放牧藏绵羊暖、冷季瘤胃VFAs与 SGLT1基因表达特征关联分析
2021-06-28沙玉柱吕卫兵罗玉柱王继卿李少斌
魏 红,沙玉柱,史 浩,吕卫兵,罗玉柱,王继卿,李少斌,胡 江,刘 秀
(甘肃农业大学 动物科学技术学院/甘肃省草食动物生物技术重点实验室,兰州 730070)
藏绵羊是青藏高原最古老的绵羊品种,与蒙古羊、哈萨克羊并称为中国三大粗毛羊品种,繁育在海拔3 000 m以上的青藏高原地区。藏绵羊对青藏高原的高寒低氧环境具有良好的适应性,不但能够在严酷的自然环境中健康生长和繁育,还能够给当地牧民提供肉、毛、燃料等生产和生活必须品,是当地牧民经济收入的主要来源,也是青藏高原重要的家畜品种和种质资源。全年放牧是藏绵羊千百年来形成的传统,尽管“暖牧冷饲”的饲养模式逐渐被牧民接受,但大多数地区还是以全年放牧为主,采食天然牧草是放牧藏绵羊获取食物的主要来源,在漫长的冷季(10月-翌年4月),藏绵羊只能通过啃食枯草来获取营养物质以满足自身营养需要。面对青藏高原暖、冷季饲草供给存在的严重不均衡,藏绵羊如何应对暖、冷季营养供给的巨大变化及冷季营养胁迫,解析这一科学问题是藏绵羊产业发展的迫切需求。
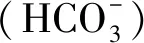
碳水化合物(Carbohydrate,CHOs)可被唾液和胰腺分泌的淀粉酶以及存在于肠上皮刷状缘膜中的寡糖、二糖和三糖酶分解[4],生成的单糖(葡萄糖、半乳糖和果糖)被肠道吸收,为机体提供能量。肠上皮组织的葡萄糖吸收方式有两种,分别是钠依赖葡萄糖共转运体(SGLT1-Sodium glucose cotransporter)进行的主动转运和葡萄糖转运蛋白(Glucose transporters,GLUTs)进行的易化扩散。其中,钠-葡萄糖共转运蛋白(SGLTs-Sodium glucose cotransporters)是小肠上皮细胞上摄取和转运葡萄糖的主要载体,除小肠上皮细胞吸收葡萄糖外,瘤胃上皮细胞也能吸收部分葡萄糖[5]。表明瘤胃内的营养物质不仅被微生物降解为VFAs吸收,还有部分葡萄糖通过SGLT1基因被瘤胃上皮吸收,但暖、冷季藏绵羊瘤胃、小肠、大肠组织SGLT1基因的表达情况未见报道。
综上所述,基于VFAs与SGLT1基因对反刍动物进行能量输送和调控中所发挥的重要作用,本研究以瘤胃上皮SGLT1基因表达与VFAs间的相关性研究为切入点,以期得到它们之间的相互联系。从而为藏绵羊应对暖、冷季营养供给严重不均衡以及冷季营养胁迫提供基础,也为藏绵羊种质资源保护提供参考。
1 材料与方法
1.1 实验动物与样品采集
在甘肃省甘南藏族自治州合作市佐盖曼玛乡(海拔3 300 m)同一牧户羊群选取1周岁(±1月龄)、身体健康、体质量相近(≤2 kg)、自然放牧状态、没有进行任何补饲的藏绵羊母羊12只。分别于7月中旬(2019年7月15日,代表暖季)和12月中旬(2019年12月15日,代表冷季)各采集6只藏绵羊瘤胃液以及组织样。
