基于MaxEnt模型的苦槠潜在地理分布格局变迁预测
2021-06-08许晓岗
缪 菁,王 勇,王 璐,许晓岗
(1.南京林业大学生物与环境学院,江苏 南京 210037;2.南京市绿化园林局,江苏 南京 210019)
人类活动与气候因素通过影响植被生境条件决定了植物现状分布,其变化会导致潜在地理分布格局的变迁,物种分布是对气候变化的响应[1]。因此研究植物分布与气候之间的变化关系,可预测某一物种潜在的适生区域,对于揭示物种分化与形成、迁移与扩散的历史有着重要意义,更对驯化培育、物种保护和种质资源管理等生产实践具有指导意义。
近年来,科学家广泛应用各种物种分布模型取代传统物种调查来进行大尺度物种分布格局的模拟和预测[1],这类模型综合运用了地理信息系统(geograplic information system, GIS)及统计学、生态学的相关理论,是寻找植物的分布区、限制因子、生境条件等的有效工具[2]。笔者所选用的MaxEnt模型(最大熵模型)属于目前物种分布模型中精确度较高的一类,它是以最大熵原理和机器学习理论为基础[3],利用已知物种分布数据和环境变量模拟物种现代潜在地理分布情况的生态位模型[1-2]。MaxEnt模型的优势在于它的计算不受野外工作和样本数量的限制且预测精度较高[2-4]。
栲属(Castanopsis)是我国长江流域以南常绿阔叶林的优势属,苦槠(C.sclerophylla)作为我国亚热带常绿阔叶次生林的常见种,是栲属中最为耐寒、在我国分布最北的一种,是划分亚热带和温带的标志树种,南京目前被认为是其分布的最北缘[5]。笔者以苦槠为研究对象,根据其全国范围内地理分布信息和气候环境数据,运用MaxEnt模型对其在末次盛冰期以来的地理分布变化和未来潜在分布进行分析,探讨气候因子变化对苦槠潜在地理分布的影响,利用其生物属性监测亚热带最北缘的动态变化,为苦槠的种源保护、生境恢复、繁育驯化等提供科学依据。
1 材料与方法
1.1 样本来源与筛选
以70°~140°E、17°~57°N区域范围作为研究区。在中国数字标本馆(CVH,http://www.cvh.org.cn)、教学标本资源共享平台(http://mnh.scu.edu.cn/main.aspx)、全球生物多样性信息网络(GBIF,http://www.gbif.org)、中国植物图像库(PPBC,http://ppbc.iplant.cn/)等数字标本平台上共检索到919条苦槠分布记录。首先删除人工引种栽培的标本,其次为了降低取样的偏差,消除集群效应[6],在每个2.5′×2.5′网格中只取唯一1个分布点,最后共得到有效的苦槠分布记录317个。
1.2 气候变量选取与处理
从世界气候数据库(http://worldclim.org)下载末次盛冰期(LGM)、现代和RCP 8.5背景下2070年的气候数据,利用DIVA-GIS V7.5.0软件从317个分布点上提取了19个气候变量的数值,并利用SPSS V13.0对这些气候变量进行Pearson相关性分析[7-8]。对于相关性较高(R> 0.8)的数据,仅选择一个与地理分布关系密切的用于模型预测,以此共筛选出10个变量(表1)。利用ArcGIS V10.0软件对气候变量图层进行剪裁,剪裁范围与研究区域相同。
