大黄素通过调节Nrf2/HO-1和MAPKs抑制炎症和氧化应激机制研究①
2021-05-25张昊悦王业皇南京中医药大学附属南京中医院肛肠中心南京210000
张昊悦 赵 蓓 王业皇 章 阳(南京中医药大学附属南京中医院肛肠中心,南京210000)
炎性肠病是一系列导致多种并发症的疾病,包括克罗恩病和溃疡性结肠炎,其中最常见的是肠纤维化和癌症[1]。溃疡性结肠炎和克罗恩病患者的黏膜炎症常表现为固有层淋巴细胞、巨噬细胞和中性粒细胞浸润[2]。巨噬细胞在维持肠道稳态中起着至关重要的作用,并且被认为是炎性肠病的关键参与者。经典激活的M1型产生一氧化氮、活性氧和促炎细胞因子,活动期炎性肠病患者肠道黏膜中的一氧化氮表达骤增,可抑制超氧化物歧化酶(su⁃peroxide dismutase,SOD)活性、促使脂质自由基形成,导致肠黏膜通透性增加、黏膜屏障功能受损[3]。并且它在诸如伤口愈合和组织的建设过程中起维修作用[4]。因此抑制巨噬细胞M1型活化可影响炎性肠病的发生发展。氧化应激是指机体受到多种病理因素刺激后,体内ROS产生过多,抗氧化能力下降,打破了机体正常氧化还原动态平衡,造成生物大分子如蛋白质、脂质、核酸等的氧化损伤,干扰正常生命活动而形成的一种严重的应激状态[5]。ROS来源增多和抗氧化能力降低是导致氧化应激水平增高的主要机制,Nrf2信号通路是细胞内氧化应激最重要的防御系统之一,当细胞发生氧化应激,Nrf2信号通路首先被激活,进而引发抗氧化物及相关酶类的大量表达,以抵御氧化应激引发的细胞损伤,减少ROS产生[6]。Nrf2可以调控具有重要的抗氧化和抗炎活性的HO-1,而HO-1在大鼠结肠炎模型有重要保护作用,HO-1的表达能抑制促炎介质的产生[7-8]。
尽管已开发出各种抗炎药物,例如水杨酸类制剂、硫唑嘌呤等免疫抑制剂药物和生物制剂,但由于这些药物的肝肾损伤、过敏反应等副作用较多,目前仍需要研发具有抗炎作用同时减少副作用的中药单体。
大黄素(Emodin,1,3,8-三羟基-6-甲基蒽醌)是一种常见的蒽醌衍生物,据报道具有多种生物学特性,例如抗炎、抗病毒、抗肿瘤和抗氧化活性[9-12]。根据文献所知大黄素能够通过抗氧化和抗炎活性减轻重症急性胰腺炎,大黄素可以通过上调lncRNA TUG1抑制LPS诱导的小鼠软骨ATDC5细胞凋亡和炎症[13-14]。大黄素可通过抑制大鼠结肠炎NF-κB炎症通路产生抗炎作用,但是关于大黄素对细胞氧化应激反应的作用未见相关报道[15]。但是大量文献表明天然蒽醌类药物能够抑制细胞体内的ROS[16]。本研究拟探讨大黄素能否通过Nrf-2/HO-1和MAPKs抑制炎症,并减轻氧化应激反应。
1 材料与方法
1.1 材料
1.1.1 药物和细胞 中药单体大黄素(中国药检所);小鼠单核巨噬细胞系(RAW264.7)购自中国科学院上海科学研究院细胞库。用含有10%胎牛血清高糖DMEM培养基培养于37℃、5%CO2培养箱。