瘤胃液采集:试验羊只归牧后,在次日早晨测量体质量,利用羊用胃管式瘤胃采样器采集瘤胃液,每只羊采集2管,并将样品立刻放入准备好的液氮罐中进行冷冻保存,带回实验室中进行 -80 ℃保存。
组织样采集:每个时期分别屠宰6只藏绵羊进行肠道内容物及组织样品的采集。采用颈静脉放血屠宰,立即取瘤胃、小肠(十二指肠)、大肠(盲肠)组织样,将其分离出来,再用4 ℃预冷的生理盐水和PBS缓冲液将瘤胃、大肠、小肠的各个部位冲洗干净,用吸水纸把样品吸干,然后再分别取瘤胃、大肠、小肠部位样各2 cm左右,放到 1.5 mL离心管中,然后将样品立刻放入到准备好的液氮罐中进行冷冻保存,带回实验室中进行 -80 ℃保存,以供后续分析。
1.2 VFAs测定
采用日本岛津(GC-2010 plus)气相色谱仪测定藏绵羊瘤胃VFAs,内标为2-乙基丁酸(2EB),色谱柱为AT-FFAP(50 m×0.32 mm×0.25 μm)毛细管柱。色谱柱升温程序:温度60 ℃保持1 min,以5 ℃/min升至115 ℃,不保留,再以15 ℃/min升至180 ℃,检测器温度260 ℃,进样口温度250 ℃。
1.3 总RNA提取和胃肠道组织cDNA合成
利用Trizol一步抽提法提取藏绵羊瘤胃、小肠、大肠组织总RNA,测定总RNA浓度和纯度,琼脂糖凝胶电泳检测RNA质量,全部样品的吸收比(A260/A280)在1.8~2.1。使用含有 gDNA Wiper酶的反转录试剂盒进行cDNA合成,严格按照试剂盒说明书进行操作。
1.4 设计与合成引物
从NCBI中搜索出绵羊SGLT1基因的 mRNA序列(GenBank No:NM_000343.4),利用Premier 5.0设计SGLT1基因扩增引物,引物信息(表1)。

表1 引物信息Table 1 Primer information
1.5 SGLT1基因Real-time PCR扩增
以β-Actin为内参基因,利用实时荧光定量PCR仪(QuantStudioTM6 Flex,ABI,美国)进行SGLT1基因及内参基因荧光定量,反应条件: 95 ℃预变性30 s;循环反应95 ℃ 10 s,60 ℃ 30 s,40个循环;每次循环的最后一步收集SYBR green荧光信号,溶解曲线(95 ℃ 15s,60 ℃ 60 s,95 ℃ 15s)。反应体系:20 μL体系,含有2×ChamQ Universal SYBR qPCR Master Mix, cDNA模板和上下游引物。以β-Actin为内参基因进行校正。
1.6 数据分析
数据均采用SPSS 22.0统计软件处理,以“平均值±标准差”形式表示。VFAs含量进行独立样本t检验,SGLT1基因表达量,以β-Actin为内参基因,采用2-ΔΔCT的方法计算相对表达量。瘤胃VFAs与SGLT1基因组织表达进行Spearman相关性分析。
2 结果与分析
2.1 藏绵羊暖、冷季瘤胃VFAs测定结果
