孕鼠肥胖对子代雄鼠早期肝脏脂肪代谢的影响和机制研究
2021-05-13祝峰韩曙光徐珂滕懿群
祝峰 韩曙光 徐珂 滕懿群
母代肥胖会导致子代成年期肥胖、心血管疾病、非酒精性脂肪肝(nonalcoholic fatty liver disease,NAFLD)和脂肪性肝炎形成[1]。Baker[2]认为早期生活环境的改变会增加子代的表型和代谢紊乱风险。动物实验表明母体孕期肥胖、哺乳期高脂饮食会导致小鼠活动和进食节律改变,并会在小鼠成年后导致代谢异常,包括糖代谢异常、脂代谢异常、NAFLD等[3]。自噬是真核细胞特有的一种分解代谢过程,其在维持细胞能量代谢平衡、控制细胞器质量、细胞应激应答中发挥重要作用。研究表明,转录因子 EB(transcription factor EB,TFEB)在肝脏中增加可以调节溶酶体和自噬相关基因表达,进而诱导肝脏自噬表达的增加,从而有效改善肝脏脂肪变性[4]。此外,研究还发现去乙酰化酶3(sirtuin3,SIRT3)过表达可以导致锰超氧化物歧化酶的去乙酰化和活化,在耗尽细胞的超氧化物后实现自噬的抑制,促使肝脏脂滴的形成[5]。临床研究发现母代肥胖和母乳喂养方式改变会导致后代17岁时更容易发生NAFLD[6],具体的发病机制尚不明确。Devarajan等[7]通过研究发现,早期宫内环境变化会使子代产生营养敏感记忆,通过自噬调节在晚年对子代肝脏代谢产生保护作用。对此,本研究提出假设,肥胖母鼠子代在早期会发生肝脏改变,并且自噬可能在其中发挥作用。目前肝代谢的研究主要集中在成年期,但是关于母代肥胖的子代早期肝脏脂滴形成易感性,及其与自噬之间的关系研究较少。因此,本研究通过高脂喂养构建肥胖母鼠模型,以探究母代肥胖及哺乳期高脂饮食对子代肝脏早期脂滴形成的易感性及可能的分子机制。
1 材料和方法
1.1 实验动物 4周龄SPF级C57BL/6J雌鼠20只及雄鼠10只,购自苏州大学,被安置在嘉兴学院实验动物中心。饲养条件为(24±2)℃,相对湿度(50±10)%,12 h/12 h明暗循环照明,自由获取水和食物。实验过程中,雄鼠均普通粮喂养。本实验经嘉兴市第二医院动物伦理委员会审批通过,实验过程中严格遵守动物实验原则指南(1996修订版)的要求。
1.2 试剂和仪器 TC(A111-1-1)、TG(A110-2-1)、HDL(A112-1-1)、LDL(A113-1-1)试剂盒均购自中国南京建成生物有限公司;Trizol、反转录试剂盒均购自日本Takara公司;自噬相关基因Beclin-1、自噬相关基因3(autophagy associated gene 3,ATG3)、自噬相关基因5(autophagy associated gene 5,ATG5)、自噬相关基因 12(autophagy associated gene 12,ATG12)、p62 及轻链 3B(light chain 