1.1.2 主要试剂 CCK8试剂盒(同仁化学研究所);Griess试剂盒(上海碧云天);活性氧(ROS)检测试剂盒(美国Solarbio公司);RNA提取试剂(美国Sigma公司);Perfect real time-PCR试剂盒(日本Ta⁃KaRa公司);SYBR Green RCR Kit(QuantiNova公司);PCR引物设计和合成(生工生物工程有限公司);核因子E2相关因子(nuclear factor E2 related factor 2,Nrf2)(1∶1 000)、血红素氧化酶1(hemeoxy⁃genase1,HO-1)(1∶1 000)、细胞外信号调节激酶(ERK)及其磷酸化(pERK)(1∶1 000)、p38丝裂原激活的蛋白激酶(p38)及其磷酸化(pP38)(1∶1 000)和GAPDH抗体(CST)(1∶1 000),均为兔抗,兔二抗(美国Jackson ImmunoResearch Laboratories)(1∶10 000);超敏ECL化学反应发光试剂盒(新赛美生物科技有限公司)。
1.1.3 主要仪器 CO2培养箱、超净工作台、酶标仪(瑞士Tecan公司);Cytoflex流式细胞仪(美国贝克曼库尔德公司)。
1.2 方法
1.2.1 CCK8法测定大黄素对RAW264.7细胞的毒性影响 将RAW264.7,接种于96孔板,每孔1×105个细胞,置于培养箱(37℃、5%CO2)中孵育12 h。加入不同浓度的大黄素继续培养24 h,浓度为25、50、100、200、300µmol/L,设定3个平行组,24 h后,每孔加入10µl的CCK8试剂,放置培养箱孵育1.5 h,酶标仪(450 nm)测吸光度值(A)。细胞活力计算公式为:细胞活力(%)=各浓度给药组的A值/空白组的平均A值×100%。
1.2.2 Griess法检测大黄素对LPS诱导RAW264.7细胞炎症介质NO的影响 RAW264.7细胞接种于96孔板培养基,每孔1×105个细胞。过夜后细胞贴壁,弃掉培养基后,分别设置对照组、LPS(LPS1µg/ml)、大黄素低剂量组(LPS1µg/ml+大黄素20µmol/L),大黄素高剂量组(LPS1µg/ml+大黄素40µmol/L)。加入各组药物后孵育24 h后,吸取50 µl的上清液至空白的96孔板,设置标准曲线,分别加入50µl Griess试剂A和Griess试剂B,孵育10 min,用酶标仪570 nm处测定各孔OD值,根据标准曲线,计算每组的NO的含量。
