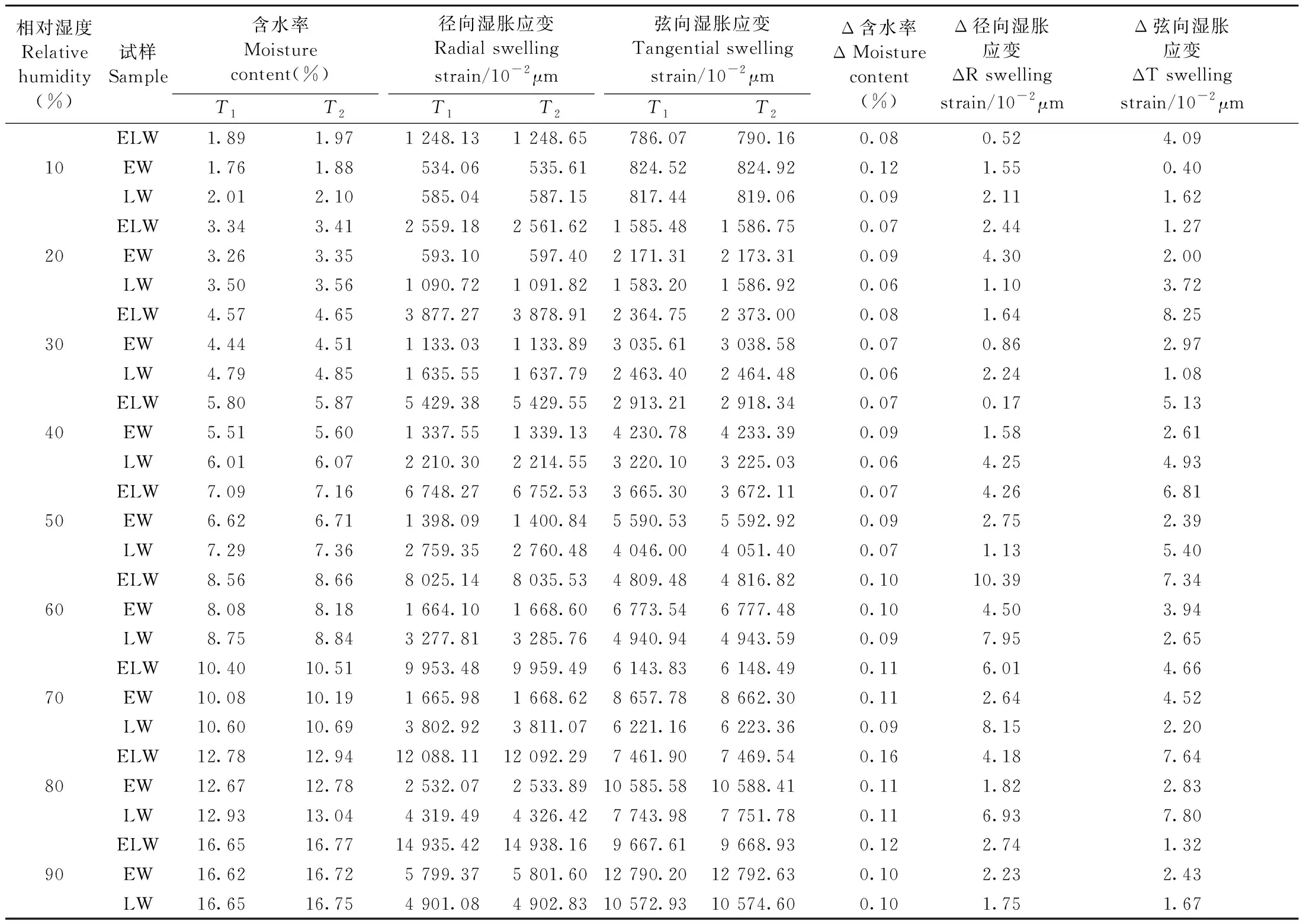
表1 水分吸着阶段(T1)和平衡含水率恒定阶段(T2)的径、弦向湿胀应变与含水率变化①Tab.1 The radial and tangential swelling strain and EMC of the water vapor sorption period(T1)and the EMC constant period(T2)
3 结论
1) 在水分吸着阶段任一相对湿度水平,晚材的平衡含水率均高于早材。当相对湿度小于50%时,二者之间的差值逐渐增大; 当相对湿度大于50%时,二者之间的差值逐渐减小。生长轮平衡含水率介于二者之间。
2) 径向湿胀: 晚材明显大于早材,且主导生长轮的径向湿胀行为; 弦向湿胀: 由湿胀应变量更高的晚材带动早材一起发生弦向湿胀,从而主导生长轮的弦向湿胀行为; 径、弦向差异湿胀: 晚材小于早材,且这一趋势在相对湿度大于40%时更为明显。
3) 在水分吸着阶段,生长轮、独立早材和独立晚材三者的“径、弦向湿胀应变”与 “含水率平衡态”具有同步性,不存在时间滞后现象。
