植物修复土壤重金属污染中外源物质的影响机制和应用研究进展
2021-05-10史广宇余志强施维林
史广宇 ,余志强 ,施维林
1. 苏州科技大学环境科学与工程学院,江苏 苏州 215009;2. 佛山市南海区苏科大环境研究院,广东 佛山 528225
随着我国国民经济的快速发展,工业化、城市化及农业现代化带来的土壤污染问题日益突显。重金属是一种持久性污染物,随冶炼、制革、电镀和印染等行业产生的废水进入土壤中,并随食物链在人体内蓄积,间接危害人类的身体健康。
《中国耕地地球化学调查报告 (2015年)》在2000—2015年期间对全国0.924×108hm2耕地土壤重金属污染状况进行了调查,结果显示土壤环境状况总体不容乐观,土壤重金属污染问题较为突出;重金属超标耕地点位比例占到了8.2%,多数地区以Cd、Ni、As等有毒元素复合污染为特征(中国地质调查局,2015)。从污染分布来看,南方污染形势更加严峻,尤其是长江三角洲、珠江三角洲等经济发达区域,土壤重金属污染已致使大片农田基本丧失生产力,对农产品的质量和产量造成了严重影响。由于这些重金属污染物的急性毒性,迫切需要开发低成本、高效和可持续的方法,将它们从环境中去除或转化为毒性较小的形式。植物修复技术就是这样一种方法,不仅廉价高效,还可以在修复时改善土壤肥力,有益于污染治理后的生态恢复。但在植物富集土壤重金属的过程中,植物通常也会受到重金属的毒性影响,只有少数被称为“超累积植物”的物种具有独特的能力,能够在几乎不受毒性影响的情况下积累极高量的重金属。
目前已发现的超富集植物大约有500多种,但植物修复技术在实践应用中的推广仍然受到诸多方面的制约,如植物生长速度缓慢、生长区域性和土壤中重金属生物利用度低等(Hemen,2011)。研究还发现,一些超富集植物可以同时富集多种重金属,而有些超富集植物只能针对某一种重金属具有富集作用,甚至在一些复合污染条件下无法生存(杨越晴等,2019)。为了解决植物修复技术在实际应用中的局限性问题,重金属污染土壤的植物修复强化技术逐渐成为当下研究热点。一些研究表明,一些外源物质的加入具有改变土壤重金属形态、缓解重金属对植物生长的抑制效应和刺激抗逆基因的表达等多种作用,通过提高重金属生物利用度、促进植物生长和缓解植物重金属毒性 3种机制影响植物对某些重金属的富集(闫研等,2009;曾小英等,2016;袁金玮等,2019;张盛楠等,2020)。并且其中一些物质的成本低廉、对环境几乎无毒无害,这对植物修复技术的应用与实践具有重要意义。植物修复效率与重金属在植物体内的积累分布及植物自身的抗性机理等因素密切相关,本文在解析这些因素的基础上,根据作用机制的不同,针对性地阐述了几种外源物质对植物积累重金属的影响,旨在为今后环境重金属污染的治理研究提供一定的参考。
1 重金属对植物的毒害效应及植物解毒机制
1.1 重金属在植物体内的积累与分布
对于植物修复而言,重金属在植物体内的分布要求因修复技术的不同而不同。例如植物提取更注重重金属在植物地上部中的累积量,而根际过滤则侧重于植物根系对重金属的累积。因此,研究重金属在植物中的积累分布机制对植物修复技术的实践应用十分有意义。研究表明,重金属在植物体内的分布因植物种类、组织及金属种类的不同而不同。Tang et al.(2017)研究了Pb胁迫对一年生旱柳(S.matsudana)枝条的生理生化影响,结果表明Pb处理诱导了旱柳枝条对Pb的浓度依赖性积累,95%以上的Pb分布在根中,各组织中Pb含量顺序依次为根>茎>嫩枝>叶。Angelova et al.(2004)指出在工业重金属污染区生长的亚麻(Linum usitatissimumL.)、苎麻(Boehmeria niveaL.)和棉花(Gossypiumspp.)各组织中重金属积累量依次为:根>茎>叶>籽,棉花中为:叶>籽>根>茎。而重金属在亚细胞水平上的分布主要与重金属的种类有关,Pb、Zn、Cu、Cd主要分布于植物细胞壁中,如生长在铅锌尾矿污染区中的3种菊科植物野艾蒿(Artemisia lavandulaefolia)、胜红蓟(Ageratum conyzoidesL.)和野茼蒿(Crassocephalum crepidioides),Cd、Pb、Zn在根、茎、叶中主要分布于细胞壁(31.2%—71.7%)和液泡的可溶性组分中(17.7%—54.7%),少量存在于细胞器组分(3.4%—22.1%)。并且3种植物对3种重金属均表现出良好的适应性和耐性,超过了一般植物体内的正常重金属含量。其中野茼蒿对Cd的富集特征更是达到了Cd超富集植物的标准,这可能与其细胞壁容纳了大部分的重金属有关(占总量的50.9%—71.1%)(朱光旭等,2017)。重金属在植物体内的积累分布机制十分复杂,不同重金属在不同植物及其器官、组织乃至细胞中的积累分布都有所差异。但值得肯定的是,细胞壁对重金属的固定、液泡对重金属的区隔化是重金属耐性植物普遍具有的细胞解毒机制。
1.2 重金属对植物的毒害机理