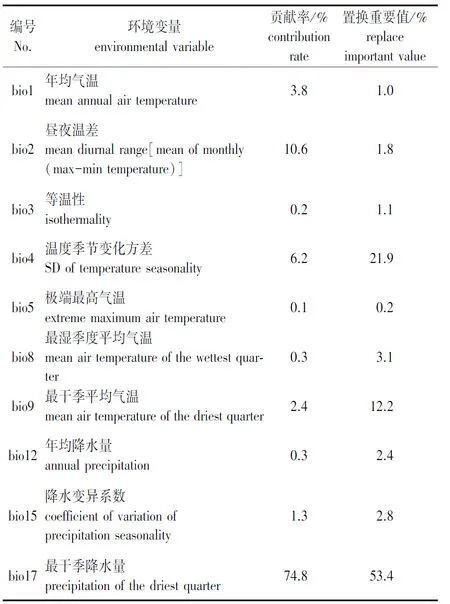
表1 气候变量及其贡献值
1.3 模型建立、优化与评估
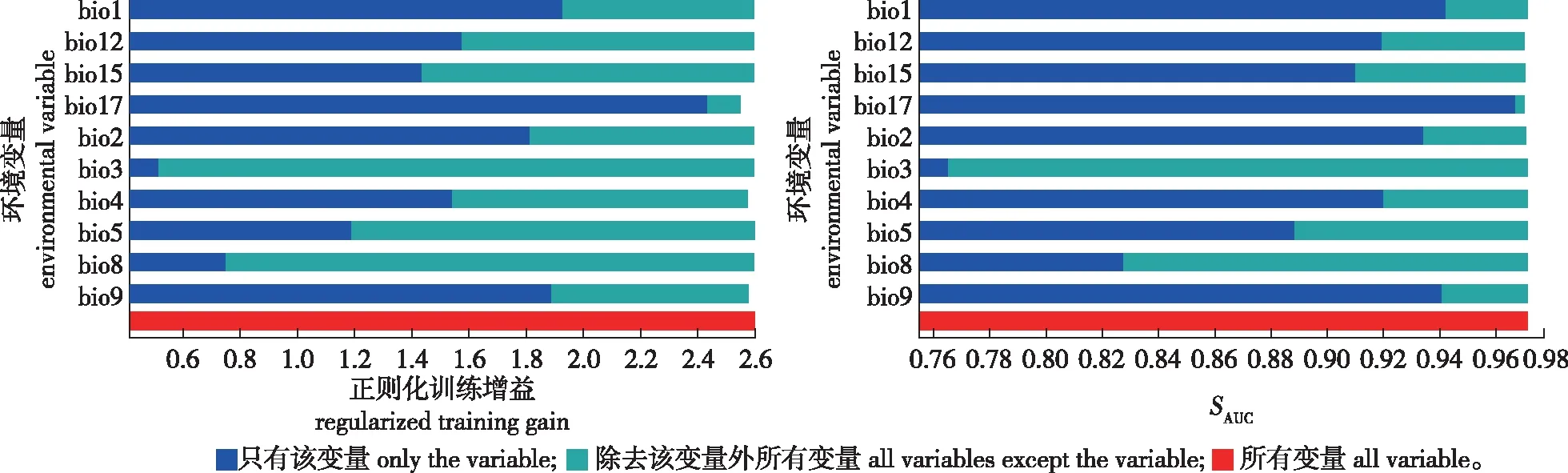
将筛选出的317个苦槠分布信息和10个环境变量导入MaxEnt 3.3.3k软件,以模型默认值为参数设置,利用ArcGIS V10.0空间分析模块计算现代、未来气候条件下苦槠的适生区域及分布区。选择交叉验证法(cross validate)作为测试方法,将所有苦槠分布记录分为10个子集,其中9个作为训练集,1个作为验证集。使用受试者工作特征曲线(receiver operating characteristic curve,ROC)下的面积(areas under curve, AUC,记为SAUC)对MaxEnt模型的预测结果进行精度检测,该值越大表示与随机分布相距越远,环境变量与预测的物种地理分布之间相关性越大,模型效果越好[9-11],具体指标如下:SAUC> 0.9(非常好);0.8 利用贡献率、置换重要值和Jackknife检验来评估环境变量与现代潜在地理分布的关联性。其三者的工作原理不尽相同,贡献率是通过最大熵程序获得最优解的算法;置换重要值以结果为导向,无关具体算法;Jackknife检验通过单独使用或删除某个环境变量来构建模型,对比不同模型的正则化训练增益,通过测试AUC值和测试增益之间的差异评估环境变量的重要性[12-14]。 通过MaxEnt模型对317个苦槠有效分布记录进行计算,对苦槠在末次冰盛期、现代和2070年的地理分布格局进行模拟和预测。结果显示AUC 值的平均值为0.971,预测结果准确度较高。在ArcGIS中产生栅格图层,按照适宜生境指数和逻辑值阀值,将苦槠在各时期的生境分为基本分布区、中度适宜区和高度适宜区。 从苦槠现代分布点及MaxEnt模型预测的不同时期潜在分布区(图1)可以看出,MaxEnt模型显示苦槠在现代的基本分布区主要位于淮河以南、五岭以北地区,包括安徽与江苏东南部、浙江、江西、湖北、湖南至福建北部、广西北部等地,呈连续分布,另外,日本九州一带也有分布。高度适宜区主要在安徽南部山区、浙江西部莫干山至天目山、浙东北雁荡山腹地、江西东南部及福建武夷山地区。中度适宜区包括苏南山区、皖南至浙西山区、江西庐山至井冈山及湖南沅江以南地区、福建东北部地区。同时结合标本有效记录多度推断苦槠的分布中心应为皖南与浙西交界山区,以及闽赣交界部分的山区。 审图号:GS(2017)3320图1 苦槠现代分布点及MaxEnt模型预测的苦槠不同时期潜在分布区Fig.1 Extant occurrence points (blue dots) and potential distributions of Castanopsis sclerophylla during different periods predicted by MaxEnt model 末次盛冰期发生在20000—18000 年 BP,气温比现代至少要低12 ℃[15]。这一时期苦槠分布范围较小,高度适生区域连续性减弱,与现代分布相比呈现向低纬度、低海拔区域退缩,主要集中在我国华南北部地区,包括湘赣交界的罗霄山中东部、衡山以及湘桂黔交界地区。同时在东海大陆架东部,至琉球群岛北部,日本九州和四国也有零星分布。