中国Ceratocystis和Huntiella真菌新种汇录:2014—2020
2021-06-15刘菲菲陈帅飞
刘菲菲,陈帅飞
中国和真菌新种汇录:2014—2020
刘菲菲,陈帅飞
(国家林业和草原局桉树研究开发中心,广东 湛江 524022)
长喙壳属真菌()包含很多重要的树木病原菌,该属真菌在全球广泛分布并在多种寄主中被报道。然而,对于这类真菌在中国树木中的分布、物种多样性和影响的研究较少。近几年从中国南方五省区的多种寄主中分离到了大量的长喙壳属()真菌以及与其亲缘关系相近的亨特长喙壳属()真菌。所采集的寄主包括相思树(spp.)、杉木()、桉树(spp.)和芋头()。本文对2014—2020年期间在我国分离并描述的12个和真菌新种的分类地位、形态特征、菌落特点、寄主范围、分布地域和危害特点等进行了概述,拓展了对长喙霉科真菌在我国的寄主分布以及致病性的了解。所涉及真菌新种包括分离自桉树的,分离自杉木的,分离自芋头的,分离自相思树的,分离自桉树的,,,,,和。
林木病害;块茎作物病害;人工林;桉树;真菌致病菌
长喙壳属()真菌属于子囊菌门(Ascomycota),粪壳菌纲(Sordariomycetes),囊菌目(Microascales),长喙壳科(Ceratocystidaceae)。最早由Halsted(1890)描述,在感染黑腐病(black rot)的美国甘薯()上分离获得。属真菌具有深色的亚球形到球形的子囊,子囊孢子聚集于其长喙的顶端,很容易粘在昆虫上并传播到其他地域或寄主。属真菌自1890年被Halsted描述之后,其分类历史在很长一段时间内都处于有争议的状态。由于具有类似的形态学特征、比较一致的生物学特征以及共享类似的生态环境,长喙壳属()真菌的分类地位一直与长喙科菌目Ophiostomatales中某些属(spp.,spp.,spp.)的真菌混淆。直到近年来多基因序列的系统发育分析方法的应用,囊菌目和长喙科菌目这两大类真菌物种分类难题才得以解决,而最早进行系统发育分析的报道表明这两大类真菌其实分属于子囊菌的不同目,从而使长喙壳类真菌的鉴定和分类取得了重大突破和进展。
21世纪以来,随着越来越多的基于多基因序列系统发育分析、形态学和生态学的物种分类学研究的应用,长喙壳科(Ceratocystidaceae)真菌的分类急需进行系统的重新修订。因此,DE BEER等在2014年对该科物种进行了重修订工作。这项研究针对该科79个已经描述的物种,通过严格的三段多基因序列(60S,LSU,MCM7)将该科重新划分为七个属,这七个属包括,,,,(以前是),(以前是)和(以前是)。在随后的几年里,该科又陆续增加了另外8个属,分别为,,,,,,和。所以,目前长喙壳科(Ceratocystidaceae)真菌已经达到了15个属。
在DE BEER等重修订长喙壳科(Ceratocystidaceae)物种之前,与属于同一个属的真菌,但是在重修订工作以后,被划分成一个独立的属。属真菌的模式物种是,最早从美国得克萨斯州的枫香树()上分离得到。最初被描述为,后来被修订到。spp.可以根据其独特的形态学特征区别于和长喙壳科的其他属。属物种的典型形态学特征为子囊壳表面通常具有伸出的刺,子囊壳颈的基部具有一个垫子。此外,与大多数属物种不同,在物种中很少发现厚垣孢子。
如今,随着分子生物学鉴定方法越来越普及,越来越多的长喙壳科(Ceratocystidaceae)真菌新物种被发现。到目前为止,属真菌已经有40个物种被报道,其中3个来自中国。而属的物种也达到了31个,其中10个来自于中国(表1)。本文对2014—2020年期间在我国首次分离和鉴定出的和属的真菌物种的分类地位、形态特征、菌落特点、寄主范围、分布地域和危害特点等进行概述,以期对桉树真菌病原物种的多样性和特征有更加深入的了解。
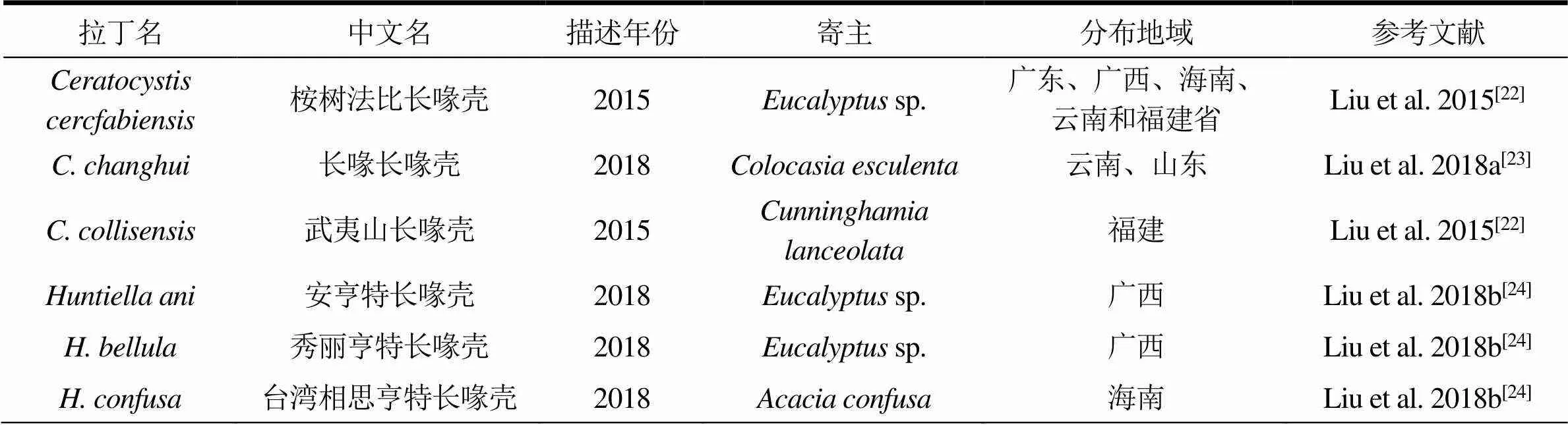
表1 我国Ceratocystis和Huntiella真菌物种简介
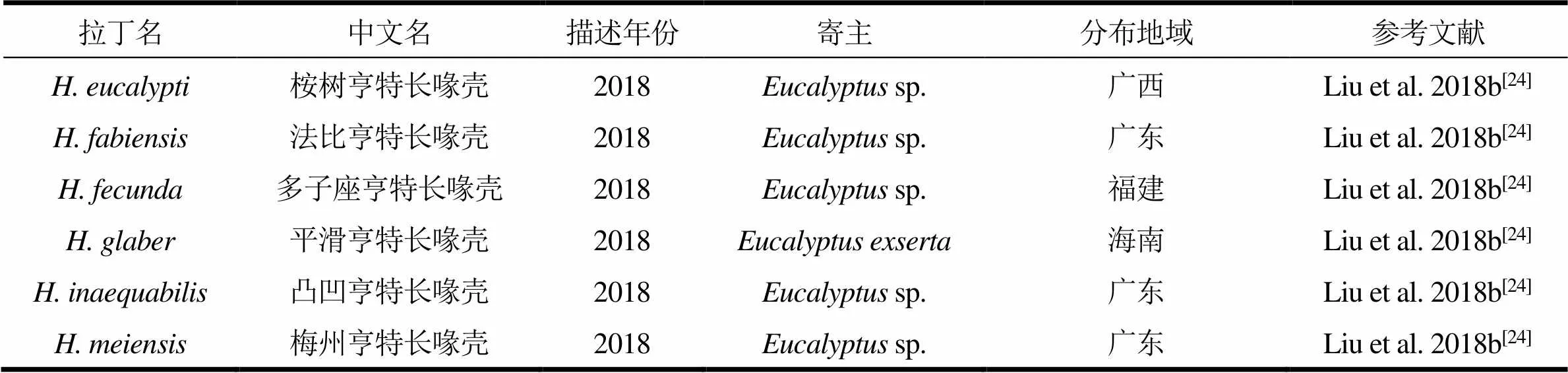
续表1
1 我国Ceratocystis和Huntiella真菌2014—2020年新描述物种简介
1.1 Ceratocystis cercfabiensis F. F. Liu, Jol. Roux & S. F. Chen sp. nov. [22]
MycoBank号码:MB811888
病原中文名称:桉树法比长喙壳
形态特征:
(1)有性阶段:子囊壳基部暗褐色至黑色,球形至近球形,基部宽(79−)138–231(−286) μm,高(100−)137–218.8(−302) μm,表面附着菌丝;子囊壳茎暗褐色至黑色,至茎顶端颜色变浅,茎基部宽(22−)30–43(−55) μm,顶端宽(14−)17–26(−33) μm,长(473–)829–1 400(−1 756) μm。子囊壳产孢孔附近菌丝呈发散状,透明。子囊未见。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团;子囊孢子侧面观呈钢盔状,无隔,透明,具鞘,带鞘长(4.1−)5.7–6.8(−7.5) μm,宽(2.5−)3.1–3.9(−4.6) μm。
(2)无性阶段:主分生孢子梗逐一单独的产生于菌丝表面,透明,基部较宽大,顶部较尖锐,长(42−)60–145(−291) μm,基部(3.2−)4–6(−7.7) μm,顶端(2.7−)3.9–5.5(−7.4) μm;次分生孢子梗逐一单独的产生于菌丝表面,透明,长(52−)57–69(−68) μm,基部宽(3−)2–4(−5) μm,顶端宽4−6 μm。分生孢子透明,无隔,圆柱形至哑铃形,(8.8−)16.2–25.6(−49.9)×(2.7−)3.4−4.6(−2.7) μm。厚垣孢子椭圆形,壁厚,表面光滑,褐色,(9.9−)12.1–15(−16.7)×(7−)9.2–11.5(−13) μm,嵌入培养基,独生或链接成念珠状。