1.2.3 RT-PCR法检测LPS诱导的细胞的IL-6、iN⁃OS、IL-1βmRNA的含量 RAW264.7细胞培养于6孔板,分组及处理同1.2.2,每孔5×105个细胞,贴壁生长过夜。加药处理24 h后收集各组RNA样品。用Trizol提取方法细胞总的RNA,用Perfect Real Time试剂盒逆转录提取总cDNA,反应体系37℃、15 min,85℃5 s,4℃保存,后用RT-PCR进行检测(引物如表1)。PCR反应条件:95℃2 min,95℃5 s,60℃10 s,共40个循环。重复3次,β-肌动蛋白(β-actin)作为内标基因。使用2-ΔΔCt法计算mRNA含量。
1.2.4 流式细胞术检测大黄素对RAW264.7细胞ROS的影响 细胞培养和处理同1.2.2,作用24 h后,收集细胞,用4℃PBS冲洗2次,10µmol/L DCF-DA 37℃孵育30 min后,用PBS清洗2遍,流式上机检测。
1.2.5 流式细胞术检测大黄素对LPS诱导的RAW264.7细胞表面膜蛋白CD86的影响 细胞培养和分组处理同1.2.2。用预冷的PBS冲洗2遍,PBS重悬细胞,调整细胞浓度为1×107个/ml,取100µl到流式管中,加入PE抗鼠CD86抗体以及IgG2a的同型对照组,4℃避光孵育20 min;1 000 r/min离心5 min,收集细胞,弃上清,再加入200µl的PBS;用Cytoflex流式细胞仪检测。以Flow Jo软件分析结果。
1.2.6 检测大黄素作用于RAW264.7细胞Nrf-2/HO-1蛋白表达 RAW264.7细胞铺于6孔板,加入不同组别的药物作用,24 h后,消化收集细胞,用预冷的PBS洗涤细胞,每孔加入RIRA裂解液100µl,收集细胞于1.5 ml离心管中。冰上放置30 min,然后4℃,12 000 g离心15 min,取上清液,蛋白定量试剂盒进行蛋白定量。加入蛋白上样缓冲液,金属浴10 min变性。使用10%SDS-PAGE电泳分离,200 mA恒流转膜90 min,5%脱脂奶粉,室温封闭1 h;4℃过夜孵育一抗(1∶1 000),一抗为Nrf-2和HO-1。TBTS溶液洗3次,每次10 min,室温孵育二抗(1∶20 000)1 h;TBST溶液洗3次,每次10 min,然后使用ECL发光法检测蛋白。使用Image J软件对图像进行分析。
1.2.7 检测大黄素对LPS诱导的RAW264.7细胞MAPK信号通路的蛋白表达 细胞培养和分组处理同1.2.2。后续处理同1.2.6,一抗为pERK、ERK、pP38和P38,内参为GAPDH。
1.3 统计学方法 应用GraphPad Prism 6统计软件分析,独立样本采用t检验分析,计量资料用±s描述,组间比较采用单因素方差分析(ANOVA),以P<0.05表示差异有统计学意义。

表1 引物序列Tab.1 Primer sequence
2 结果