许多重金属是植物生长发育必需的矿质营养元素,如Cu、Zn、Mn等,可以直接或间接的被植物吸收利用,在植物生长发育过程中发挥重要作用。然而,当环境中重金属含量超过阈值时,就会对植物产生一定的毒性作用;轻则植物代谢过程紊乱、生长速率减缓,重则导致植物枯萎死亡。
有学者研究认为,重金属对植物的毒性可能有以下两种生物学途径:一是大量的重金属离子进入植物内干扰了离子间原有的平衡系统,造成正常离子的吸收、运输、渗透和调节等方面的障碍,从而使代谢过程紊乱。二是较多的重金属离子进入植物体内后,不仅与核酸、蛋白质和酶等大分子物质结合,还可以直接取代某些特定元素功能所必需的酶和蛋白质,使其变性或活性降低(Zhang,1997)。例如,Ni能引起脯氨酸含量的剧烈变化,破坏细胞膜稳定,导致细胞渗透失衡(张义贤等,2006);Cd和硝酸还原酶中巯基有很高的亲合性,能破坏酶的活性(陈愚等,1998);Pb离子能诱导生物大分子如核酸和蛋白质构象的改变,在分子水平上造成DNA的损伤与甲基化(Surgun-Acar,2018;丁国华等,2017)。此外,也有学者认为与其它形式的氧化胁迫类似,重金属胁迫也会降低植物体内一些抗氧化酶(SOD、POD、CAT等)的活性(罗立新等,1998a),导致大量的活性氧自由基(ROS)产生,当植物体内重金属诱导的ROS含量超过植物所能承受的临界点时,蛋白质和核酸等主要生物大分子会受到自由基引起的膜脂过氧化的伤害,是造成植物伤害的重要原因之一(Luna et al.,1994)。罗立新等(1998b)推测 ROS代谢失衡引起的膜脂过氧化在小麦(Triticum aestivumL.)Cd损伤过程中起主要作用,这与张义贤等(2006)的研究结果相吻合。
综上所述,重金属对植物的毒害作用主要是以直接或其诱导产物间接的形式,对植物生理代谢平衡、膜质、蛋白质和DNA等的损伤等实现。虽然目前重金属对植物的毒害机理研究已有很多,但观点并不一致,加之同一种重金属元素对不同植物的影响也是有所差异,是否存在新的毒害机理,有待进一步研究。
1.3 植物对重金属的解毒机制
关于植物对重金属的解毒机制,较为普遍的看法有以下两种(Baker,2010):(1)避性机制,植物通过某种外部机制抑制其从环境中直接吸收重金属,从而免受伤害;(2)耐性机制,植物体内具有某些特定的生理机制,能够直接清除重金属及其诱导产生的毒害物质,从而缓解重金属的毒害效应。而且这两条途径并不相互排斥,往往能统一作用于同个植物上。
在避性机制下,植物通过限制重金属离子跨膜运输、体外分泌物络合等方式减少对重金属的吸收,提高其在重金属胁迫下的生存能力,这种情况下植物体内的重金属含量并不高。植物的耐性机制则是通过自身抗氧化酶和非酶抗氧化剂所组成的抗氧化系统对 ROS的去除、金属硫蛋白(Metallothionein,简称为 MTs)和植物螯合肽(Phytochelatin,简称为PCs)对重金属离子的络合以及细胞壁、液泡的区隔化作用等多种途径减轻重金属对植物的伤害,使植物能够在重金属高污染环境下生存。而植物对重金属的富集水平与其生长环境的重金属浓度具有一定的相关性已被人们所知,即耐性高的植物在重金属高污染环境中能够富集更多重金属(Ray et al.,1976)。这意味着提高植物重金属耐性可以强化植物对重金属的提取作用,甚至可能使某些植物的重金属富集系数达到超富集植物标准(Soudek et al.,2016)。此外,还有研究认为一些耐性植物通过在根部积累大量的重金属,限制向地上部分的运输的方式来保护地上部分免受损害,从而提高植物重金属耐性。例如,在重金属污染环境中生长的印度芥菜(Brassica juncea)根中重金属离子的含量明显高于地上部分(Salt et al.,1995);受 Zn、Pb、Cd污染的芦苇(Phragmites australis(Cav.) Trin. ex Steud.),其根部这些离子的含量比地上部分高10倍以上(Ye et al.,2006)。
2 外源物质对植物积累重金属的影响
重金属胁迫下植物抗氧化系统、重金属积累及分布机制能保护植株免受伤害,但是植物的这种自我保护机制也有一定的局限性。通过引入外源物质,如螯合剂和表面活性剂,或多胺、水杨酸、硝普钠、硒这类具有生长调节功能的物质,针对其如何强化植物吸收转运重金属及诱导自身的解毒机制,提高植物对重金属的生理耐性,这对植物修复效率的提高具有非常重要的理论价值,同时对植物修复工程技术的实践具有指导意义。
2.1 螯合剂和表面活性剂
在治理某些复杂土质重金属污染土壤过程中,土壤中的重金属和土壤有机质或无机盐紧密结合,大部分与固相牢固结合的重金属难以直接去除,若直接采用植物修复技术对土壤进行修复,效果往往不尽人意(Vigliotta et al.,2016)。针对这种情况,一类具有活化重金属和提高重金属生物利用性作用的化学强化剂诞生了。在进行植物修复时向土壤中添加化学强化剂,使重金属由不溶态变为可溶态,从而使植物对重金属的吸收、积累和转运能力大大增强(Ali et al.,2013);也可以认为强化剂的添加使环境中的重金属以较低毒性的金属螯合物形态存在,规避了植物的避性机制,促进了植物对土壤中重金属离子的吸收。常见的化学强化剂应用简单、广谱性和修复周期短等特点,已引起国内外学者广泛的关注。