藏绵羊暖、冷季瘤胃VFAs测定结果显示(表2),藏绵羊瘤胃在冷季的总VFA含量显著高于暖季(P<0.05),除丁酸外,其他VFA含量在暖、冷季均差异显著(P<0.05)。冷季的乙酸、丙酸含量均显著高于暖季(P<0.05),异丁酸、异戊酸、戊酸的含量为暖季显著高于冷季(P<0.05),丁酸在暖、冷季差异不显著(P>0.05)。另外,冷季乙酸/丙酸(A/P)值显著高于暖季(P<0.05)。
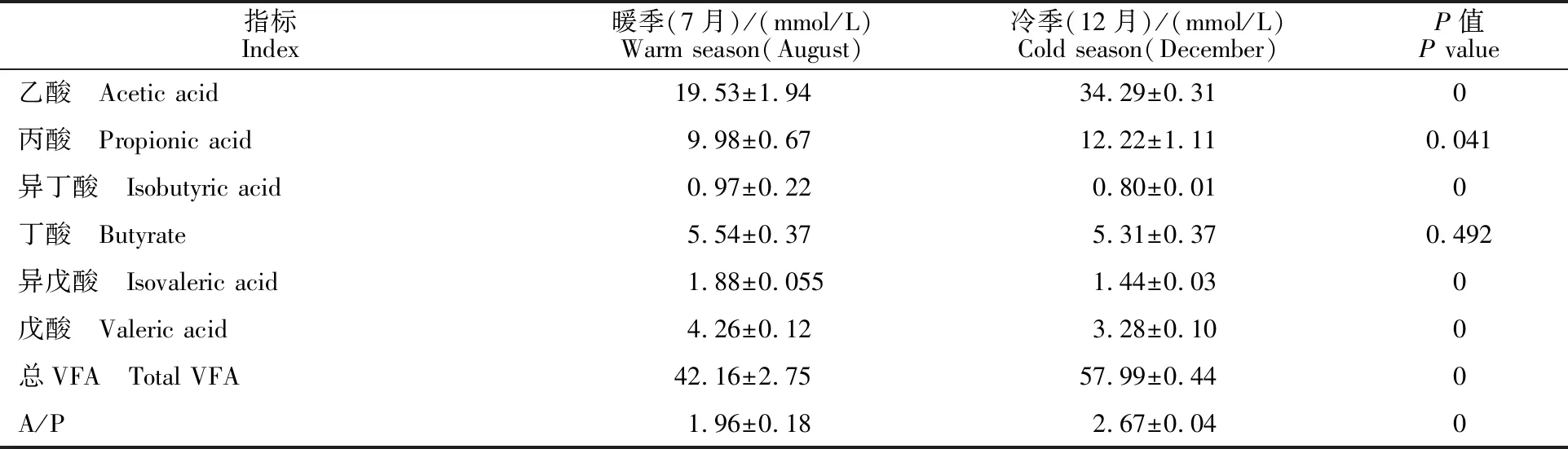
表2 藏绵羊暖、冷季瘤胃VFAs含量测定结果Table 2 Determination of VFAs concentration in rumen of Tibetan sheep in warm and cold seasons
2.2 藏绵羊暖、冷季胃肠道 SGLT1基因表达特征
通过对藏绵羊暖、冷季胃肠道组织(瘤胃、小肠、大肠)SGLT1基因进行实时荧光定量检测发现(图1),藏绵羊暖、冷季胃肠道组织的SGLT1基因表达存在显著性差异(P<0.05)。其中,暖季的瘤胃、小肠、大肠组织的SGLT1基因表达量显著高于冷季(P<0.05),同一季节不同肠道组织的SGLT1基因表达也存在一定差异,表现为:小肠组织>瘤胃组织>大肠组织。藏绵羊小肠组织的SGLT1基因表达量显著高于瘤胃和大肠组织(P<0.05),在暖、冷季无差异。其中,暖季比冷季小肠组织的表达量高出1.8倍。
2.3 藏绵羊瘤胃VFAs与 SGLT1基因之间相关性分析
藏绵羊瘤胃VFAs和SGLT1基因之间存在一定的相关性(表3),其中,乙酸、丙酸与SGLT1基因呈显著性负相关(P<0.05),相关性系数均大于0.9;异丁酸、异戊酸、戊酸与SGLT1基因呈显著性正相关(P<0.05),相关性系数均在0.98以上。另外,6种VFA之间也存在一定的相关性,除丁酸外,其他VFA之间的相关性均差异显著(P<0.05)。
3 讨 论
3.1 藏绵羊暖、冷季瘤胃VFAs含量分布特征
VFAs既是碳水化合物在瘤胃中的终产物,也是瘤胃微生物生长所需的重要碳架和能量来源,能够维持正常的瘤胃内环境[6]。瘤胃VFAs含量和组成受日粮、饲料加工处理、瘤胃微生物、瘤胃PH等因素的影响[7]。在本研究中,藏绵羊在冷季的VFAs含量显著高于暖季(P<0.05),其中,冷季的乙酸和丙酸的含量显著高于暖季 (P<0.05),可能是由于暖季饲草料丰富、营养价值高,为瘤胃微生物生长提供了充足养分,为产甲烷菌生长增殖提供了充足氮源。瘤胃内纤维分解菌生长增殖快,纤维分解效果大大提高,为产甲烷菌的生长增殖提供了充足的氢源,产甲烷菌与产乙酸菌竞争氢生成甲烷或乙酸[8]。反刍动物主要的排氢途径是在瘤胃内生成甲烷。因此,暖季瘤胃内甲烷含量高,导致乙酸含量降低。Fonty等[9]研究发现,有产甲烷菌的羔羊氢利用能力高达90%以上,无产甲烷菌时羔羊的氢利用能力降低,仅为28%~46%。在正常生长的羔羊瘤胃发酵过程中,乙酸的产生仅占一小部分,但在无产甲烷菌时则占瘤胃发酵的21%~25%[10]。