3B,LC3B)等mRNA引物由中国通用生物技术公司合成;Beclin-1(A10101)、ATG3(A5809)、ATG5(A0203)、ATG12(A19610)、p62(A7758)、LC3B(A5618)、生物素二抗(AS014)等蛋白抗体均购自中国ABclonal公司;微管蛋白(Tubulin)(KM9003)购自天津 Sungene Biotech公司;多功能酶标仪(SPARK)购自瑞士TECAN公司;普通PCR仪购自德国Eppendorf公司;Thermo Fisher-QS3实时定量PCR仪购自美国Thermo公司;Bio-Rad凝胶成像仪购自美国Bio-Rad公司。
1.3 方法
1.3.1 动物分组与喂养 采用随机数字表法将20只雌鼠分为肥胖母鼠(obese maternal,OM)组和普通母鼠(common maternal,CM)组,每组10只。OM组雌鼠接受高脂饲料(含60%脂肪,江苏南通特洛菲生物有限公司)喂养;CM组雌鼠接受普通饲料(含4%脂肪,江苏南通特洛菲生物有限公司)喂养,喂养4周后,两组雌鼠出现明显体重差异。将普通雄鼠与雌鼠按1∶2进行交配,它们的后代被定义为OM子代鼠(OM offspring,OM-O)、CM子代鼠(CM offspring,CM-O)。两组母鼠哺乳期间统一高脂饲料喂养,按照每只母鼠2 g/d进行饲喂。两组子代鼠均母乳喂养4周,并在4周时断乳,同时每组随机选取7只子代雄鼠,麻醉后分别收集眼球静脉血和肝脏组织,并保存于-80℃冰箱备用。所有子代雄鼠均采用脱颈椎法处死。
1.3.2 肝脏病理切片检测 采用油红O染色法。取4周龄子代雄鼠的新鲜肝脏右叶小块组织,迅速放入4%多聚甲醛中固定肝脏样本,石蜡包块,4 μm厚切片,常规油红O染色(红色代表脂滴沉积),显微镜下观察肝脏组织变化。
1.3.3 血清脂代谢指标检测 采用ELISA法。取4周龄子代雄鼠眼球静脉血,收集的全血4℃隔夜放置,在4℃低温下3 500 r/min离心10 min,取上清液。严格按照试剂盒操作说明书检测TC、TG、HDL、LDL水平。
1.3.4 自噬相关基因mRNA表达水平检测 采用RTPCR法。用Trizol试剂从肝脏组织中提取总RNA。在Thermo Fisher-QS3实时定量PCR仪上检测Beclin-1、ATG3、ATG5、ATG12、p62及 LC3B 等 mRNA 表达情况。以β-actin为内参。实验步骤:变性程序(95℃10 min),扩增定量程序重复40次(95℃ 15 s,60℃ 10 s,72℃和15 s,单荧光测量),熔融曲线程序(60~95℃,加热速率0.1 C/s,连续荧光测量)。采用 2-ΔΔCt法测定相对 mRNA的表达。相关基因引物序列见表1。

表1 相关基因引物序列