2.1.1 螯合剂
螯合剂强化植物修复的原理是向土壤中投加螯合剂,使土壤中的重金属与螯合剂发生螯合作用,改变重金属在土壤中赋存的形态,形成稳定的环状结构的金属螯合物。并且这种螯合物在土壤中具有较好的溶解性,使重金属的生物有效性大大增强,从而增强植物对靶向重金属的吸收。目前各种人工和天然螯合剂,以及一些天然小分子有机酸类物质已广泛应用于重金属污染土壤的修复(Yan et al.,2017)。应用较多的人工螯合剂有 EDTA、EGTA、HEDTA、CDTA、DTPA等,其特点是施用量少、螯合能力强且形成的螯合物拥有极佳的稳定性。然而其成本较高,对植物还具有一定毒性,且在自然环境中难以降解、容易导致一些非靶向重金属元素扩散到周围环境甚至通过雨水淋滤进入地下水,造成环境二次污染(Han et al.,2019)。因此,人工螯合剂在环境污染中的应用和推广受到了限制。
从生物中提取的天然螯合剂 EDDS、NTA、GLDA等拥有良好的金属螯合能力,同时具有可快速生物降解特性,对环境的影响很小。常用的柠檬酸、草酸、苹果酸等天然小分子有机酸类物质对植物无毒且易被降解,除了能螯合金属离子外,也可降低土壤pH值,增强重金属在土壤中的流动性。韩廿等(2019)报道了不同螯合剂(NTA、EDDS、EGTA和EDTA)对油葵(Helianthus annuusL.)修复Cd-As复合污染农田土壤的影响,结果表明施用 NTA、EDDS、EGTA、EDTA均促进了油葵对Cd的吸收。与对照组相比,油葵花盘中的Cd含量分别提高了30.2%、41.9%、55.1%和43.3%,总Cd积累量分别提高了32.8%、40.5%、45.3%和41.6%;根系As含量分别提高了23.6%、15.6%、18.1%、和15.4%。陈立等(2017)对比了不同浓度EDDS、草酸和柠檬酸协同向日葵(Helianthus annuus)修复Cd污染土壤效果,结果发现3种螯合剂均提高了向日葵对Cd富集和转运能力,以EDDS质量摩尔浓度为5 mmol·kg-1时效果最佳,向日葵地上部、地下部Cd富集系数分别达到了6.18、5.94,是对照的4.52、2.34倍;且EDDS和柠檬酸处理还在一定程度上促进了Cd从向日葵地下部向地上部的转运。整体而言,可生物降解的天然螯合剂的重金属螯合能力有待加强,对植物提取重金属的促进效应不够明显。但由于其环境友好的特性,在实际应用过程中拥有很好的前景。因此,开发螯合能力更强,降解率更高的天然螯合剂是螯合剂未来的研究趋势。
2.1.2 表面活性剂
表面活性剂是指加入少量就能改变介质体系界面状态和性质的物质,具有增溶、增流等特性,可以使重金属以络合物的形态存在于土壤溶液中。这些形态极易流动,使重金属在土壤中的流动性大大提高,进而促进植物对重金属的吸收,实现提高植物修复效率的目的(刘江红等,2019)。Gadelle et al.(2001)在水培条件下用非离子型Tween-80、阳离子型CTAB、阴离子型LAs等3类表面活性剂处理Cd胁迫下的小麦,显著提高了小麦叶片中Cd的含量,分别为 3.725、5.125、4.594 mg·kg-1,是对照的 3.09、4.25、3.81倍。混合表面活性剂 SDS-Span80和SDS-Span80-GSH显著促进了蜈蚣草(Pteris vittataL.)对As的提取,总As含量分别提高了31.7%和94.2%,土壤有效态As提高了73.4%和81.4%(Xiang et al.,2020)。说明表面活性剂的施用活化了土壤中的重金属,增加了重金属的移动性和生物可利用性,从而促进植物对重金属的提取。
也有学者认为,表面活性剂促进植物吸收重金属的作用机理可能与表面活性剂破坏细胞膜透性有关(陈玉成等,2004)。表面活性剂是一种可溶性、两亲性的特殊脂类化合物,不同于构成生物膜成分的不溶性和具膨胀性的脂类化合物,它在水中有较高的单体溶解度,其两亲性使之能与膜中成分的亲水和亲脂基团相互作用,从而改变膜的结构和透性,促使重金属在植物体内累积。例如,CTAB(十六烷基三甲基溴化铵)可改变细胞表面性质和细胞膜渗透性,细胞表面疏水性从3%增加到14%;Zeta电位从-14.5 mV增加到-10.2 mV;胞内ATP含量从 28.599 μg·g-1下降到 9.737 μg·g-1,胞外 ATP含量从33.051 μg·g-1增加到82.809 μg·g-1;K+和 Ca2+浓度分别提高到 3.9 mg·L-1和 2.1 mg·L-1,扫描电镜图像显示细胞表面有明显的形态变化和破坏(Wang et al.,2019)。此外,与其他化学强化剂类似,表面活性剂对植物的生长表现出一定的抑制性,并且其本身在环境中难以降解,易造成二次污染。