依普黄酮对骨质疏松大鼠颌骨成骨细胞的线粒体相关因子的影响
2021-06-27于书娟刘洪臣
于书娟 刘洪臣
由于颌骨作为全身骨骼的一部分,在很多方面是相似的,有研究表明,全身骨的骨量减少与颌骨的骨量减少非常相似,并且有一定的相关性[1-3],所以很多学者在颌骨的骨量减少的防治上使用全身骨质疏松症的治疗药物。依普黄酮(ipriflavone,IP)是人工合成的异黄酮,已被报道能增加骨质疏松患者的骨密度[4]并具有抗氧化特性[5],其虽然具有雌激素的作用,但对子宫内膜和乳腺的刺激作用小于雌激素[6,7],作为骨质疏松症药物在临床已经应用多年,并且引起口腔医学者的关注。有学者研究发现依普黄酮可以促进牙周膜细胞的体外增殖和成骨分化,促进牙齿移动后的牙周组织的重组,预防正畸后的复发[8]。也有学者的研究结果提示依普黄酮可以增加颌骨的骨密度[9],从而对颌骨的骨质疏松有一定的防治效果。本实验通过研究依普黄酮对骨质疏松大鼠颌骨成骨细胞的线粒体相关因子解偶联蛋白2(uncoupling protein 2,UCP2)、腺苷三磷酸(adenosine triphosphate,ATP)和活性氧(reactive oxygen species,ROS)的影响,为依普黄酮应用于口腔颌骨骨质疏松防治的研究提供理论基础,并且初步探索其分子作用机制。
1.材料和方法
1.1 主要试剂和设备 依普黄酮(Sigma,美国),杜尔贝科改良伊格尔培养基(dulbecco′s modified eagle medium,DMEM)(Gibco,美国),胰蛋白酶(Amresco,美国),新生牛血清(四季青,杭州),胎牛血清(Gibco,美国),培养细胞总RNA 提取试剂盒(TIANGEN,北京),逆转录聚合酶链式反应试剂盒(TIANGEN,北京),SDS-PAGE 凝胶制备试剂盒(BOSTER,武汉),总蛋白抽提试剂盒(BOSTER,武汉),辣根酶标记的山羊抗兔IgG(Santa,美国),兔抗鼠UCP2 单克隆抗体(BOSTER,武汉)。其余试剂为国产分析纯。
双能X 线骨密度仪(Lunar,美国),ELx800uv全自动酶标仪(Bio-Tek,美国),倒置相差显微镜(Leica,德国),紫外分光光度计(Beckman-DU640,美国),多功能荧光分析仪(FLUOstar Omega,BMG LABTECH,德国),TDZS-WS 型多管架自动平衡高速离心机(湘仪公司,长沙),低温高速离心机(Eppendorf,德国),ST-Ⅰ型半干式转移电泳槽(竞迈生物,大连)。
1.2 动物来源20 只SD 大鼠(雌性,12 周龄,约300g)在实验动物房中进行适应性喂养,观察一周,大鼠均健康。
1.3 骨质疏松大鼠动物模型的建立 实验大鼠被随机平均分为两组[去势组(n=10)和假手术组(n=10)]。麻醉后取仰卧位,正中切开腹部,去势组对双侧卵巢结扎切除,假手术组对双侧卵巢附近的与卵巢大小相似的小块脂肪切除,严密缝合腹部伤口,术中注意无菌操作,术后注射抗生素防止感染。大鼠在术后12 周时,通过综合分析评价,成功建立骨质疏松动物模型[10]。
1.4 骨质疏松大鼠颌骨成骨细胞的培养和传代 全麻下处死已建立骨质疏松模型的大鼠,无菌条件下取出下颌骨,去除颌骨上的肌肉和筋膜,完整的拔除颌骨上的牙齿,运用课题组人员熟练使用的成骨细胞培养方法,即组织块法和酶消法共同培养的方法[10,11]。首先,在无菌器皿中用无菌剪刀将下颌骨,剪成约2mm 直径大小的骨片,倒入0.125%胰蛋白酶将骨片完全覆盖,约8min 后,将首次消化的液体倒掉。再用胰蛋白酶连续消化骨片5次,依次收集,高速离心,将离心管中上清液弃去,管底部细胞重悬后接种到培养瓶中。然后胰蛋白酶消化后留下的骨片加入培养基,继续用无菌剪刀将骨片剪成约1mm 直径大小,倒入培养瓶中培养。待成骨细胞长满全瓶后传代。
