区域景观中生境特异性对昆虫多样性的影响*——以西双版纳为例
2021-05-07卢志兴高舒桐唐春英陈又清
张 翔, 卢志兴, 王 庆, 高舒桐, 唐春英, 李 巧, 陈又清**
区域景观中生境特异性对昆虫多样性的影响*——以西双版纳为例
张 翔1, 卢志兴1, 王 庆1, 高舒桐1, 唐春英2, 李 巧2, 陈又清1**
(1. 中国林业科学研究院资源昆虫研究所 昆明 650224; 2. 西南林业大学 昆明 650224)
生境特异性(habitat-specific)在维持生物多样性上发挥着重要作用。本研究旨在了解在区域景观中, 不同类型生境对不同类群昆虫多样性的贡献及群落结构差异, 从大尺度上探讨昆虫多样性分布格局和维持机制。于2019年8—9月, 在西双版纳地区利用样线调查法, 调查了保护区、次生林、人工林和农田4种生境中蝴蝶、蝽和甲虫的物种数、特有物种数, 分析了昆虫群落的生境特异性指数、群落结构相似性及物种丰富度与生态因子的关系。共采集昆虫2588头, 其中蝴蝶94种(744头), 蝽197种(1094头), 甲虫129种(750头), 保护区的昆虫物种数和特有物种数均高于其余类型生境。保护区昆虫群落的生境特异性指数实测值高于期望值, 而其余类型生境昆虫群落的生境特异性指数实测值均低于期望值; 不同类型生境间, 3个类群的昆虫群落汇总的生境特异性指数不存在显著差异(3,57=2.054), 甲虫生境特异性指数差异显著(3,55=3.478), 蝴蝶(3,38=1.504)和蝽类(3,53=1.153)生境特异性指数差异不显著。群落结构相似性分析显示, 保护区和农田的3个类群昆虫群落汇总的群落结构差异显著; 次生林和人工林的蝴蝶群落结构差异显著, 保护区和次生林的蝽类群落结构差异显著, 甲虫昆虫群落结构差异不显著。本研究还发现, 只有生境类型对昆虫群落的物种丰富度产生的影响极显著(<0.01), 而其他生态因子的影响不显著。在大尺度区域景观中, 保护区对昆虫群落的多样性影响最大, 生境类型与昆虫群落的物种丰富度密切相关, 保护较好的天然林是维持区域昆虫群落分布格局和多样性的重要机制。
区域景观; 昆虫群落生境特异性; 昆虫多样性; 群落结构; 西双版纳
人类活动正大幅度地改变生态系统结构, 特别是自然生境的转换和退化, 对全球生物多样性已经造成了严重的威胁[1-2]。生态系统功能退化、物种灭绝是当今世界面临的重要生态环境危机之一[3], 对生物多样性进行保护和管理是全球性的课题, 尤其是大尺度上, 生境结构变化导致生物多样性格局及其形成机制的改变是当前研究的热点[4-5], 确定生物的多样性格局并理解其维持机制是制定有效保护策略的前提[6-7]。
农业景观是由集约化农业用地和非农业生境组成。不同类型生境为不同类群生物提供多样化的环境条件, 提供更多的生态位, 有利于更多生物的生存栖息[8]。近年来, 以“农业集约化”为标志的农业景观格局的急剧变化, 导致了农业生态系统生物多样性下降[9-10]。昆虫作为多样性较高的生物类群[11-12], 在生态系统中扮演着不同的角色, 农业景观改变也是昆虫多样性减少的重要原因[13]。
蝴蝶、蝽和甲虫等昆虫类群是许多农业生态系统较为常见的类群, 是传粉者、植食性物种和捕食者, 对农业生态系统的功能和服务发挥着重要作用[14]。其中蝴蝶是重要的授粉昆虫, 对栖息地的寄主植物、蜜源植物和温湿度等环境因子要求较高, 并能够快速响应生境结构的变化[15]; 而蝽类在农林复合系统中, 多数为植食性物种, 主要危害农林作物, 少数为捕食性物种, 可用作生物防治的对象, 其种群和群落结构的变化可以揭示生境性质或质量的变化, 对于保障整个系统的健康具有现实意义[16]; 甲虫在农林系统中作为授粉昆虫和蛀干害虫, 由于扩散能力较差, 专一性较强, 一旦生境丧失或破碎, 一些专一性的物种很有可能消失[17]。