菌落特点:菌落呈橄榄绿,菌丝竖立生长,表面或嵌入培养基生长,最适生长温度25℃,14 d后长满90 mm培养基。5℃以下或35℃以上停止生长。菌丝表面光滑,菌丝具隔,隔处未缢痕。在10℃、15℃、20℃、25℃和30℃下生长14 d后,在90 mm培养基上的菌落大小依次为:7、29、50、73和57 mm。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,广东、广西、福建和海南。
标本信息:中国,海南省,临高县。2013年9月,spp.,主模式标本:PREM 61229,主模式标本上分离的模式菌株:CMW 43029 = CERC 2170 = CBS 139654。
其他菌株信息:中国,广西,玉林市,博白县。2013年10月,spp.,菌株标本:PREM 61230,标本所分离到的菌株:CMW 42515 = CERC 2345 = CBS 139655;中国,广东省,江门市,新会市。2014年1月,spp.,菌株标本:PREM 61231,标本所分离到的菌株:CMW 42795 = CERC 2687 = CBS 139656。
1.2 Ceratocystis collisensis F. F. Liu, M. J. Wingf. & S.F. Chen sp. nov. [22]
MycoBank号码:MB811889
病原中文名称:武夷山长喙壳
形态特征:
(1)有性阶段:子囊壳基部暗褐色至黑色,球形至倒梨形,基部宽(134−)164–224(−255) μm,高(152−)174–253(−304) μm,表面附着菌丝;子囊壳茎暗褐色至黑色,至茎顶端颜色变浅,茎基部宽(18−)25–33(−39) μm,顶端宽(12−)15–20(−25) μm,长(208–)301–423(−527) μm。子囊壳产孢孔附近菌丝呈发散状,透明。子囊未见。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团;子囊孢子侧面观呈钢盔状,无隔,透明,具鞘,带鞘长(5.6−)6.5–7.8(−8.4) μm,宽(2.7−)3.6–4.6(−5.3) μm。
(2)无性阶段:主分生孢子梗逐一单独的产生于菌丝表面,透明,基部较宽大,顶部较尖锐,长(46−)47–337(−129) μm,基部(2.3−)4.2–6.2(−7.7) μm,顶端(2.3−)3.6–4.9(−6.2) μm;次分生孢子梗逐一单独的产生于菌丝表面,透明,长(15−)34–79(−121) μm,基部宽(2.8−)3.8–5(−6.4) μm,顶端宽(3.3−)4.5–7(−9) μm。分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形至哑铃形,(10.7−)14.5–21.3(−32.3)×(2.9−)3.4−4.4(−5.6) μm;次分生孢子透明,无隔,木桶状,成链状,(5.4−)6.1–8.7(−10.9)×(3.7−)4.4−6.3(−7.9) μm。厚垣孢子椭圆形,壁厚,表面光滑,深褐色,(9.2−)11.3–14.4(−17.6)×(7.2−)8.8–11.1(−13.7) μm,嵌入培养基,独生或链接成念珠状。
菌落特点:菌落呈橄榄绿,菌丝竖立生长,表面或嵌入培养基生长,最适生长温度 25℃,5℃以下或35℃以上停止生长。菌丝表面光滑,菌丝具隔,隔处未缢痕。在10℃、15℃、20℃、25℃和30℃下生长14 d后,在90 mm培养基上的菌落大小依次为:10、30、42、50和41 mm。
中国分离来源:近期砍伐的新鲜的杉木()树桩。
中国寄主以及分布:杉木,中国福建。
标本信息:中国,福建省,漳州市,长泰县。2013年10月,,主模式标本:PREM 61232,主模式标本上分离的模式菌株:CMW 42552 = CERC 2459 = CBS 139679。
其他菌株信息:中国,福建省,南平市,建瓯县。2013年11月,,菌株标本:PREM 61233,标本所分离到的菌株:CMW 42553 = CERC 2465 = CBS 139646;中国,福建省,南平市,建瓯县。2013年11月,,菌株标本:PREM 61234,标本所分离到的菌株:CMW 42554 = CERC 2466 = CBS 139647。
1.3 Ceratocystis changhui F.F. Liu, I. Barnes & S.F. Chen sp. nov. [23]
MycoBank号码:MB815611
病原中文名称:长喙长喙壳
形态特征:
(1)有性阶段:子囊壳基部褐色至黑色,椭圆形至球形,半透明状,基部宽(139−)153.5–209(−261) μm,高(132.5−)154–207.5(−257) μm,表面附着菌丝;子囊壳茎暗褐色至黑色,至茎顶端颜色变浅,茎基部宽(28.5−)37–52.5(−63.5) μm,顶端宽(19.5−)24.5–35.5(−47) μm,长(453.5–)594–810(−896.5) μm。子囊壳产孢孔附近菌丝呈发散状,透明。子囊未见。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团;子囊孢子侧面观呈钢盔状,无隔,透明,具鞘,带鞘长(4.5−)5–6(−6.5) μm,宽(2.5−)3–3.5(−4) μm。