1.3.5 自噬相关基因蛋白表达水平检测 采用Western blot法。提取肝脏组织总蛋白,使用BCA蛋白质测定试剂盒测定总蛋白浓度。将各样本的蛋白浓度调整一致,再与1/4体积的蛋白上样缓冲液混合后煮沸,变性;通过10%十二烷基硫酸钠-聚丙烯酰胺凝胶(SDS-PAGE)电泳分离总蛋白,转膜;在室温下用5%脱脂奶粉封闭膜2 h,按照 1:1 000 稀释 Beclin-1、ATG3、ATG5、ATG12、p62及LC3B等抗体原液,根据分子量大小分别以Tubulin作为内参,于4℃过夜孵育;第2天洗膜,用5%脱脂奶粉稀释HRP标记的二抗(1∶1 000)于摇床上孵育90 min,洗膜后,加发光液,用Bio-Rad成像系统曝光,采用Image J软件测定灰度值,目标蛋白分别与内参灰度值相比为各自相对表达水平。
1.3.6 肝脏组织LC3B含量检测 采用免疫组化染色法。肝脏组织切片以新鲜配置体积分数为3%的H2O2灭活细胞内源性过氧化物酶,然后用血清封闭,根据测试指标分别滴加LC3B以及相应生物素二抗AS014;DAB显色,封片。采用Image J软件对肝脏中LC3B的积分光密度(IOD)值进行定量分析。
1.4 统计学处理 采用SPSS 22.0统计软件。计量资料以表示,方差齐时,组间比较采用两独立样本t检验;方差不齐时,组间比较采用Mann-Whitney U检验。P<0.05为差异有统计学意义。
2 结果
2.1 两组子代雄鼠肝脏组织病理变化比较 与CM-O组比较,OM-O组肝脏切片中脂滴形成不明显,肝脏脂滴沉积较轻,见图1(插页)。
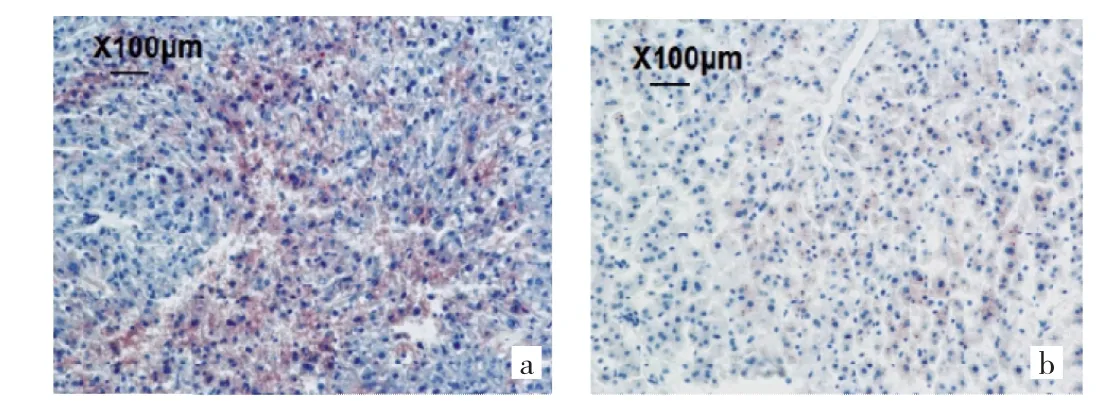
图1 两组子代雄鼠肝脏组织病理变化[a:普通母鼠子代鼠(CM-O)组;b:肥胖母鼠子代鼠(OM-O)组;油红O染色,×100]
2.2 两组子代雄鼠血清脂代谢指标比较 两组子代雄鼠血清TC、TG、HDL、LDL水平比较差异均无统计学意义(均 P>0.05),见表 2。
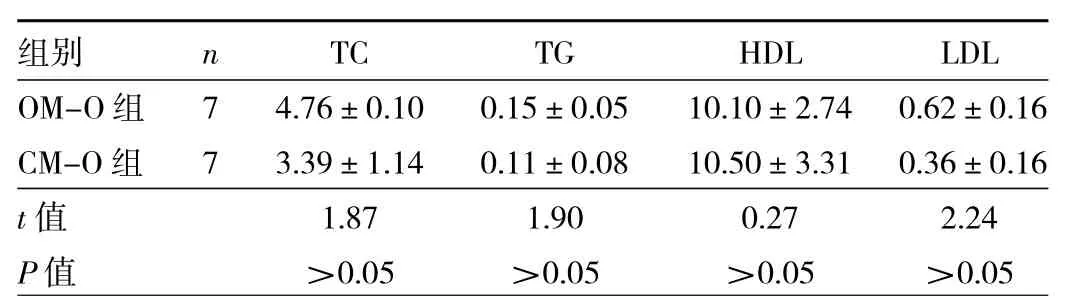
表2 两组子代雄鼠血清脂代谢指标比较(mmol/L)
2.3 两组子代雄鼠肝脏自噬相关基因mRNA和蛋白表达水平比较 与CM-O组比较,OM-O组Beclin-1、LC3B mRNA表达水平均上调,p62 mRNA表达水平下调,差异均有统计学意义(均P<0.05);而两组子代雄鼠ATG3、ATG5、ATG12 mRNA表达水平比较差异均无统计学意义(均P>0.05),见表3。与CM-O组比较,OM-O组 Beclin-1、ATG12、LC3B蛋白表达水平均上调,p62蛋白表达水平下调,差异均有统计学意义(均P<0.05);而两组子代雄鼠ATG3、ATG5蛋白表达水平比较差异均无统计学意义(均P>0.05),见图2和表4。
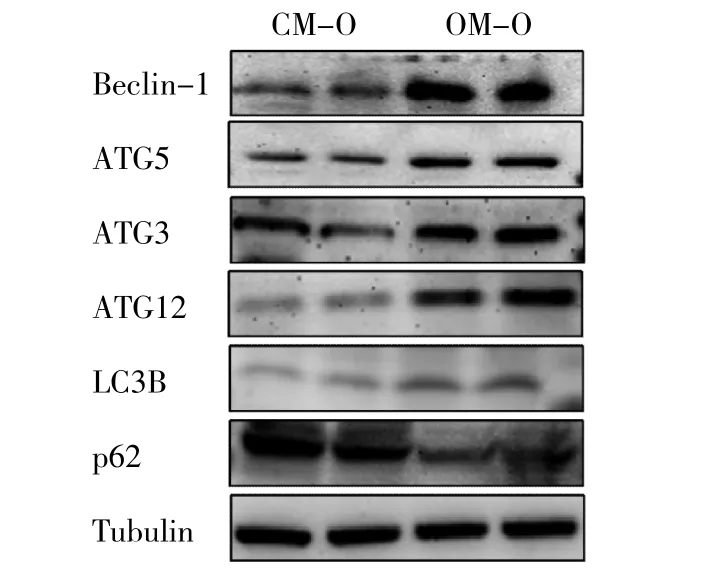
图2 两组子代雄鼠肝脏自噬相关基因蛋白表达的电泳图(肥胖母鼠子代鼠为OM-O;普通母鼠子代鼠为CM-O;自噬相关基因3为ATG3;自噬相关基因5为ATG5;自噬相关基因12为ATG12;轻链3B为LC3B;Tubulin为微管蛋白)

表3 两组子代雄鼠肝脏自噬相关基因mRNA表达水平比较

表4 两组子代雄鼠肝脏自噬相关基因蛋白表达水平比较