近年来,高效廉价、可生物降解的生物表面活性剂在重金属污染修复领域越来越受到关注,其中研究较多的生物表面活性剂有鼠李糖脂、皂角苷、槐糖脂等。由于其本身无毒或低毒性,不会对植物生长产生不利影响,也不会改变土壤结构和物化性质,被认为是一种应用前景良好的重金属污染植物修复强化措施(卢宁川等,2009)。CTAB、鼠李糖脂和皂角苷处理Pb、Zn胁迫下的超富集植物小花南芥(Arabis alpinaL. var. parviflora Franch),均显著促进小花南芥对Pb和Zn的富集,并且位移系数和富集系数都大于1;只有CTAB处理质量浓度为7.5 g·L-1时,小花南芥叶长、冠幅、根长及地下部和地上部生物量有不同程度的下降,分别为对照的 0.61—0.88倍(王吉秀等,2010)。5.0 g·L-1的鼠李糖脂能够促进钒矿污染土壤中紫穗槐(AmorphafruticosaL.)的生长,地上部生物量、根部生物量及根系活力分别较对照组增加了48.38%、13.22%和38.97%;并使土壤中碳酸盐结合态和残渣态重金属向可交换态、铁锰氧化物结合态、有机结合态转化,提高了土壤中 V和Cr的生物有效性,显著促进紫穗槐对V和Cr的吸收及转运(李昊等,2020)。
2.2 新型植物激素
植物激素是植物自身产生的一种内源生长调节物质,微量浓度就足以对植物生长代谢活动产生影响。其在植物体内某些部位合成、可被运送到作用部位,且对植物具有特殊的生长调节功能,对植物的生长发育起到了重要作用。植物激素可以利用其独特的生理作用改变重金属胁迫下植物对外界营养物质、水分的需求,增强植物对逆境的防御能力,从而达到提高植物提取重金属能力的目的。此外,有研究显示植物激素对一些超富集植物发芽率低、生长缓慢和生物量低等问题有明显改善作用(Ji et al.,2015;Du et al.,2015;He et al.,2015;Fässler et al.,2010)。因此,利用植物激素强化植物修复重金属污染具有积极意义。目前普遍认为的植物激素主要有生长素、细胞分裂素、赤霉素、乙烯、脱落酸、油菜素甾醇六大类。近年来,研究发现被认为是新型植物激素的多胺、水杨酸具有调控植物积累重金属的重要作用,引起人们的关注。
2.2.1 多胺
多胺(PAs)是由氨基酸衍生而来的低分子质量有机聚合物。在结构上,PAs是含有两个或多个氨基的脂肪链,现在人们普遍认为 PAs是类似cAMP(环磷酸腺苷)的“第二信使”或新型激素。植物中普遍存在的 PAs主要有腐胺(Put)、亚精胺(Spd)、精胺(Spm),它们广泛分布于植物组织中,并参与了植物的各种重要生理过程(Alcázar et al.,2006;Tiburcio et al.,2018)。目前关于其与植物生长发育、延缓衰老和逆境响应之间的关系的研究较多,尤其是关于缓解植物重金属毒害及其作用机理等方面。研究表明,PAs提高植物重金属抗性可能有两条作用途径,一是由于PAs是具有阳离子性质的低分子有机化合物,能够与重金属发生螯合作用,从而抑制金属离子进入细胞,也可以与带负电的核酸、蛋白质和细胞膜的磷脂头部分等结合形成结合态 PAs,具有调节阴阳离子平衡、生物大分子合成、稳定蛋白和细胞膜结构的作用(Rady et al.,2015;Zepeda-Jazo et al.,2018;李霞等,2018)。申璐等(2014)通过盆栽实验发现,外源1.0 mmol·L-1Spd对茶树(Camellia sinensis(L.) O. Kuntze var.Longjing43)高浓度Pb胁迫具有明显的缓解作用,茶树叶片可溶性蛋白含量和叶绿素含量明显上升,脯氨酸和丙二醛的含量下降。此外,还显著较低了叶片的相对电导率。这说明外源Spd有效阻止了Pb胁迫对茶树生理生化造成的负面效应,促进了功能蛋白的合成,抑制了代谢平衡紊乱,并在一定程度上保护了叶片细胞膜的稳定性,提高了植物重金属耐性。二是PAs能够直接的或间接的清除重金属诱导产生的大量ROS,Drolet et al.(1986)报道了PAs能够有效地清除由衰老的微粒体膜所产生的超氧阴离子(O2-·)和化学和酶系统产生的自由基,其中Spd、Spm的清除效率大于 Put。Hg胁迫下生长的荇菜(Nymphoides peltatum),SOD、CAT、APX、POD活性发生了显著变化,SOD、CAT、APX活性大幅降低,POD活性却大大上升,这可能与 POD的双重作用有关,它既可以与SOD、CAT和APX一样消除植物体内的ROS,也能促进ROS的产生。外施低浓度的 Spm明显改善了这一现象,提高了SOD、CAT、APX的活性,极显著地降低POD活性,从而降低了植物体内的ROS水平,缓解了重金属诱导的氧化胁迫(王学等,2008)。Put浸种预处理大大缓解了 Cd、Pb对小麦的胁迫,提高了小麦幼苗的生长量及SOD、CAT、POD等抗氧化酶的活性,DNA链的断裂也明显减少(Taie et al.,2019)。