当培养的成骨细胞在培养瓶中达到约80%的融合率时,换成不含血清的培养基继续培养24h,以使得细胞达到同步化,然后将细胞分成依普黄酮组和对照组,分别采用含10-8M 依普黄酮[12]的培养基和不含依普黄酮的培养基培养成骨细胞72h。然后进行ATP含量检测、ROS含量检测、UCP2的RT-PCR基因检测和UCP2 的western blot 蛋白检测。
1.5 成骨细胞的ATP 含量检测 将成骨细胞以3×105细胞/ml 接种于5个6 孔板,用无血清培养基同步化,每个6 孔板中3个孔使用药物干预,3个孔用作对照。药物干预72h 后。使用ATP 检测试剂盒(Beyotime,中国)中提供的试剂,溶解成骨细胞10min。溶解后,混合物在12000g 和4oC下离心10min。将ATP 检测工作液在不透明的96孔板中孵育5min,以减少ATP 的背景。然后将离心后的成骨细胞上清液转移到96 孔板中2s,立即用多功能荧光分析仪(FLUOstar Omega,BMG LABTECH,德国)在562nm 波长下测量化学发光(荧光素酶催化荧光素产生荧光)。根据标准曲线计算出ATP 浓度。
1.6 成骨细胞的ROS 含量检测 将成骨细胞以1×106细胞/ml 的密度接种于5个6 孔板,每个6 孔板中3个孔使用药物干预,3个孔用作对照。细胞同步化和药物干预培养后,用PBS 洗涤细胞两次,并将用无血清培养基稀释的DCFH-DA 探针(1∶1000 稀释)添加到培养板中,继续培养细胞20min 后,用无血清培养基连续洗涤3次,用于去除多余的DCFH-DA,然后以800 转/分离心6min。用PBS 洗涤细胞并在0.3ml PBS 中重新悬浮,将0.2ml 含有细胞的悬浮液转移到96 孔板中,用多功能荧光光分析仪(FLUOstar Omega,BMG LABTECH,德国)测量荧光(488nm 的激发/波长,525nm 的散发波长)。通过减去自身荧光背景校正,最终获得成骨细胞的ROS 含量。
1.7 RT-PCR 将成骨细胞以1×105细胞/ml接种于3个12 孔板,每个12 孔板中6个孔使用药物干预,6个孔用作对照。培养72h 后,用PBS 连续冲洗三次,用总RNA 提取试剂盒抽提成骨细胞的总RNA。用逆转录聚合酶链反应式试剂盒反转录1.5μg 的mRNA 为cDNA,进行PCR。引物的合成通过专业生物技术公司(上海英俊公司,中国)完成,见表1。
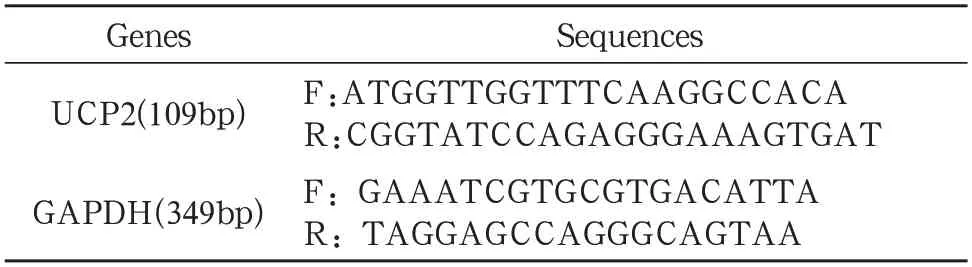
表1 用于RT⁃PCR的基因引物
每个25μl 的PCR 反应体系包括1.5μl 的cDNA,0.5μl 的dNTPs(10mM),2.5μl 的10×PCR 缓冲液,0.5μl 的β-肌动蛋白f(25pmol/ul),0.5μl 的β-肌动蛋白r(25pmol/ul),0.5μl的Taq 酶(2u/ul)和ddH2O(补足至25μl)。设置循环参数,在94℃预变性2min,再按94℃/30s、63℃/30s 和72℃/30s 扩增30个循环。然后用琼脂糖凝胶电泳30min,获得图像,用Labworks 4.0 图像采集分析软件(PerkinElmer,Waltham,Massachusetts,USA)对各条带积分光密度值(IOD)进行分析,以目标基因与GADPH 基因的IOD 值之比作为目标基因表达水平,每一个样品分析三次。