(2)无性阶段:主分生孢子梗逐一单独的产生于菌丝表面,透明,基部较宽大,顶部较尖锐,长(43.5−)60–95(−122) μm,基部(3.5−)4.5–6(−7.5) μm,顶端(3−)3.5–4.5(−6) μm。分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(6.5−)10.5–16.5(−22.5)×(3.5−)4.5−5.5(−6.5) μm;次分生孢子透明,无隔,木桶状,成链状,(3.5−)5.5–9(−10.5)×(3.5−)4−5.5(−6) μm。厚垣孢子椭圆形,壁厚,表面光滑,深褐色,(8.5−)10–12.5(−14)×(76−)8–10(−11.5) μm,嵌入培养基,独生或链接成念珠状。
菌落特点:菌落呈橄榄绿,菌丝竖立生长,表面或嵌入培养基生长,最适生长温度 25℃,5℃、10℃或35℃以上停止生长。菌丝表面光滑,菌丝具隔,隔处未缢痕。在15℃、20℃、25℃和30℃下生长14 d后,在90 mm培养基上的菌落大小依次为:30.1、38.8、58.3和33.6 mm。
中国分离来源:芋头()黑腐病病斑。
中国寄主以及分布:芋头(),中国云南,山东。
标本信息:中国,云南省,昆明市,长水镇。2014年8月,,主模式标本:PREM 61232,主模式标本上分离的模式菌株:CMW 43281 = CERC 3615 = CBS 139797。
其他菌株信息:中国,云南省,昆明市,长水镇。2014年8月,,菌株标本:PREM 61242,标本所分离到的菌株:CMW 43272 = CERC 3605 = CBS 139798;中国,云南省,昆明市,长水镇。2014年8月,,菌株标本:PREM 61243,标本所分离到的菌株:CMW 43273 = CERC 3606 = CBS 139799。
1.4 Huntiella ani F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826735
病原中文名称:安亨特长喙壳
形态特征:
(1)有性阶段:子囊壳基部暗褐色至黑色,球形至倒梨形,基部表面具有刺突和菌丝,刺突暗褐色至黑色,长(6−)8.5–15.5(−21) μm;子囊壳基部宽(104.5−)124–167(−224) μm,高(117.5−)128.5–173(−232) μm;子囊壳茎暗褐色至黑色,茎基部宽(29.5−)33–44.5(−54) μm,顶端宽(9.5−)10.5–12.5(−14) μm,长(493−)520–667(−845) μm,基部盘状;子囊壳产孢孔附近菌丝呈发散状,透明,长(14.5−)19.5–28.5(−33) μm。子囊未见。子囊孢子侧面观呈钢盔状,具鞘,无隔,透明,带鞘长(4.5−)4.5–5.5(−6) μm,宽(1.5−)2.5–3(−3.5) μm。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(18.5−)27–45.5(−59) μm,基部(2−)3–4.5(−5.5) μm,顶端宽(1.5−)1.5–2.5(−3) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(4−)5–8.5(−12.5)×(1.5−)1.5−2.5(−3) μm;次分生孢子透明,无隔,木桶状,成链状,(4−)5–7.5(−9)×(2.5−)3−4(−5) μm。未见厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度25℃,在5℃、30℃和35℃停止生长,在10℃、15℃、20℃和25℃下生长4 d后,在90 mm培养基上的菌落大小依次为:7.1、32.2、51.7和66.4 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,中国广西。
标本信息:中国,广西,南宁市。2014年1月,spp.,主模式标本:PREM 62020,主模式标本上分离的模式菌株:CMW 44686 = CBS 143282 = CERC 2829。
其他菌株信息:中国,广西,南宁市。2014年1月,spp.,菌株标本:PREM 62021,标本所分离到的菌株:CMW 44684 = CBS 143283 = CERC 2827;中国,广西,柳州市。2014年1月,spp.,菌株标本:PREM 62022,标本所分离到的菌株:CMW 49315 = CBS 143284 = CERC 2871。
1.5 Huntiella bellula F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826738
病原中文名称:秀丽亨特长喙壳
形态特征:
(1)有性阶段:子囊壳基部暗黑色,球形至倒梨形,基部表面具有刺突和菌丝,刺突暗褐色至黑色,长(5−)7.5–16(−20) μm;子囊壳基部宽(172.5−)179–228.5(−250.5) μm,高(184−)195–230.5(−244) μm;子囊壳茎暗褐色至黑色,茎基部宽(42.5−)45–59.5(−69) μm,顶端宽(10.5−)11.5–13.5(−14.5) μm,长(395−)450–631(−707.5) μm,基部盘状;子囊壳产孢孔附近菌丝呈发散状,透明,长(18−)20–29.5(−32.5) μm。子囊未见。子囊孢子侧面观呈钢盔状,具鞘,无隔,透明,带鞘长(5−)5.5–6.5(−6.5) μm,宽(2.5−)2.5–3(−4) μm。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(21.5−)26.5–38.5(−50) μm,基部宽(2−)2.5–4(−6) μm,顶端宽(1−)1.5–2(−2.5) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(5.5−)6.5–9(−12)×(1.5−)1.5−2(−2) μm。未见次分生孢子和厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度25℃,在35℃停止生长,在5℃、10℃、15℃、20℃、25℃和30℃下生长4 d后,在90 mm培养基上的菌落大小依次为:5.3、10.5、33.6、54.4、75.5和5.7 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,中国广西。