目前有关 PAs强化植物修复的报道极少,但PAs可以影响重金属在植物体内的积累已经得到证明。Shevyakova et al.(2011)报道了外施Put显著降低了Ni对甘蓝型油菜(Brassica napusL.)的毒性作用,并促进了油菜对 Ni的积累。袁菊红等(2011)通过盆栽实验考察了外源 Spd对锰超富集植物美洲商陆(Phytolacca americanaL.)富集城市污泥中Cu、Zn、Cd、Hg的影响,结果表明外施1 mmol·L-1Spd能够明显促进美洲商陆植株生长及其生物量的增加,对Zn、Hg、Cd的富集和转运系数也有不同程度的提高。在Tang et al.(2019)的研究中,Spd的施用显著促进了 Cd在一年生旱柳枝条根、茎中的积累,并加强了细胞对重金属的区隔化作用,增加了Cd在细胞壁中的分布。但也有报道称PAs增强植物重金属耐性的同时,也会抑制植物对重金属的吸收和积累(Nahar et al.,2016)。这可能与PAs的施加浓度、时期及重金属对植物的作用时间有关,因此今后有必要进一步研究PAs在植物中作用的分子机理,探索外源PAs调控的具体水平、浓度与最佳时期,从而更好地了解PAs在植物逆境响应中的生理功能,在实际修复过程中利用PAs的生物强化功能实现增强植物修复的目的。
2.2.2 水杨酸
水杨酸(SA)是植物体内普遍存在的一种信号小分子酚类物质,对植物生长发育具有多种生理调节效应,在植物抗生物和非生物胁迫中扮演重要角色。关于SA在植物抗重金属胁迫的作用,已经有了一些研究,但其具体作用机制尚不明确。Krantev et al.(2008)认为SA的作用可能是直接的,可作为一种抗氧化剂直接清除活性氧类物质,减轻重金属诱导的氧化胁迫对植物的伤害。而更多的研究则是认为 SA缓解重金属的毒害作用是通过激活抗氧化反应来调节植物体内的氧化还原平衡,或通过增加H2O2的含量来触发信号转导网络,或者通过改变植物中钾、镁等矿质元素的含量和分布来间接作用的(Guo et al.,2007)。但也有报道称SA在缓解重金属胁迫中并不起作用,如在Zawoznik et al.(2007)的研究中,野生型拟南芥(Arabidopsis thaliana)Cd胁迫5 d后开始出现毒害症状,叶绿素含量和抗氧化保护酶活性下降,H2O2和内源SA的含量显著上升。而SA缺失的转基因拟南芥受Cd胁迫的影响并不显著,但可以推断一定水平的内源SA是作为保证或加强Cd诱导的氧化伤害的必要信号分子。
SA对植物吸收积累重金属的影响主要依赖于植物种类和重金属胁迫水平。喷施不同浓度SA对番茄(Solanum lycopersicumL.)幼苗进行预处理,大大地提高了幼苗在Cd胁迫环境中的生存能力,主要表现为减缓Cd诱导的氧化胁迫和植株生长量的增加,但同时也降低了番茄幼苗对Cd吸收。与对照组相比,番茄地上部Cd积累降低了 24.4%—58.3%,根部Cd积累降低了50.5%—75.5%(王小红等,2019)。这可能是由于SA的生长调节作用提高了植物对大量元素和微量元素的吸收,促进Ca2+、Mg2+等离子的内流,占用了离子通道,从而阻挠了 Cd2+进入植物根系抑制根的生长。而对于Cd污染土壤中生长的大豆幼苗,当Cd胁迫水平3 mg·kg-1时,外源 SA处理促进了大豆(Glycine max)幼苗对Cd的吸收,Cd胁迫水平为6 mg·kg-1时,添加的SA对Cd的吸收并没有明显影响(Drazic et al.,2005),这表明SA缓解Cd毒害作用可能并不是通过阻止植物对 Cd的吸收实现的。Khan et al.(2018a)的研究也证实了这点,其研究发现喷施SA处理显著促进了鹰嘴豆(Cicer arietinumL.),对Cd、Pb、Ni的提取及Ni从根部向叶片中的转运,增强了其修复重金属污染土壤的潜力。此外,他们对干旱与重金属复合胁迫下的向日葵也进行了相同研究,并得到了一致的结果(Khan et al.,2018b)。这可能是由于SA通过一些特殊的机制降低了重金属毒性,如与重金属结合成复合物导致重金属钝化,或是通过小泡吞噬重金属离子等途径。现有的研究均证实了SA可以某种程度上促进或抑制植物对重金属的吸收和积累,但其中的一些机理探究仍停留在猜测和假说阶段,这意味着将来需要进一步用生物化学的、分子生物学和染色体甚至遗传学的方法关键对SA在植物抵御生物的和非生物的胁迫中的作用进行全方位的深入研究。加之SA在不同种类植物或同种植物的不同基因型中诱导抗重金属胁迫的机制可能都会有所不同,甚至相互矛盾,其中有很多问题仍待解决。
2.3 其他外源物质
除了上述外源物质外,关于缓解植物重金属胁迫的物质当今报道较多的还有硝普钠和硒,对重金属在植物体内的积累分布同样表现出一定的调控作用。
2.3.1 硝普钠
蠕虫状链是Porod和Kratky在1949年提出的概念[2],因此蠕虫状链又称Porod-Kratky链.无论统计单元数是多少都可以用这种链模型进行处理,因此蠕虫状链既可以描述柔性链,也可以描述刚性或半刚性高分子链,还可以描述分子量较低时的齐聚物柔性链.