2.4 两组子代雄鼠肝脏组织中LC3B含量比较 两组肝脏组织免疫组化染色结果见图3(插页)。OM-O组LC3B的 IOD值为 2.43±1.02,高于 CM-O组的0.70±0.20,差异有统计学意义(Z=2.88,P<0.05)。

图3 两组小鼠肝脏中轻链3B(LC3B)免疫组化染色结果[a:普通母鼠子代鼠(CM-O)组;b:肥胖母鼠子代鼠(OM-O)组;×100]
3 讨论
母代肥胖或早期营养过剩对子代代谢影响深远。既往研究发现孕前肥胖导致的宫内环境改变或母代哺乳期高脂饮食会导致子代肥胖、脂代谢异常[8-9]。目前代谢异常的研究都集中于成年期,很少有研究关注到幼年的代谢改变。儿童早期健康状况往往是其成年后健康的重要基础,例如临床研究发现儿童早期肥胖或超重会导致成年后更容易出现NAFLD和ALT异常[10],儿童青春期血压变化将影响成年后的人体代谢[11],因此关注早期子代代谢改变将对预防成年后的疾病有重要意义。本研究选择4周龄子代雄鼠进行研究,结果发现两组子代雄鼠血清脂代谢指标比较差异均无统计学意义,但在哺乳期高脂饮食的母乳喂养下,两组子代雄鼠肝脏内出现了脂质沉积。这也与之前的临床研究:哺乳期母代高脂饮食容易导致子代青年期NAFLD的形成[6]相一致。然而,本研究发现,与CM-O组比较,OM-O组肝脏脂沉积等代谢异常表现较轻。进一步的研究发现,OM-O组肝脏中自噬相关基因Beclin-1、LC3B mRNA表达水平均上调,p62 mRNA表达水平下调;在蛋白层面的验证下,本研究发现Beclin-1及LC3B蛋白表达水平均上调,p62蛋白表达水平下调,结果表明与CM-O组相比,OM-O组子代雄鼠肝脏内自噬表达增加。Liu等[12]发现高脂喂养的SD大鼠,肝脏脂肪变性前期自噬会增加,而晚期脂肪变性加重同时会导致自噬降低,这一结论与本研究结果一致。
自噬不仅能维持体内细胞的平衡状态及正常功能,还可以作为一种重要的脂质代谢途径[13]。动物研究发现抑制自噬可以导致饥饿小鼠胚胎成纤维细胞里的脂滴数量和尺寸更大[14],在使用氯喹(自噬抑制剂)后,自噬表达降低加重了脂肪变性。而当使用雷帕霉素(自噬激活剂)时,激活后的自噬可显著减轻肝细胞中的脂质沉积[12]。Devarajan等[7]通过对母代孕期限饲喂养后,发现子代小鼠肝脏自噬表达在成年期出现下降而形成脂代谢异常,但在老年期会因为营养敏感记忆出现肝脏的脂代谢改善,这表明母代的营养状况可以通过记忆的方式传递给子代,从而激发子代的自我保护机制。本研究发现母代肥胖的子代雄鼠在早期虽然没有明显出现肝脏脂质改变,但自噬出现增加,这可以认为是一种保护机制,母代肥胖对子代雄鼠肝脏脂代谢稳态的影响可能与肥胖母代早期的营养记忆有关,这一记忆通过遗传传递给后代,使子代早期激活了自噬调节,对自身肝脏脂代谢形成代偿作用。
综上所述,本研究发现母代肥胖对其子代肝脂代谢影响可能与自噬在其中发挥了一定的积极作用有关。当然,本研究还存在一定的不足:(1)未通过调节自噬的表达来进一步明确自噬与子代雄鼠肝脏脂质改变之间的相关性;(2)未对这一变化的发生机制进行深入探究,今后将进一步开展后续工作。