标本信息:中国,广西,柳州市。2014年1月,spp.,主模式标本:PREM 62023,主模式标本上分离的模式菌株:CMW 49314 = CBS 143285 = CERC 2862。
其他菌株信息:中国,广西,柳州市。2014年1月,spp.,菌株标本:PREM 62024,标本所分离到的菌株:CMW 49312 = CBS 143286 = CERC 2854;中国,广西,来宾市。2014年1月,spp.,菌株标本:PREM 62025,标本所分离到的菌株:CMW 49316 = CBS 143287 = CERC 2880。
1.6 Huntiella confusa F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826740
病原中文名称:台湾相思亨特长喙壳
形态特征:
(1)有性阶段:未观察到。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(17−)20–33(−48) μm,基部宽(2−)2.5–4.5(−6.5) μm,顶端宽(1.5−)1.5–2(−3) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(4.5−)5.5–7.5(−10)×(1.5−)1.5−2(−3) μm。未见次分生孢子和厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度30℃,在5℃停止生长,在10℃、15℃、20℃、25℃、30℃和35℃下生长4 d后,在90 mm培养基上的菌落大小依次为:2.3、19、40.5、59.5、69.3和30.4 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:倒掉的台湾相思树()伤口。
中国寄主以及分布:台湾相思树(),中国海南。
标本信息:中国,海南省,临高县。2013年9月,,主模式标本:PREM 62026,主模式标本上分离的模式菌株:CMW 43453 = CBS 143288 = CERC 2162。
其他菌株信息:中国,海南省,临高县。2013年9月,,菌株标本:PREM 62027,标本所分离到的菌株:CMW 49300 = CBS 143289 = CERC 2141;中国,海南省,临高县。2013年9月,,菌株标本:PREM 62028,标本所分离到的菌株:CMW 43452 = CBS 143577 = CERC 2158。
1.7 Huntiella eucalypti F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826741
病原中文名称:桉树亨特长喙壳
形态特征:
(1)有性阶段:未观察到。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(24.5−)27–44.5(−56) μm,基部宽(2.5−)2.5–3.5(−4.5) μm,顶端宽(1.5−)1.5–2(−2) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(4−)6–8.5(−11)×(1.5−)1.5−2(−2.5)μm。未见次分生孢子和厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度25℃,在35℃停止生长,在5℃、10℃、15℃、20℃、25℃和30℃下生长4 d后,在90 mm培养基上的菌落大小依次为:10.5、14.4、32.9、53.2、67.5和4.5 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,中国广西。
标本信息:中国,广西,河池市。2014年1月,spp.,主模式标本:PREM 62029,主模式标本上分离的模式菌株:CMW 44693 = CBS 143290 = CERC 2841。
其他菌株信息:中国,广西,河池市。2014年1月,spp.,菌株标本:PREM 62030,标本所分离到的菌株:CMW 44692 = CBS 143291 = CERC 2840。
1.8 Huntiella fabiensis F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826743
病原中文名称:法比亨特长喙壳
形态特征:
(1)有性阶段:子囊壳基部暗黑色,球形,基部表面具有刺突和菌丝,刺突暗褐色,长(7−)8–15.5(−18) μm;子囊壳基部宽(146.5−)163.5–219.5(−244.5) μm,高(158.5−)163–222.5(−259.5) μm;子囊壳茎暗褐色至黑色,茎基部宽(34−)37.5–57.5(−71) μm,顶端宽(10.5−)11.5–15(−17.5)μm,长(254.5−)316–431.5(−470.5) μm,基部盘状;子囊壳产孢孔附近菌丝呈发散状,透明,长(12.5−)16.5–27.5(−28.5) μm。子囊未见。子囊孢子侧面观呈钢盔状,具鞘,无隔,透明,带鞘长(5.5−)6–7(−8) μm,宽(2−)3–3.5(−4) μm。