2.1 大黄素对RAW264.7细胞的毒性作用 大黄素在浓度为0~40µmol/L时对正常的RAW264细胞活力无毒性作用,超过40µmol/L,细胞活力下降(见图1)。所以本研究选用了20、40µmol/L两个浓度进行后续的研究。
2.2 大黄素抑制LPS诱导的RAW264.7细胞中IL-6、iNOS、IL-1βmRNA表达 根据图2可以得知,LPS组中IL-6、iNOS、IL-1β细胞因子的mRNA表达与对照组相比较差异有统计学意义(P<0.001);大黄素组与LPS组相比,炎症因子IL-6、iNOS和IL-1β mRNA随着药物剂量的增加而降低,差异具有显著的统计学意义(P<0.001)。
2.3 大黄素对RAW264.7细胞中NO含量的影响如图3所示,与对照组相比,LPS诱导的炎症RAW264.7细胞中NO含量明显增加;与LPS组比较,不同浓度作用的大黄素中NO含量明显降低(P<0.001),且具有浓度依赖性。
2.4 大黄素抑制LPS诱导的ROS的产生 图4结果显示,与对照组相比,LPS单独处理组的ROS含量具有统计学意义;大黄素组与LPS组相比能够明显的抑制ROS生成(P<0.001),且具有浓度依赖性。
2.5 大黄素对RAW264.7膜表面蛋白CD86的影响 如图5可知,LPS组高表达CD86膜表面蛋白分子,和对照组相比具有统计学意义(P<0.001)。给予大黄素干预极化的RAW264.7细胞,RAW264.7细胞的CD86明显的降低,且具有浓度依赖性。

图1 大黄素对RAW264.7细胞活性的影响Fig.1 Effect of Emodin on activity of RAW264.7 cell
2.6 大黄素能够促进RAW264.7细胞的Nrf-2/HO-1蛋白的表达 由图6可知,LPS组的Nrf-2和HO-1蛋白的表达明显降低(P<0.05)。大黄素组与LPS组相比,能显著Nrf-2和HO-1蛋白的表达(P<0.05,P<0.01),且存在浓度依赖性。
2.7 大黄素对LPS诱导的MAPK信号通路相关蛋白的影响 如图7所示,与对照组相比,LPS刺激的RAW264.7细胞,ERK、p38磷酸化水平明显升高(P<0.01),与LPS组相比,大黄素组的ERK、p38磷酸化水平明显降低(P<0.05,P<0.01)。结果表明大黄素能够抑制MAPK的信号通路。

图2 大黄素对LPS诱导的RAW264.7细胞中IL-6、iNOS、IL-1βmRNA的影响Fig.2 Effect of Emodin on IL-6,iNOSand IL-1βmRNA in LPS-induced RAW264.7 cell
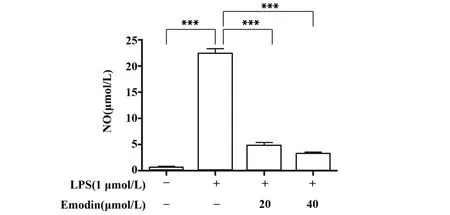