1.8 Western Blotting 将成骨细胞以1×105细胞/ml接种于3个12孔板,每个12孔板中6个孔使用药物干预,6个孔用作对照。药物干预培养72h后,将待检测的成骨细胞用PBS洗涤三次后加入细胞裂解液,冰上轻摇晃15min。刮取各孔的成骨细胞并且收集于1.5ml试管中,在12000g下4℃离心15min,上清液经标准化后分装。含有等量蛋白质(30μg)的成骨细胞上清液,用、SDS-PAGE 分离蛋白,电转移至PVDF膜。转移后,用40ml的0.1%Tris缓冲盐水吐温(Tris-Buffered Saline Tween-20,TBST)洗涤缓冲液(含5%脱脂乳,pH值7.4)并在4℃下过夜封闭,加入稀释度1∶300的兔抗鼠UCP2抗体(BOSTER,武汉),稀释度1∶2000的β-肌动蛋白,在37℃下孵育2h后,洗涤膜三次后,加入稀释度1∶2000的辣根酶标记的山羊抗兔IgG(Santa,美国),继续37℃下孵育1h,滴加化学发光试剂,X光片盒中曝光。通过Quantity One软件分析得出结果。以β-肌动蛋白的百分比表示成骨细胞的UCP2蛋白表达。
1.9 统计学方法 采用统计软件SPSS 25.0进行实验数据的统计分析,数据结果采用平均值±标准差的方式表示,对于依普黄酮组和对照组之间数据的比较采用t检验,P<0.05认为两组间有显著性差异。
2.结果
2.1 倒置显微镜下成骨细胞的形态观察 骨质疏松大鼠颌骨成骨细胞的形态如图1 所示,多呈长梭形、三角形或多角形等多种形态,有多个长或短突起,与相邻的细胞连接;细胞核呈圆形或椭圆形,居于细胞中间或略偏向一侧,可看到明显的一个或两个核仁;在细胞重叠生长的区域,可见紧靠的多个细胞核。
图1 骨质疏松大鼠颌骨成骨细胞(×200)
2.2 成骨细胞ATP 含量检测的结果 骨质疏松大鼠颌骨成骨细胞ATP 含量的荧光检测结果如图2 所示,依普黄酮组(15.3±1.31μmol/g)明显高于对照组(9.9±0.78μmol/g),并且这种差别有统计学意义(P<0.01)。

图2 成骨细胞ATP水平的检测结果(*代表P<0.05)
2.3 成骨细胞的ROS含量检测结果 检测实验组和对照组成骨细胞ROS 含量的结果显示(如图3),依普黄酮组(276±20)明显低于对照组(427±35),有高度显著性差异(P<0.01)。
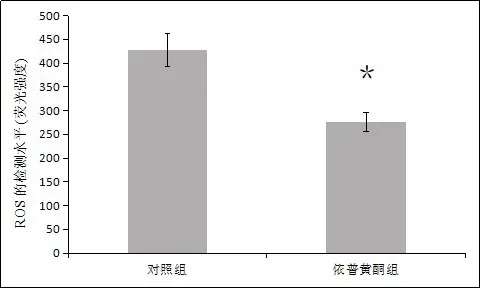
图3 成骨细胞ROS含量的检测结果(*代表P<0.05)
2.4 成骨细胞UCP2 的mRNA 基因表达水平图4 显示的是成骨细胞UCP2 的mRNA 基因表达情况,与对照组(0.68±0.05)相比,依普黄酮组(0.84±0.08)出现了显著的升高,P<0.05。
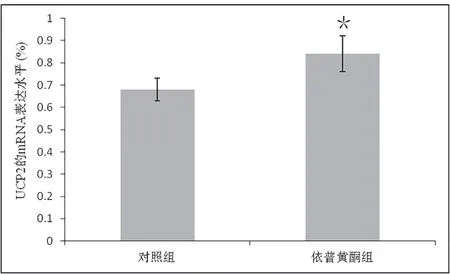
图4 成骨细胞UCP2的mRNA的基因表达水平(*代表P<0.05)
2.5 成骨细胞UCP2 的蛋白表达水平UCP2蛋白检测结果显示,依普黄酮组较高(0.51±0.04),对照组较低(0.39±0.03),两组之间的差异具有统计学上的意义(P<0.05),见图5。
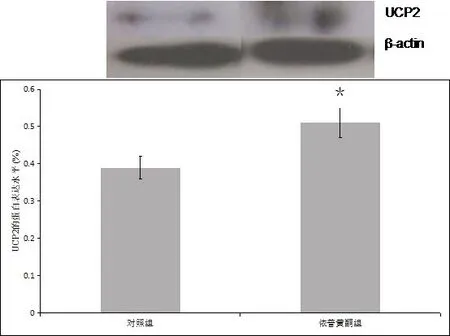
图5 成骨细胞UCP2蛋白水平表达的检测结果(*代表P<0.05)
3.讨论