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(19−)24.5–52(−75.5) μm,基部宽(2.5−)3–4(−5.5) μm,顶端宽(1.5−)1.5–3(−4) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(4−)6–10(−13.5)×(1.5−)1.5−2(−2.5) μm;次分生孢子透明,无隔,木桶状,成链状,(3−)5–8(−10.5)×(2.5−)3−5(−7) μm。未见厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度25℃,在35℃停止生长,在5℃、10℃、15℃、20℃、25℃和30℃下生长4 d后,在90 mm培养基上的菌落大小依次为:7.7、16.7、35.5、59.6、70.9和3.4 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,中国广东。
标本信息:中国,广东省,梅州市。2014年1月,spp.,主模式标本:PREM 62031,主模式标本上分离的模式菌株:CMW 49309 = CBS 143292 = CERC 2763。
其他菌株信息:中国,广东省,梅州市。2014年1月,spp.,菌株标本:PREM 62032,标本所分离到的菌株:CMW 44370 = CBS 143293 = CERC 2736;中国,广东省,梅州市。2014年1月,spp.,菌株标本:PREM 62033,标本所分离到的菌株:CMW 49307 = CBS 143294 = CERC 2753。
1.9 Huntiella fecunda F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826744
病原中文名称:多子座亨特长喙壳
形态特征:
(1)有性阶段:子囊壳基部暗褐色至黑色,球形至倒梨形,基部表面具有刺突和菌丝,刺突暗褐色,长(5−)7–18.5(−23) μm;子囊壳基部宽(92.5−)106–173(−234) μm,高(111−)118.5–184(−253) μm;子囊壳茎暗褐色至黑色,茎基部宽(23.5−)29.5–47.5(−66.5) μm,顶端宽(9.5−)10–12(−13.5)μm,长(365.5−)521–828(−1052.5) μm,基部盘状;子囊壳产孢孔附近菌丝呈发散状,透明,长(15−)18–23.5(−25.5) μm。子囊未见。子囊孢子侧面观呈钢盔状,具鞘,无隔,透明,带鞘长(4.5−)5–5.5(−5.5) μm,宽(2−)2–2.5(−3) μm。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(16−)18.5–32.5(−53) μm,基部宽(2−)2.5–3.5(−5.5) μm,顶端宽(1.5−)1.5–2(−2.5) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(4−)5–7(−8.5)×(1.5−)1.5−2(−3) μm。未见次分生孢子及厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度25 ~ 30℃,在5℃和35℃停止生长,在10℃、15℃、20℃、25℃和30℃下生长4 d后,在90 mm培养基上的菌落大小依次为:4.8、29.2、45.6、61和63.7 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,中国福建。
标本信息:中国,福建省,漳州市。2013年10月,spp.,主模式标本:PREM 62034,主模式标本上分离的模式菌株:CMW 49303 = CBS 143295 = CERC 2451a。
其他菌株信息:中国,福建省,漳州市。2013年10月,spp.,菌株标本:PREM 62035,标本所分离到的菌株:CMW 49301 = CBS 143304 = CERC 2446;中国,福建省,漳州市。2013年10月,spp.,菌株标本:PREM 62036,标本所分离到的菌株:CMW 49302 = CBS 143296 = CERC 2449。
1.10 Huntiella glaber F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826745
病原中文名称:平滑亨特长喙壳
形态特征:
(1)有性阶段:子囊壳基部黑色,球形,基部表面具有刺突和菌丝,刺突暗褐色,长(8−)8–17.5(−26) μm;子囊壳基部宽(133−)152.5–210.5(−237) μm,高(128.5−)146.5–207.5(−228) μm;子囊壳茎暗褐色至黑色,茎基部宽(42−)49–67.5(−78.5) μm,顶端宽(8.5−)10–15(−23) μm,长(259.5−)328–548.5(−722.5) μm,基部盘状;子囊壳产孢孔附近菌丝很少或缺失。子囊孢子侧面观呈钢盔状,具鞘,无隔,透明,带鞘长(4.5−)5.5–6.5(−7) μm,宽(2−)2.5–3(−3.5) μm。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(19−)25–45.5(−79) μm,基部宽(2.5−)3.5–4.5(−6) μm,顶端宽(1.5−)2–2.5(−3) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(7−)8.5–12(−15.5)×(1.5−)2−3(−3) μm。未见次分生孢子及厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度30℃,在5℃停止生长,在10℃、15℃、20℃、25℃、30℃和35℃下生长4 d后,在90 mm培养基上的菌落大小依次为:5.7、26.4、56、79.6、86和42.3 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:窿缘桉()伤口。