硝普钠(SNP)是一种在水中溶解时能够释放NO的化合物,因此被普遍当作外源NO供体。NO是一种广泛存在于生物体内的自由基气体信号分子,可通过得失电子形成自由基(NO·),亚硝酰离子(NO+)、硝酰基阴离子(NO-)快速透过细胞膜发挥作用,参与多种植物生理反应(Wojtaszek,2000)。重金属胁迫条件下,NO除了可以清除脂质自由基和超氧阴离子调节植物体内物质代谢,还可以穿梭于细胞内外传递胁迫信息,加强植物对逆境的响应能力。NO能够诱导重金属胁迫下的植株表达更多的PCs和MTs等含巯基肽类物质络合过多的重金属离子,降低其生物毒性;还可以通过调节膜脂过氧化、抗氧化系统、渗透调节物质等多方面的作用来缓解重金属胁迫抑制作用(陈银萍等,2015;王逸筠等,2018)。也有学者认为,NO缓解植物重金属伤害是在分子水平上实现的,它可以通过翻译后修饰(PTMs,如 S-亚硝基化和硝化作用)直接改变蛋白质,调控基因表达与细胞内生理过程,进而影响植物激素的合成代谢、信号传导及ROS信号调节机制,减轻重金属对植物的伤害(María et al.,2016;Cui et al.,2018;Terrón-Camero et al.,2019)。
关于SNP对植物吸收重金属的影响较为复杂,已有的研究表明,不同植物或同种植物的不同基因型、SNP浓度、重金属胁迫水平等都是SNP影响植物积累重金属的重要因素。应用1—50 mg·L-1SNP可以有效控制水稻(Oryza sativaL.)对Hg的积累,通过在水稻生长各时期喷洒适当浓度的SNP溶液,使生产的稻米的Hg含量均降低30%—50%(朱诚等,2015)。朱涵毅等(2013)研究了两种水稻品种籼稻品种ZF802和粳稻品种ZH11在外源SNP介导下对Cd的积累影响,结果表明1.5 mmol·L-1SNP对两种水稻幼苗的Cd毒害的缓解效果最明显,但对于Cd积累存在一定的基因型差异,施加外源SNP后籼稻品种ZF802,地上部Cd含量下降,根部含量增加,总Cd含量增加;粳稻品种ZH11却相反,地上部Cd含量略有升高,在根部Cd含量却降低,总 Cd含量减少。而在冉烈等(2012)研究中,不同浓度外源 SNP均促进了超富集植物东南景天(Sedum alfrediiHance)对Cd的吸收与累积,并且东南景天的生物量、不同部位的Cd含量和累积量随着外源SNP浓度的增加而增加,累积量依次为叶片>茎>根。另外在 10 μmol·L-1和 100 μmol·L-1Cd胁迫下,外源SNP介导下东南景天的Cd吸收和转运速率也存在着不同程度的提高,最高分别可达79.8%和81.9%,但这种促进效应依赖于SNP浓度和Cd胁迫水平。
研究NO的具体功能和一般作用,例如它参与重金属在不同植物中的积累和分布,可提高重金属吸收率较低作物的生产或强化重金属复合污染土壤的植物修复(Terrón-Camero et al.,2019)。尽管NO参与植物对重金属胁迫的响应已经得到证实,但有关其与 ROS以及其它信号组件如激素、转录因子等之间的串扰机制研究才刚刚开始,有待更深入的研究。
2.3.2 硒
硒(Se)是维持生物体内氧化还原平衡的一种重要微量元素。对于植物而言,其虽然不是生长的必需元素,但适量的 Se可以促进植物对营养元素的吸收。植物的 Se来源是土壤中的硒酸盐(SeO42-)、亚硒酸盐(SeO32-)以及少量的有机硒化物,其增强植物抵御生物与非生物胁迫的能力(Reynolds et al.,2018)。近年来,Se被证明具有保护植物受重金属影响的能力,其与重金属的相互作用机制逐渐成为当下的研究热点。研究表明,Se能够通过多种作用途径参与植物抗重金属胁迫过程,如可以增加根部细胞壁中果胶和半纤维素的含量,使细胞壁与重金属的结合能力增强,从而阻止重金属离子向细胞内转移;根吸收的SeO32-在根部会迅速被转化为有机形式,硒氨基酸与重金属形成复合难溶解性物,增加根中 Se的保留率,抑制重金属向地上部转移;还可以刺激 O2-自发歧化成H2O2(没有SOD的催化)、直接清除O2-和OH·、调节抗氧化酶活性;同时 Se也是谷胱甘肽过氧化物酶(GR)的重要组成成分,参与谷胱甘肽的合成。因此,Se对ROS的清除也有着不可忽视的作用(Yu et al.,2018;Zhao et al.,2019;陈思蒙等,2020)。