到2070年,预测苦槠的分布整体呈现向东北高纬度地区迁移趋势,适生区急剧缩小并出现断裂,中度适生区破碎度提高,高度适生区聚集在浙东沿海北雁荡山附近和日本的九州四国地区。 由环境变量及其贡献率表(图2)可以看出,环境因子对苦槠分布贡献率从大到小依次为: 最干季降雨量(bio 17)、平均日温差(bio2)、温度季节变化方差(bio4)。置换重要值最高的3个环境变量分别为最干季降雨量(bio17)、气温季节变化方差(bio4)、最干季平均气温(bio9)。而图2显示,Jackknife分析对苦槠分布影响最大的环境因子中排在前三位的分别为最干季降雨量(bio17)、年均气温(bio1)和最干季平均气温(bio9);AUC值最高的前三位是最干季降雨量(bio17)、年均气温(bio1)、最干季平均气温(bio9)。使用除此变量以外的其他变量,正则化训练增益降低最多的依次是最干季降雨量(bio17)、温度季节变化方差(bio4)、最干季平均气温(bio9)。AUC值降低最多的是最干季平均气温(bio9)和温度季节变化方差(bio4),其余基本相似。由此可见,本模型中影响苦槠分布的主要气候因素为降水因子(最干季降雨量)和温度因子(最干季平均气温、温度季节变化方差)。 图2 苦槠现代分布区环境变量重要性的Jackknife 检验Fig.2 The jackknife test of variable importance of current potential distribution MaxEnt模型预测结果显示,在末次盛冰期时,苦槠的高度适宜区为我国华南地区,特别是湖南以南和湘黔桂交界地区。孢粉证据表明末次盛冰期时我国亚热带常绿阔叶林带退缩至华南南部[16-17],这一区域是被指定的25个全球生物多样性热点中心之一,因其地形复杂,没有遭受与冰期有关的极端寒冷和干旱,形成了有利于物种生存的小生境[18],因而,这一地区可能为苦槠在末次盛冰期的避难所。苦槠现代地理分布的最北缘位于江苏南部,如南京、张家港等地,而本研究发现末次盛冰期时这一边缘居群所在地并非苦槠的适生区。群体遗传学研究表明,南京牛首山、苏州弯窿山、宜兴龙池山等地的苦槠居群在冰期中可能经历了显著的瓶颈效应,由此可以推测,江苏南部包括南京的苦槠居群在末次盛冰期可能经历过灭绝,现在位于分布区北缘的苦槠居群可能是冰期后重新扩散而来的,也就是说苦槠在冰期的分布区北缘并不是现在的南京,而是位于低纬度地区[19],这与本研究的预测结果一致。同时,末次盛冰期时,东亚古地理的另一最大变化在于大片的陆架出露成陆[20]。MaxEnt模型预测当时苦槠在东海至日本九州、四国等高纬度地区也有分布,暗示了这一树种可能以东海大陆架为桥梁从日本逐步向南迁移。 进入现代,气候转暖使植物分布普遍存在向高纬度地区扩张的趋势,中东部地区温润的气候和丰富的降水更适于苦槠的生长,苦槠高度适宜区也由西南内陆地区向华东沿海地区转移、扩展。随着人类社会的不断发展,人口向东北部迁移,鼠类、鸟类也随之迁移,而苦槠作为壳斗科栲属植物,种子淀粉含量高,是啮齿动物秋冬季节的主要食物来源之一[21],因此,其种子传播很可能通过啮齿动物和鸟类在东部地区广泛传播和扩散。 本研究中,选取RCP8.5气候变化情景预测2070年苦槠的未来分布情况。