中国寄主以及分布:窿缘桉(),中国海南。
标本信息:中国,海南省,澄迈县。2013年9月,,主模式标本:PREM 62037,主模式标本上分离的模式菌株:CMW 49299 = CBS 143297 = CERC 2133。
其他菌株信息:中国,海南省,澄迈县。2013年9月,,菌株标本:PREM 62038,标本所分离到的菌株:CMW 43436 = CBS 143298 = CERC 2132。
1.11 Huntiella inaequabilis F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826746
病原中文名称:凸凹亨特长喙壳
形态特征:
(1)有性阶段:未观察到。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(27.5−)35–56(−72) μm,基部宽(2.5−)2.5–4(−7) μm,顶端宽(1.5−)1.5–2(−2) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(4−)4.5–9(−17.5)×(1.5−)1.5−2(−2) μm。次分生孢子透明,无隔,木桶状,成链状,(4.5−)5.5–7.5(−9.5)×(3.5−)3.5−5(−6) μm。未见厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度25℃,在35℃停止生长,在5℃、10℃、15℃、20℃、25℃和30℃下生长4 d后,在90 mm培养基上的菌落大小依次为:4.3、9.1、19.1、26.5、31.6和14.6 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,中国广东。
标本信息:中国,广东省,梅州市。2014年1月,spp.,主模式标本:PREM 62039,主模式标本上分离的模式菌株:CMW 49306 = CBS 143299 = CERC 2749。
其他菌株信息:中国,广东省,梅州市。2014年1月,spp.,菌株标本:PREM 62040,标本所分离到的菌株:CMW 44372 = CBS 143300 = CERC 2740。
1.12 Huntiella meiensis F.F. Liu & S.F. Chen, sp. nov. [24]
MycoBank号码:MB826747
病原中文名称:梅州亨特长喙壳
形态特征:
(1)有性阶段:子囊壳基部黑色,球形至椭圆形,基部表面具有刺突和菌丝,刺突暗褐色,长(6−)8–17(−28) μm;子囊壳基部宽(119.5−)139–195(−219.5) μm,高(129.5−)145–194.5(−220) μm;子囊壳茎暗褐色至黑色,茎基部宽(33−)40–59(−77.5) μm,顶端宽(10−)11–13(−14.5) μm,长(290−)299.5–347.5(−416.5) μm,基部盘状;子囊壳产孢孔附近菌丝很少或缺失。子囊孢子侧面观呈钢盔状,具鞘,无隔,透明,带鞘长(4.5−)5.5–6.5(−7) μm,宽(2−)2.5–3.5(−4) μm。子囊孢子聚集呈浅黄色,在子囊壳茎顶端粘聚成团。
(2)无性阶段:分生孢子梗逐一单独的产生于菌丝表面,透明,长(28−)31.5–52(−73) μm,基部宽(3−)3.5–4.5(−6) μm,顶端宽(1.5−)1.5–2.5(−2.5) μm;分生孢子有两种类型,初级分生孢子透明,无隔,圆柱形,(4−)5–7.5(−10.5)×(1.5−)1.5−2.5(−3.5) μm;次分生孢子透明,无隔,木桶状,成链状,(5.5−)6–8.5(−11)×(3.5−)4−5.5(−6.5) μm。未见厚垣孢子。
菌落特点:菌落在MEA培养基上白色,菌丝竖立生长,最适生长温度25℃,在5℃和35℃停止生长,在10℃、15℃、20℃、25℃和30℃下生长4 d后,在90 mm培养基上的菌落大小依次为:12.7、38.4、60.2、71.5和5.8 mm。菌丝表面光滑或者有颗粒状突起,菌丝具隔,隔处未缢痕。
中国分离来源:砍伐1个月左右的桉树树桩。
中国寄主以及分布:桉树,中国广东。
标本信息:中国,广东省,梅州市。2014年1月,spp.,主模式标本:PREM 62041,主模式标本上分离的模式菌株:CMW 44376 = CBS 143301= CERC 2746。
其他菌株信息:中国,广东省,梅州市。2014年1月,spp.,菌株标本:PREM 62042,标本所分离到的菌株:CMW 44374 = CBS 143302 = CERC 2742;中国,广东省,梅州市。2014年1月,spp.,菌株标本:PREM 62043,标本所分离到的菌株:CMW 49305 = CBS 143303 = CERC 2744。
2 讨论
长喙壳属()真菌是长喙壳科(Ceratocystidaceae)最大的属,包括许多重要的植物病原菌,能侵染多种植物寄主,例如树木以及块茎作物等。长喙壳属()病原菌能引起多种感病症状,例如植物维管组织变色、枝干溃疡、枯萎、黑腐病、甚至导致树木迅速死亡等。病原菌需要伤口来感染寄主,寄主伤口部位被感染后,病原菌可在短期内引起感病部位的变色、腐烂,一段时间(最快7 ~ 10 d)后生成成熟的子囊果。另外,有些物种可以分泌出芳香的气味,能吸引昆虫携带并传播。相对于,则被普遍认为致病性较弱,甚至大多数物种仅作为腐生菌存在,只有在树木自身生长受到胁迫时候才会导致轻微的病症,针对物种的致病性需要进一步的研究来明确。