由于冷季饲草养分质量低,产甲烷菌不能获得足够的养分,限制产甲烷菌的生长繁殖,因而冷季产甲烷菌数量明显低于其他季节[11],以致冷季的乙酸含量高于暖季。另外,异丁酸、异戊酸、戊酸的含量为暖季显著高于冷季(P<0.05),主要是因为VFAs中异丁酸和异戊酸是由蛋白质分解产生的[12]。藏绵羊在暖季可以采食能量和蛋白质含量较高的牧草,瘤胃中可利用碳水化合物充足,氨利用率高,蛋白质合成强度大,从而导致异丁酸、异戊酸等暖季含量大于冷季。研究发现,不管是冷季还是暖季,乙酸的含量都较高,约占VFA总产量的70%~75%[13],主要是因为乙酸是反刍动物新陈代谢的主要能量来源。同时,乙酸通过葡萄糖经磷酸戊糖途径代谢和葡萄糖存在依赖关系[14]。丙酸是唯一生成葡萄糖的VFA,是肝脏组织中糖异生的重要葡萄糖前体,在经瘤网胃壁吸收过程中为肌肉组织提供能量[15]。冷季的A/P值显著高于暖季(P<0.05),且A/P值均在 1~3,从能量利用的角度看,A/P比值越低,对饲料能量利用效率越高,放牧藏绵羊冷季的A/P值高于暖季,说明放牧藏绵羊对其牧草的利用效率较高。综上所述,藏绵羊为应对冷季营养物质严重匮乏,进化出了较强的能量适应机制。
3.2 藏绵羊暖、冷季胃肠道 SGLT1基因表达 特征
致使暖季SGLT1基因表达量远高于冷季的原因较多[16-17],如季节、日粮、饥饿状况、Na+摄入水平、激素水平、炎症、应激、肠黏膜上皮细胞沿肠绒毛轴的分化及其细胞周期等[18]。本研究中,小肠组织中的SGLT1基因表达量均显著高于瘤胃和大肠组织(P<0.05),小肠上皮细胞上摄取和转运葡萄糖的主要载体是SGLTs,介导肾脏和肠道中葡萄糖的跨膜转运,所以小肠的SGLT1基因表达量较高。藏绵羊暖季瘤胃、小肠、大肠组织的SGLT1基因表达量显著高于冷季(P< 0.05),这可能与全年放牧有关,由于冷季饲草匮乏,个体之间存在更激烈的觅食竞争,导致个体在获取饲料资源方面存在更大的差异。随着暖季食物可得性的增加,个体采食竞争减小、饲草料丰富且营养价值较高[19],以及暖季牧草中的含糖量显著高于冷季[20-21]。景小平等[22]研究发现,当胃肠道里面的葡萄糖含量慢慢增加时,SGLT1基因也在增加,进而可以让胃肠道对葡萄糖的吸收效率得到较大的提高,且葡萄糖含量越高,SGLT1基因的活性和效率越高,所以,暖季胃肠道SGLT1基因表达量高于冷季。另外,卢垚等[18]的研究发现,在漫长的冷季,由于寒冷的气候,藏绵羊发生炎症的几率增大,受到的应激变大,所以冷季SGLT1基因表达量较低。王修启等[23]、伦志国[24]研究表明,由于过瘤胃葡萄糖的作用,导致瘤胃中的葡萄糖以某种方式保护起来,使其在反刍家畜瘤胃内不易被降解,而直接进入瘤胃后消化系统(主要是小肠)进行消化、吸收和利用,所以导致小肠的SGLT1基因表达量最高。但本研究主要针对暖、冷季的瘤胃SGLT1基因的表达特征进行分析,虽说瘤胃中大部分葡萄糖直接进入后消化系统进行消化吸收或者直接被发酵为VFAs,但是,在暖季牧草量充足的情况下,瘤胃组织直接将多余的葡萄糖通过SGLT1蛋白转运进入血液,节约了发酵所需的能量[25]。