模拟预测结果表明,在未来全球变暖气候条件下到2070年,苦槠的潜在分布总面积和位置与现代分布区基本相似,但中、高度适宜区面积显著缩小,向东部沿海急剧收缩,高度适宜区锐减,在高纬度地区几乎消失,仅存在于浙江东部沿海地区和日本的九州、四国地区,分布区连续性减弱,破碎度增高,在华中地区发生断裂,尤其是江西南部、湖北及湖南北部适宜区丢失明显。有研究预测,在未来气候情景下,21世纪中期到后期30 a,随着气候变暖,东亚夏季风将增强且向我国北方推进,尤其是西南气流显著增加,导致东部地区长江中下游及其以北地区极端降水将大范围增加,东部地区春、秋季冷日减少,热日增多[22,这可能是造成苦槠高度适生区不断向东部沿海及日本海附近降水丰富地区东移的原因。而21世纪后期,湖北、湖南、江西三省交界处的气候变化程度与周边地区有差异,表现为夏季和秋季热日增加率比周边地区小,秋季冷日和热日比周边地区略多[23],这种区域小气候造成的水热不均衡,可能是造成苦槠在江西南部、湖南、湖北适宜区丢失的原因。而华南地区温度上升,降水减少,极端高温事件发生频率明显升高,这些变化可能会导致未来气候条件下华南地区分布的苦槠适宜区也不断向东部沿海地区集中。这也预示在未来应加强对这一地区苦槠野生资源的动态监测,及时掌握其种群变化状况。 气候因素是影响物种分布的重要因素之一。刘茂松等[24]研究表明栲属植物的物种丰富度分布主要与年降水、最高气温呈负相关,与1月平均气温、最小月降水量呈正相关,也就是说适合栲属植物生存的气候条件应是年降水量小而最干季降水量大,常年保持一定湿度,最干季平均气温要高,但最高气温要低。而本研究在对MaxEnt模型预测贡献排名前列的气候因子进行分析时发现,影响苦槠分布的最主要气候因素分别是最干季降雨量、最干季平均气温和温度季节变化方差。最干季的降水反应地区极端水条件,最干季均温反应水热平衡情况,这与栲属的适生环境研究结果完全吻合。而目前苦槠主要分布的长江中下游,华东、华中等地区,受亚热带季风气候影响,夏季高温多雨,冬季温和少雨,1月平均气温普遍在0 ℃以上,昼夜温差小,年降水量一般在800~1 000 mm,均符合本研究所证明的苦槠适生气候特征。苦槠分布对气候的响应也源自自身的生理特征,苦槠种子含水量较高[25],极端干旱或冬季寒冷的环境下易造成种子失水、丧失活力,从而限制其自然分布。 本研究利用MaxEnt模型对苦槠在末次盛冰期、现代和2070年3个时期的地理潜在分布构建进行了模拟和预测,分析得出最干季降水量和最干季平均气温是影响苦槠地理分布的最主要环境因素,温暖湿润、水热平衡的适生环境有利于其生长,这也与模型预测所显示的未来其高度适宜区不断向东南沿海低海拔温润地区迁移相对应,这对于应对气候变化、制定科学有效的苦槠资源生态保护对策具有重要意义[26]。但鉴于未来气候变化的不确定性,预测结果只是基于当前模型中的气候因子进行测算,对地形、土壤及人工干扰等因素考虑较少。因此,随着对未来气候变化认知水平的提高,可以对预测结果不断地加以完善。1.4 环境变量的重要性评估
2 结果与分析
2.1 苦槠现代地理分布格局
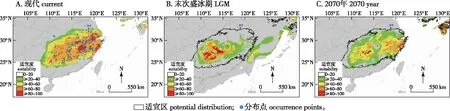
2.2 苦槠在末次盛冰期和未来的潜在分布
2.3 环境变量重要性
3 讨 论
3.1 不同时期苦槠地理分布的变迁
3.2 苦槠未来地理分布预测
3.3 影响苦槠分布的主要气候因子