在过去的20年间,由病原菌引起的林木特别是桉树和相思树上的病害被广泛的报道,包括一些引起林木短期内枯萎和死亡的重大病害。本文所报道的和新物种,主要是在我国华南地区的桉树人工林及周边其他人工林中被分离和鉴定得到,这是针对我国华南地区人工林中和的首次大范围的系统研究报道。致病力测试结果表明,对桉树有较强的致病力,能引起芋头块茎黑腐病发生,9个新物种在接种的桉树苗茎干上可产生病斑。在调查过程中发现,广泛分布于广东、广西、海南、云南和福建等省(区),为中国所报道物种的优势物种。在桉树以及相思被广泛种植的东南亚其他国家和地区,病原菌给桉树以及相思人工林的健康发展带来了巨大的危害,而且它们对人工林潜在的威胁常常被低估。为确保我国桉树以及相思人工林避免遭受的威胁,有必要对菌在我国桉树以及相思人工林中的分布进行全面系统的调查和研究,进而选育出抗病的林木基因型,最终降低桉树和相思人工林由病原菌带来的危害。
[1] HALSTED B D. Some fungus diseases of the sweet potato [J]. New Jersey Agricultural Experiment Station Bulletion, 1890,76: 7-14.
[2] UPADHYAY H P. A monograph ofand[M]. Athens: University of Georgia Press, 1981.
[3] WINGFIELD M J, SEIFERT K A, WEBBER J F. Ceratocystis andOphiostoma,[M]//Hanssen HP. Volatile metabolitesproduced by species ofand. St Paul: APS Press, 1993: 117-126.
[4] WINGFIELD M J, DE BEER Z W, SLIPPERS B, et al. One fungus, one name promotes progressive plant pathology [J]. Molecular Plant Pathology, 2012,13(6): 604-613.
[5] WINGFIELD M J, BARNES I, DE BEER Z W, et al. Novel associations between ophiostomatoid fungi, insects and tree hosts: current status-future prospects [J]. Biological Invasions, 2017,19(11): 3215-3228.
[6] HAUSNER G, REID J, KLASSEN G. Ceratocystis and Ophiostoma,[M]//Grouping of isolates and species ofon the basis of molecular and morphological characters. 1993: 93-104.
[7] HAUSNER G, REID J, KLASSEN G. On the phylogeny of,, and, and relationships withinbased on partial ribosomal DNA sequences [J]. Canadian Journal of Botany, 1993,71(9): 1249-1265.
[8] HAUSNER G, REID J, KLASSEN G R. On the subdivision of, based on partial ribosomal DNA sequences [J]. Canadian Journal of Botany, 1993,71(1): 52-63.
[9] SPATAFORA J W, BLACKWELL M. The polyphyletic origins of ophiostomatoid fungi [J]. Mycological Research, 1994,98(1): 1-9.
[10] WINGFIELD B D, VAN WYK M, ROOS H, et alThe ophiostomatoid fungi: expanding frontiers. [M]//: emerging evidence for discrete generic boundaries. 2013: 57-64.
[11] DE BEER Z W, DUONG T, BARNES I, et al. Redefiningand allied genera [J]. Studies in Mycology, 2014,79: 187-219.
[12] DE BEER Z W, MARINCOWITZ S, DUONG T A, et al., a new genus to accommodate the oak wilt fungus,(Microascales, Ascomycota) [J]. MycoKeys, 2017,27: 1-19.
[13] MAYERS C G, MCNEW D L, HARRINGTON T C, et al. Three genera in the Ceratocystidaceae are the respective symbionts of three independent lineages of ambrosia beetles with large, complex mycangia [J]. Fungal Biology, 2015,119(11): 1075-1092.