由于 Se在维持人体免疫功能和降低疾病风险等方面的重要作用,富硒农作物的生产一直受到人们的重视。目前,作物品种筛选、硒肥施用及其他生物措施是生产富硒农产品的主要途径。Se也是“重金属的天然解毒剂”,早在1973年,Francis et al.(1974)就已发现Se与Cd存在拮抗作用,外源添加适当浓度Se可以提高植物体内Se含量,同时减少Cd在植物中的积累。而当Se浓度过高时,则会对植物表现出与重金属类似的毒性,与重金属产生协同效应,增加植物对Cd的吸收。例如,低质量分数(10—15 mg·kg-1)的Se处理能够抑制延龄草(Trillium tschonoskiiMaxim)对Cd、As、Pb、Hg的吸收,而施加较高质量分数(25 mg·kg-1)的Se能够促进延龄草对4种重金属的吸收,且延龄草没有出现毒害症状(江念等,2016)。Cd胁迫下,叶面施Se可以减轻花生(Arachis hypogaeaL.)Cd中毒的毒副作用,提高花生体内的Se浓度,抑制根、叶对Cd的吸收,降低营养器官中Cd的含量;而随着施Se浓度增高,花生叶片及根系中Cd含量也逐渐升高(卞威乐斯等,2018)。此外,Se与Cd的这种拮抗或协同作用也反映在其他植物中。低浓度的Se可显著降低小白菜(Brassica chinensisL.)地上部对于Cd的吸收,浓度过高的Se促进了小白菜地上部对 Cd的富集(刘达等,2016)。而在曾宇斌等(2017)的研究中,除了土壤 Se质量分数为2.56 mg·kg-1时加重了 Cd对大豆的毒害,表现出Se-Cd 的协同作用;在 1.56、3.56、6.56、11.56 mg·kg-1Se质量分数处理中,大豆茎、果、壳、叶中Cd的积累均随着土壤 Se质量分数的增加而减少。周健等(2017)报道了施用 Se2-与 Se4+可有效增强 Cd胁迫下小油菜(Brassica campestrisL.)的生理特性,抑制Cd在其体内的积累;而施加Se6+增强了Cd对小油菜的胁迫作用,且根部和地上部Cd含量明显增加,综上所述,Se对Cd的作用主要取决于植物类型和Se形态、浓度及添加方式等因素。
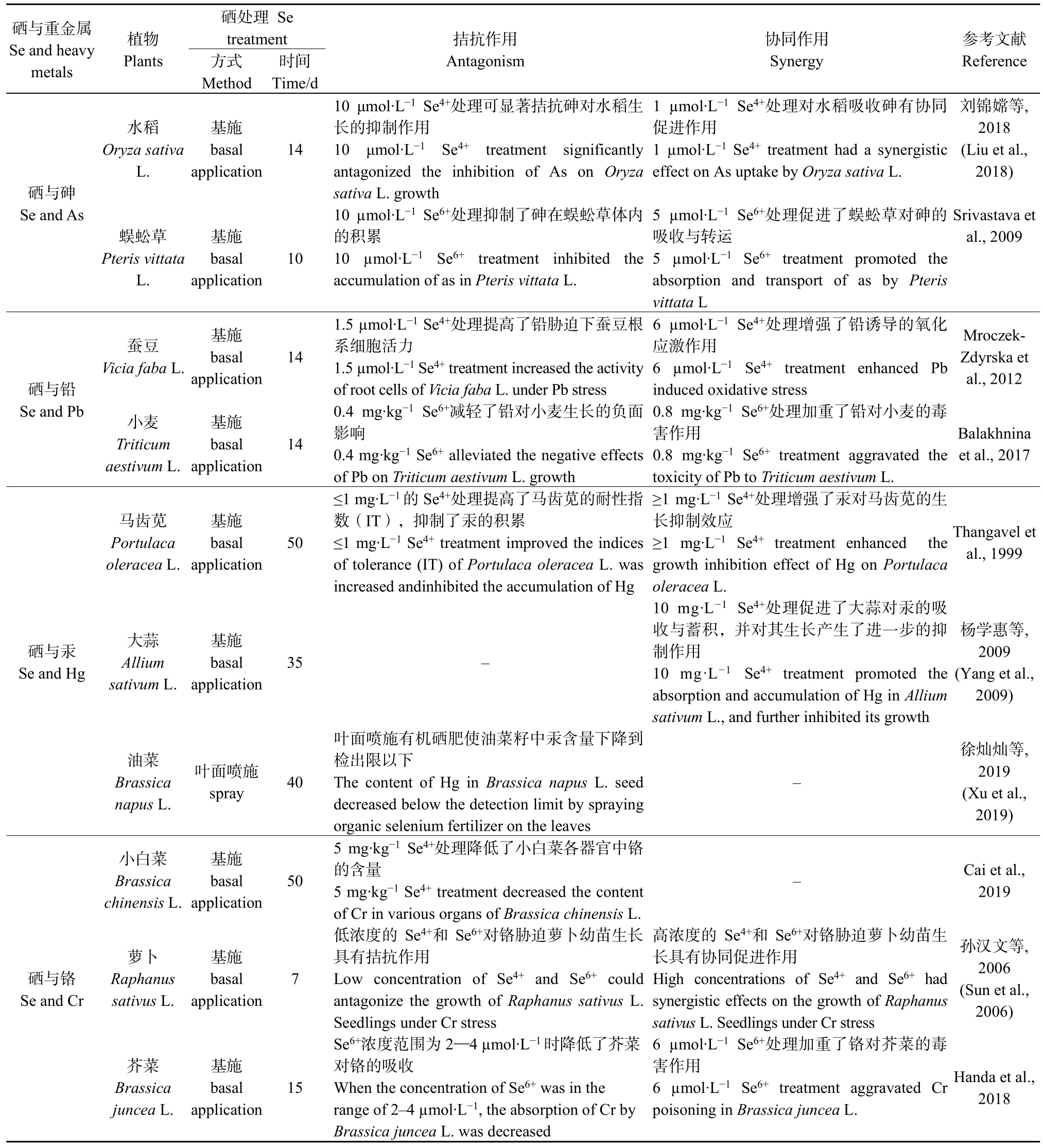
表1 植物中硒与重金属的相互作用Table 1 Interaction between selenium and heavy metals in plants
2.4 外源物质的联合应用
与施用单一外源物质相比,几种外源物质的联合使用在缓解植物重金属诱发的毒性、增强植物在高浓度重金属环境下的生存和提取能力方面具有更好的效果。植物激素及其它一些具有生长调节功能的物质能够促进植物在重金属胁迫下的生长,将其与螯合剂提高重金属生物利用度的功能相结合,可以增强植物对重金属的富集与转运能力。
Fässler et al.(2010)研究发现植物生长素IAA(吲哚-3-乙酸)和螯合剂EDDS联合应用促进了向日葵对 Pb、Zn的提取,提高了植物对重金属污染土壤的修复潜力。Sun et al.(2014)报道在激动素KN(6-糠氨基嘌呤)与 EDDS的共同作用下,Cd和 Pb从苎麻地下部向地上部的转运系数分别提高了 47%和 112%;与 IAA单独应用相比,IAA和EDDS联合应用使苎麻根系中 Cd和 Pb增加了约2.02、2.62倍。张倩等(2019)的研究表明,外源添加SNP与EDTA均可缓解Cd对黑麦草(Lolium perenneL.)的胁迫,且二者复合处理效果更为明显。与对照组相比,EDTA+SNP处理大大缓解了Cd对黑麦草的生长抑制,生物量及抗氧化胁迫能力大大提升。同时,地上部与地下部Cd含量分别提高了89.7%和30.2%,提高了黑麦草修复Cd污染土壤的潜力。