近年来,人们越来越关注骨质疏松症对口腔医学的影响,如种植体植入后的骨整合,拔牙后牙槽骨的骨吸收,或牙周炎的骨改变等[13-16]。依普黄酮作为一种有效的防治骨质疏松的药物,在多个国家得到了广泛的应用。其不但可以抑制骨吸收,并且具有雌激素和降钙素的作用。依普黄酮可能在GPER1 的p38MAPK 信号通路途径中发挥重要的作用[17]。因此,依普黄酮对颌骨以及口腔疾病的影响也越来越受到学者的关注,对于其相关的分子机制研究也成为热点。有研究表明依普黄酮可以增加牙周膜干细胞的成骨分化,并且GPR30 介导的PI3K/Akt 通路在其中起着重要的作用[8]。p38MAPK 通路和PI3K/Akt 通路均能通过磷酸化某些因子调控着细胞的增殖和凋亡。而磷酸化的过程是在线粒体中完成的。在本研究中,通过体外培养骨质疏松大鼠下颌骨成骨细胞,研究依普黄酮对成骨细胞的线粒体相关细胞因子的影响,也为口腔医学的进一步研究提供了一定的研究思路。
线粒体是一种重要的细胞器,为细胞功能活动提供能量,有“电力站”之称,同时,它也是有氧呼吸的重要场所,与ATP 和ROS 的生成直接相关。而UCP2 作为线粒体内膜上的质子转运蛋白,使质子通过直接渗漏回线粒体基质的方式,部分解偶联氧化磷酸化,使得ATP的产生出现减少的情况[18,19];同时,UCP2 通过减少呼吸链电子漏的发生和降低线粒体内膜两侧的电化学梯度,能减少ROS 的产生。对于ATP 来说,Megha Rajendran 等认为,ATP 不仅是细胞进行各种功能活动能量的直接来源,也是生物体内外新陈代谢和信号传导的中心分子[20]。Lukas Jaroslaw 等研究报道UCP2 通过调控ATP 的浓度起到调控细胞钙的摄取[21];而钙离子在细胞行使功能活动时起到重要的作用。同时,对于ROS 来说,ROS 是衰老、炎症及某些疾病的发病机制之一,可以通过氧化应激反应诱导细胞凋亡,从而起到调节机体细胞生死平衡的作用。已有多个研究表明,当线粒体内膜电势升高时,UCP2可抑制ROS 的产生[22,23]。而在口腔医学研究中,也有学者研究发现炎症微环境可以显著地提高牙周膜干细胞的ROS表达水平,从而起到抑制细胞线粒体生成和抗氧化反应[24]。Fatokun AA 等[25]的研究也表明,ROS 起到影响成骨样细胞作用,从而推测其可能与骨质疏松症有一定得关系。所以说,线粒体相关因子UCP2、ATP和ROS三者既关系密切,又各自在细胞的各种功能活动中起着重要的作用,从而为依普黄酮对骨质疏松症颌骨成骨细胞影响的分子机制的探讨,提供一种好的研究思路和途径。
已经有学者研究发现,去势后大鼠脂肪组织中UCP2的表达出现了降低[26]。并且我们前期的研究结果显示,骨质疏松大鼠下颌骨成骨细胞表达UCP2 较低,而ROS 和ATP 较高[10]。在本实验中,研究发现依普黄酮能促进UCP2 的表达,降低ROS的表达,部分纠正了骨质疏松大鼠颌骨成骨细胞的异常表达,通过降低ROS对成骨细胞的结构和功能具有保护作用,也提示依普黄酮可能通过UCP2 和ROS 来调节骨质疏松大鼠颌骨成骨细胞的功能活动。然而,本实验还表明,依普黄酮对成骨细胞的作用,除了ROS 的减少和UCP2 的增加,ATP 的产生也增加,这似乎是使得骨质疏松大鼠颌骨成骨细胞本来就很高的ATP 的表达更高了,但是,其一方面表明ATP的产生可能不仅仅受UCP2的调控,其他基因或蛋白质也可能对ATP 的产生起调控作用,另一个方面也说明依普黄酮通过增加ATP 水平为成骨细胞的各种功能活动提供足够的能量支持,促进骨生成,从而起到一定的防治骨质疏松的作用。
总之,依普黄酮可能起到调控骨质疏松大鼠颌骨成骨细胞的线粒体相关因子UCP2、ROS 和ATP等的作用,但是其作用机制的研究非常复杂,我们的研究结果只是提供了一定的研究基础和理论依据,仍然需要进一步开展更多的实验研究来探索。