[14] MAYERS C G, HARRINGTON T C, MASUYA H, et al. Patterns of coevolution between ambrosia beetle mycangia and the Ceratocystidaceae, with five new fungal genera and seven new species [J]. Persoonia, 2020,44: 41-66.
[15] NEL W J, DUONG T A, WINGFIELD B D, et al. A new genus and species for the globally important, multi-host root pathogen[J]. Plant Pathology 2018,67(5): 871-882.
[16] VON SCHRENK H. The “bluing” and the “red-hot” of the western yellow pine, with special reference to the Black Hills Forest Reserve [J]. U.S. Department of Agriculture,Bureau of Plant Industry Bulletin, 1903,36: 1-46.
[17] HEDGCOCK G G. Studies upon some chromogenic fungi which discolor wood [J]. Missouri Botanical Garden Annual Report, 1906,17: 59-124.
[18] MOREAU C. Coexistence of the formsandin a strain ofmajor (van Beyma) n. comb. Remarks on variations in[J]. Revue de Mycologie, 1952,17(suppl): 17-25.
[19] DAVIDSON R W. Fungi causing stain in logs and lumber in the southern states, including five new species [J]. Journal of Agricultural Research, 1935,50: 789-807.
[20] VAN WYK M, ROUX J, BARNES I, et al. Molecular phylogeny of thecomplex and description ofsp. nov. [J]. Fungal Diversity, 2006, 21: 181-201.
[21] MBENOUN M, WINGFIELD M J, BEGOUDE BOYOGUENO A D, et al. Diversity and pathogenicity of the Ceratocystidaceae associated with cacao agroforests in Cameroon [J]. Plant Pathology, 2016,65(1): 64-78.
[22] LIU F F, MBENOUN M, BARNES I, et al. Newspecies fromandin South China [J]. Antonie van Leeuwenhoek, 2015,107(6): 1451-1473.
[23] LIU F F, BARNES I, ROUX J, et al. Molecular phylogenetics and microsatellite analysis reveal a new pathogenicspecies in the Asian Australian Clade [J]. Plant Pathology, 2018,67(5):1097-1113.
[24] LIU F F, LI G Q, ROUX J,et al. Nine novel species offrom southern China with three distinct mating strategies and variable levels of pathogenicity [J]. Mycologia, 2018,110(6): 1145-1171.
[25] 陈帅飞,李国清,周建清,等.桉树枝干枯萎病研究进展[J].桉树科技,2013,30(4):34-43.
[26] KILE G A. HARRINGTON T C, YUAN Z Q, et al.sp. nov., a vascular stain fungus from eucalypts in Australia [J]. Mycological Research, 1996, 100(5): 571-579.
[27] 陈帅飞.中国桉树真菌病原汇录:2006—2013[J].桉树科技,2014,31(1):37-65.
[28] ENGELBRECHT C J B, HARRINGTON T C. Intersterility, morphology and taxonomy ofon sweet potato, cacao and sycamore[J]. Mycologia, 2005, 97(1): 57-69.
[29] ROUX J, WINGFIELD M J.species: emerging pathogens of non-native plantationandspecies [J]. Southern Forests: a Journal of Forest Science, 2009, 71(2): 115-120.
[30] AL ADAWI A O, BARNES I, KHAN I A, et alassociated with a serious wilt disease of two native legume trees in Oman and Pakistan [J]. Australasian Plant Pathology, 2013, 42(2): 179-193.
[31] BARNES I, FOURIE A, WINGFIELD M J, et al. Newspecies associated with rapid death ofin Hawaii [J]. Persoonia, 2018, 40: 154-181.
[32] 陆道调,吴保国,王希群,等.相思树种研究发展综述[J]. 福建林学院学报,2004,24(1):92-96.
[33] 谢耀坚.世纪初的桉树研究[M].北京:中国林业出版社,2006.2014–2020
Research Advances ofandin China:2014–2020
LIU Feifei, CHEN Shuaifei
()
The Ascomycete genusoccurs on a wide range of hosts and has a global distribution. Information on the species identification, species diversity, origin and impact of these fungal pathogens in China is, however, limited. Recently, large numbers ofisolates as well as of the closely related genushave been collected from different hosts in five provinces of China.andisolates were obtained from different tree hosts, including fresh cut stumps and wounds ofspecies,species and, as well as from some rotten corms of(taro). In this study, a checklist ofandspecies described from China during 2014 to 2020 is supplied and for each of these fungi a concise description is provided of its: taxonomic status; morphological characteristics; culture morphology; host range; and, geographical distribution. These fungi include three novel species of, namelyfrom,fromandfrom, along with nine novel species of, namelyfromspecies,,,,,,,andobtained fromplantations. This study provides a greater understanding of the host range and distribution of these important genera of fungi in China. Further studies will consider the biology of these fungi, their genetics and their population biology.
forest health; tuber crop diseases; plantations forestry;; fungal pathogens
10.13987/j.cnki.askj.2021.02.013
S718.81
A
国家重点研发计划项目“桉树高效培育技术研究”课题“桉树高效可持续经营技术”(2016YFD0600505)
刘菲菲(1987— ),博士,博士后,助理研究员,主要从事森林病害研究,E-mail: shelleyfei@163.com
陈帅飞(1982— ),博士,研究员,主要从事森林病害研究,E-mail:cerccsf@126.com