图3 大黄素对LPS诱导的RAW264.7细胞NO产生的影响Fig.3 Effect of Emodin on NO production in LPS-in⁃duced RAW264.7 cell
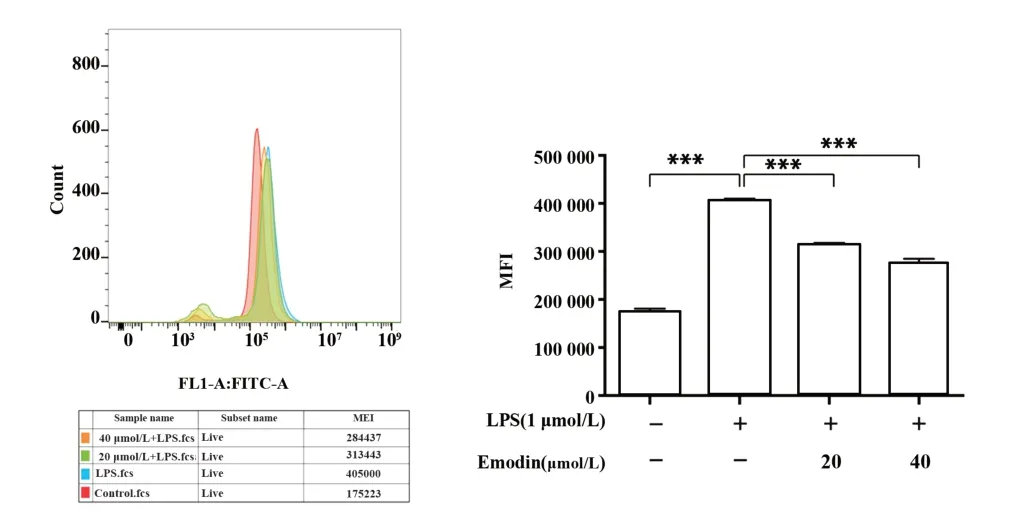
图4 大黄素对LPS诱导的RAW264.7细胞中ROS产生的影响Fig.4 Effect of Emodin on ROS production in LPS-in⁃duced RAW264.7 cell
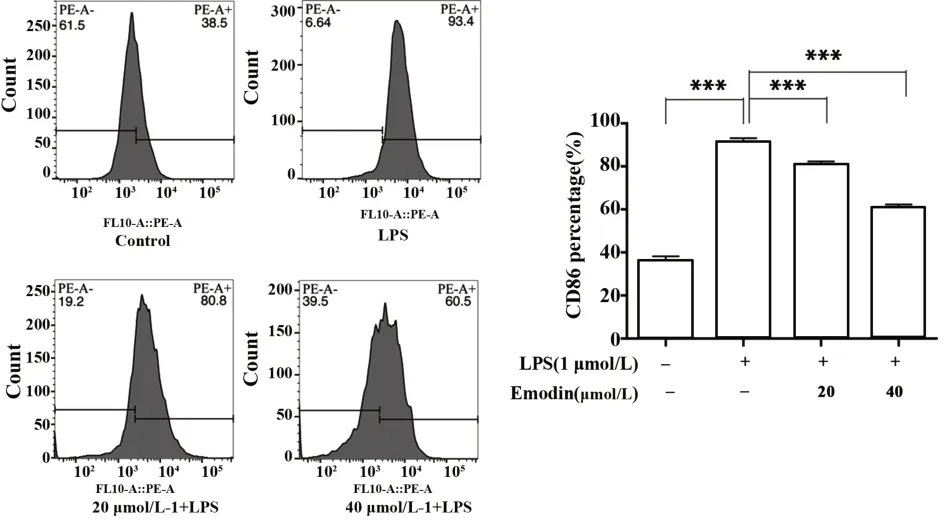
图5 大黄素对于LPS诱导的RAW264.7细胞膜蛋白CD86的影响Fig.5 Effect of Emodin on CD86 in LPS-induced RAW-264.7 cell

图6 大黄素对LPS诱导的RAW264.7细胞中Nrf2、HO-1蛋白表达的影响Fig.6 Effect of Emdin on expression of Nrf2,HO-1 in LPSindued RAW264.7 cell
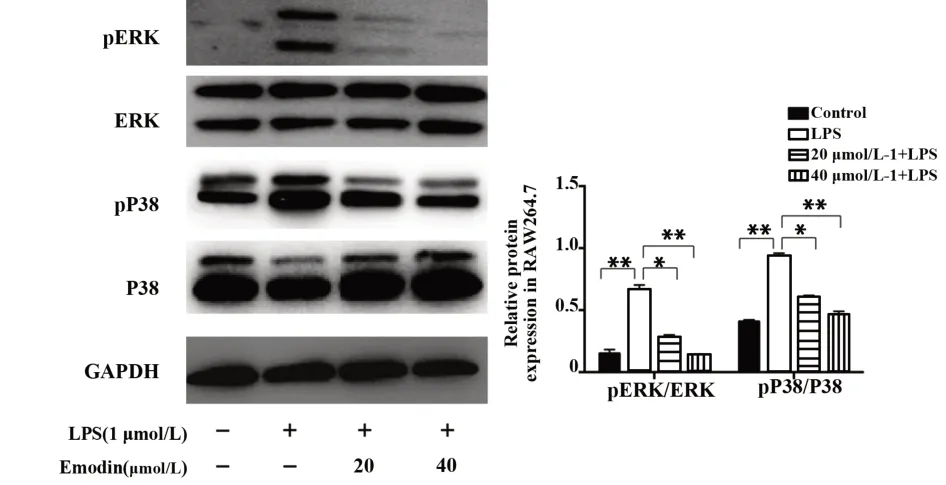
图7 大黄素对LPS诱导的RAW264.7细胞ERK、P38磷酸化蛋白表达的影响Fig.7 Effect of Emodin on expression of p ERK,p P38 in LPS-induced RAW264.7 cell
3 讨论
单核细胞和巨噬细胞是炎症和先天免疫反应的关键效应和调节细胞[17]。巨噬细胞可由革兰氏阴性菌的脂多糖LPS等有害物刺激产生多种炎症介质,如NO,iNOS,IL-6和IL-1β等[18]。这些促进炎症的介质大量产生导致了炎症[19-20]。因此大量研究在寻找这些促炎介质的抑制剂和抗炎促进剂[20]。
脂多糖(LPS)诱导的RAW264.7细胞炎症模型是经典的体外炎症模型。LPS是被Toll样受体4(TLR4)识别并启动细胞内信号转导,并能活化细胞的核因子κ轻链增强子(NF-κB)和丝裂原活化蛋白激酶K(MAPK);当MAPK相关蛋白如细胞外信号调节激酶(ERK)和p38丝裂原激活的蛋白激酶(p38 MAPK)信号激活,导致哺乳动物中的炎性介质如一氧化氮表达(NO)的产生,活化的巨噬细胞上的TNF-α和IL-6升高,同时能够上调活性氧(ROS)[20-24]。ROS又称活性氧,其产生与炎症密切相关,炎性肠病中,活化的巨噬细胞和中性粒细胞会持续过量产生ROS,使炎症条件和不受控制的免疫系统氧化负荷增加[25]。炎性肠病的发展过程中会出现高水平的ROS,产生氧化应激,导致体内稳态丧失,随后出现细胞氧化应激损伤,包括脂质过氧化产物、蛋白质修饰和促炎症细胞因子的过度生成[26]。同时局部炎症细胞因子水平升高,特别是IL-1β、IL-6、IL-12、IL-17、IL-23、IFN-γ和TNF-α,是炎性肠病免疫系统失控的重要特征[27]。因此本研究采用了这个模型模拟了炎性肠病疾病过程中发生的炎症反应。