也有研究表明,一些植物激素或信号分子物质之间的互作也能产生类似效应,如0.2 mmol·L-1Put和1 mmol·L-1SNP单独或联合处理绿豆(Vigna radiatacv. BARI Mung-2)幼苗,结果发现联合处理可以更明显地缓解其Cd毒害效应,降低根、茎、叶对Cd的吸收(Nahar et al.,2016)。盆栽实验表明,脱落酸(ABA)和SA的复合处理提高了高羊茅(Festuca elataKeng ex E. Alexeev)地上部Cd含量至对照的1.97倍,提高了印度芥菜地上部Cd含量至对照的1.70倍,分别达到69.2 μg·pot-1和27.8 μg·pot-1的水平。这种复合信号分子处理能够诱导植物衰老,进而调控重金属在修复植物体内的迁移与分布,提高地上部重金属的含量,而HMA3基因在这一过程中扮演着重要的作用(Zhu et al.,2020)。这预示着基于外源物质调控植物积累重金属的基因工程技术可以成为选育重金属抗性作物和超富集植物的新途径,也为土壤重金属污染及其他类型污染的植物修复研究和应用提供了新方法和新思路。
3 前景与展望
土壤中过量的重金属通过多种途径影响植物生长发育,如干扰光合作用、水分代谢、矿质元素吸收等,同时重金属也会随植物进入食物链中,对其他生物造成严重影响。由于一些外源物质在调节植物生理代谢和吸收积累重金属等方面的重要作用,在重金属污染植物修复领域受到越来越多的重视。螯合剂、表面活性剂可以通过转变土壤中重金属离子的形态,间接促进植物对特殊地质土壤中重金属的吸收,提高植物修复的效率。PAs、SA、SNP、Se这类具有调节植物生长作用的物质的应用可以有效改善重金属污染环境中植物的生长状况,刺激重金属螯合物质和清除 ROS活性物质的表达,平衡重金属诱导的代谢紊乱,调节重金属在植物体内的积累。对于超富集植物,施用其可以解决其在修复重金属污染土壤过程中面临的生长缓慢和生物量低等问题,从而提高植物对重金属离子的提取量。而对于一些重金属富集植物,施用其可以提高它们的重金属累积上限,甚至可以使其达到超富集植物水平,并且其对植物亲和性高、应用方式简单,叶面喷施或种子浸泡处理即可达到上述效果,这在植物修复工程的实践应用中占有相对优势,但其中也存在许多问题亟待解决。
外源植物生长调节物质对植物普遍具有促进生长,增强抗逆性的功能,但这种功能与其应用浓度、时期、方式及植物种类等多种因素密切相关,如何合理地控制这些因素,提高植物对重金属的耐性,减少或增加重金属在不同植物中的积累,对重金属污染区的植物修复和农业生产具有重要意义,也是目前迫切需要解决的问题。此外,关于它们缓解植物重金属毒害的具体作用机制尚不明确,已有的研究主要从植物光合作用、生理代谢、抗氧化酶活性变化等方面进行探讨,而植物对重金属胁迫的响应与植物体内的多种信号物质密切相关,如植物内源激素、Ca2+、ROS、MAPK级联和 microRNA等,这些物质相互关联、相互作用形成一个复杂庞大的信号网络,调控植物的整个生长发育过程。目前,这些外源物质与整个信号网络在植物响应重金属胁迫中的交互作用仍不清晰,其在这种交互作用中的地位,以及是否存在通过影响植物体内信号物质的合成、运输和代谢来间接调控植物生理活动的可能,这些都是不容忽视的问题。在分子水平上,它们参与调控植物耐重金属有关基因表达的背后是基因表达量的增加或是基因转录后修饰的结果尚无定论。另一方面,植物内生和根际微生物在其抵御重金属胁迫或吸收转运重金属离子扮演着不可或缺的角色,而一些外源物质(如PAs和Se)对微生物的生命活动同样具有重要的生理功能,但它们对植物的作用过程中对植物内生和根际微生物的生命活动是否同样有影响,也是很有意义的问题。综上所述,外源物质应用条件对不同种类植物吸收转运重金属的影响及其调控机理的探究,将成为重金属污染植物修复领域的研究热点。在未来的研究中需要借助更多的手段从植物内源激素及信号分子间的互作、抗逆基因表达、内生及根际微生物等不同层面上探讨这些外源物质与植物相互作用的本质,并在此基础上开展外源物质与其他植物修复强化技术的联合应用研究,以解决植物修复技术在实践化、经济化过程中遇到的难题。