此外,暖季瘤胃SGLT1基因表达量显著高于大肠组织(P<0.05)。瘤胃中的SGLT1基因吸收葡萄糖,降低反刍动物前胃葡萄糖浓度,可改善或预防反刍动物乳酸酸中毒。有研究认为[26],反刍动物对碳水化合物的消化吸收主要是由其在瘤胃中转化为VFAs的转化效率来决定的,赵小刚等[27]研究证实,瘤胃上皮SGLT1基因的转录和功能性表达水平对反刍动物利用葡萄糖也具有很大的影响。因此,对研究胃肠道葡萄糖稳态的调控具有重要意义。
3.3 藏绵羊暖、冷季瘤胃VFAs与 SGLT1基因之间的相关性
进一步评估VFAs与SGLT1基因表达之间的相关性,可以发现乙酸、丙酸与SGLT1基因呈显著性负相关(P<0.05),相关性系数均大于 0.9,异丁酸、丁酸、异戊酸、戊酸与瘤胃上皮的SGLT1基因之间呈显著性正相关(P<0.05)。如前所述,在反刍动物中,日粮的碳水化合物发酵成VFAs主要由瘤胃微生物发酵,通过瘤胃上皮吸收,在肝脏中被用于糖异生来为动物提供能量,部分瘤胃没有发酵消化的非结构性碳水化合物、葡萄糖及与微生物相关的蛋白质,可以经SGLT1基因介导直接被瘤胃上皮吸收进入血液中,在血液中供机体使用,剩余的蛋白质被小肠再次吸收,在小肠消化酶的作用下,进一步变为小肽、氨基酸和单糖转运载体[28]。因此,到达小肠的单糖量减少,随着到达小肠的单糖含量降低,小肠的SGLT1基因含量和mRNA水平均显著降低,表明VFAs与SGLT1基因之间存在着一定的关联。有研究表明[5],营养物质不仅仅以VFAs的形式被瘤胃上皮吸收,还有部分葡萄糖直接被瘤胃上皮SGLT1基因介导吸收进入血液;此外,反刍动物体内葡萄糖相对于VFAs的能量值远高于单胃动物,这是因为反刍动物几乎完全依赖葡萄糖糖异生来满足其对葡萄糖的需求[29]。通过SGLT1基因直接摄取葡萄糖可以减少动物糖异生的代谢消耗[30]。在漫长的冷季,饲草料严重匮乏,藏绵羊通过高产量的VFAs以维持自身新陈代谢和提供能量。并且在SGLT1基因的作用下,有效调节了胃肠道内葡萄糖含量的波动。因此,探讨藏绵羊暖、冷季瘤胃VFAs与SGLT1基因的相互调控作用,从而为藏绵羊应对暖、冷季营养供给严重不均衡以及冷季营养胁迫提供基础,也为藏绵羊种质资源保护提供 参考。
4 结 论
放牧藏绵羊瘤胃中的VFAs含量在冷季显著高于暖季(P<0.05)。除丁酸外,其他VFA在暖、冷季均存在显著性差异(P<0.05)。对放牧藏绵羊瘤胃、大肠、小肠SGLT1基因的表达量测定发现,藏绵羊暖季的瘤胃、大肠和小肠SGLT1基因的表达量显著高于冷季(P<0.05),在暖、冷相同的季节中小肠组织的SGLT1的基因表达量最高。对藏绵羊瘤胃VFAs与SGLT1基因表达间的相关性分析发现,乙酸、丙酸与SGLT1基因表达呈显著性负相关(P<0.05),异丁酸、丁酸、异戊酸、戊酸与瘤胃上皮的SGLT1基因之间呈显著性正相关(P<0.05)。