研究报道,一切抗氧化酶,如血红素加氧酶(HO-1),参与iNOS和NO的调控作用,生理状态下HO-1低表达,可因为缺氧和ROA等刺激后可迅速增加,HO-1增加有助于细胞对刺激的反应,HO-1能够保护RAW264.7免受ROS的攻击,HO-1是核转录因子E2相关因子2(Nrf2)信号通路的下游蛋白HO-1主要受Nrf2的调控,所以Nrf2/HO-1和MAPK通路一样,都能够抑制炎症和氧化应激反应[28-29]。最近,一些关于炎性肠病的研究将注意力集中在Nrf-2上,它是一种重要的转录因子,参与了包括肠道在内的许多器官的纤维化过程,同时已经证实Nrf-2参与了包括结直肠癌在内的多种肿瘤的发展、进展和转移[29-31]。
大量研究证实了很多中药及其主要活性都具有能够抑制炎症和氧化应激[32-34]。本研究采用了LPS刺激RAW264.7细胞制备体外炎症模型。大黄素对于RAW264.7具有低细胞毒性,可以抑制LPS诱 导的RAW264.7细胞,降 低IL-6、iNOS、IL-1β mRNA的表达,且抑制效应呈一定的浓度依赖性。研究表明天然蒽醌类药物能够抑制RAW264.7中ROS的产生[16]。与此同时,我们通过实验证实了大黄素能使细胞中ROS和NO水平显著降低和细胞膜蛋白CD86含量的减少,表明大黄素具有抗炎作用,能够降低由LPS引起的细胞氧化应激反应。有研究表明大黄素在体外能够通过抑制TLR4信号通路减轻LPS诱导的急性肝损伤,可以降低体内外TLR4下游蛋白的水平,并且还能抑制细胞培养上清液和血清中TNF-α和IL-6的表达[35]。大黄素能够大黄素通过PPARγ依赖途径抑制脂多糖诱导的RAW264.7细胞炎症[36]。虽然大黄素对已经证实了对RAW264.7细胞具有很强的抗炎效应,但是对抑制RAW264.7细胞的炎症和ROS的表达没有做深入的机制研究,所以我们对此进行了探讨。为了进一步阐明大黄素在介导炎症和氧化应激反应中的分子机制,通过Western blot实验,检测大黄素对LPS诱导后的RAW264.7的Nrf-2、HO-1和MAPK通路表达的影响。本研究结果显示大黄素能够显著增加由LPS诱导的后细胞中Nrf-2、HO-1的蛋白表达,对细胞起到一定的保护作用。同时大黄素还能够通过抑制MAPK通路的活化从而对LPS诱导的炎症反应起到抑制作用。
综上所述,大黄素能够调节Nrf2/HO-1信号和MAPK信号通路抑制LPS诱导的RAW264.7细胞炎症和氧化应激作用,这些信号通路与炎性肠病密切相关。本研究进一步探索了大黄素体外抗炎的活性及其机制,为大黄素治疗炎性肠病提供证据,为临床应用奠定基础。