农业景观的植被结构、空间位置和周围景观碎片化程度都会影响本地昆虫群落的结构[18]。例如, 农业用地与非农业的自然生境以及半自然生境(草地边缘和林地等), 由于植被结构的不同, 会直接影响昆虫的繁殖地、栖息地、食物源和避难所, 对于维持昆虫多样性的作用具有一定的差异[18-20]。另一些研究也表明, 在不同类型生境间, 植物多样性是影响蝽类物种丰富度、多度和物种组成的关键因素[21], 生境中植被丰富度越高, 蝽类昆虫的多样性也随着升高[22-23]。比较土地利用类型和海拔梯度对昆虫群落特征的影响发现, 昆虫物种丰富度与土地利用类型没有显著关系, 但与海拔之间存在显著关系, 说明空间位置对昆虫多样性产生一定影响[24]。生境连通性和微生境多样性对保护甲虫多样性具有重要意义[17], 也是维持甲虫[25]和蝴蝶[15,26-27]群落稳定性的重要因子。但这些都是在小尺度上探讨昆虫对生境结构变化的研究, 而从大尺度下研究昆虫群落结构变化更有助于我们理解昆虫对生境结构变化的响应[28], 特别是针对农业生态系统中不同功能作用的昆虫[18], 以及不同类型生境中的植被、地形、气候等生态因子联合或独立作用对昆虫群落的影响, 更有待进一步研究验证。
云南西双版纳自治州属于中国生物多样性保护的关键地区和热点地区, 也是我国面积最大、生态系统保存较完整的热带雨林地区, 孕育了丰富的物种多样性[6,29]。近年来, 西双版纳林地不断减少, 橡胶()和茶()的种植不断增加[30], 使该地区农业景观结构发生了变化, 并对生态系统造成了负面影响。
本研究在西双版纳3县市4种生境(保护区、次生林、人工林和农田), 调查分析不同类型生境中蝴蝶、蝽类和甲虫的多样性和群落结构的差异, 并分析植被类型、气候、地形等因子的耦合及独立作用对昆虫物种丰富度的影响, 探讨1)大尺度区域景观中3种类群昆虫群落如何响应生境特异性?不同类型生境对不同类群昆虫多样性的影响是否存在差异?2)不同类群昆虫在不同类型生境的群落结构是否存在差异?3)哪个生态因子对昆虫多样性影响起主要作用?本研究结果将有助于我们更加全面地了解特定生境对不同功能昆虫多样性的影响, 并为制定有针对性的保护措施和可持续景观规划提供数据支撑。
1 材料与方法
1.1 研究区概况
研究区位于云南西双版纳自治州(21°08′~ 22°33′N, 99°58′~101°34′E), 与缅甸和老挝接壤, 全区总土地面积19 120 km2, 下辖景洪市、勐海县、勐腊县3县市。西双版纳属北回归线以南热带湿润区[30]。全州各地年均温在20 ℃以上, 年均降雨量在1500~2000 mm。境内有多个国家级自然保护区和州级自然保护区。西双版纳国家级自然保护区由地区上互不相连的勐养、勐仑、勐腊、尚勇、曼搞5个子保护区组成, 总面积2474 km2; 纳板河流域国家级自然保护区分属景洪市和勐海县, 总面积266 km2; 布龙州级自然保护区属于勐海县, 面积354.85 km2; 易武州级自然保护区属于勐腊县, 面积333.70 km2。该地区热带经济作物主要有橡胶、茶叶、甘蔗(), 粮食经济作物以水稻()、旱稻、玉米()、大豆()为主, 森林植被以热带雨林植物、热带季风雨林和亚热带季风常绿阔叶林为主[31], 土地利用类型主要包括林地、橡胶林、茶园、水田、旱地等[32]。
1.2 样线设置及调查方法
于2019年8—9月采用样线调查法对西双版纳景洪、勐海、勐腊3县市的昆虫进行采样, 调查时, 采用地图规划与实地情况相结合的方法, 基于全面性、代表性、可达性原则布设调查样线, 在每个县市分别设置40条样线, 以10 km×10 km为一个网格, 网格内设置1~2条样线, 部分小于10 km (具体样线分布及设置情况见图1, 表1)。在调查区域内, 选择保护区、次生林、人工林和农田4种类型生境, 各类型生境具体情况如下: 1)保护区: 森林类型多样, 植物种类极为复杂, 主要为多年生高大乔木, 壳斗科(Fagaceae)、樟科(Lauraceae)等乔木为主, 受人为干扰较少。2)次生林: 原始林受到人为干扰(采伐、放牧、农业活动等)后形成次生林, 在西双版纳地区分布广泛, 常见优势树种有山黄麻()、白背桐()、中平树()等, 干扰较大的林缘多有入侵植物为主的灌草丛。3)人工林: 主要以橡胶、桉树()、杉木()、柚木()、西南桦()等为主, 林下主要是棕叶狗尾巴草()、飞机草()、紫茎泽兰()等草本植物。4)农田: 以农田、旱地为主, 种植水稻、旱稻、玉米、大豆等农作物, 田边主要是一些禾本科(Gramineae)和菊科(Compositae)植物为主的灌草丛。每天于昆虫活动频繁的9:00—18:00采集, 每条样线长1500 m, 宽约5 m, 进行匀速网扫采集, 整个调查, 每条样线网扫次数不少于500次, 时间为1 h。记录采集地点、样线编号、采集人、采集时间、生境类型、生境照片、GPS信息、路径信息以及昆虫生态照。将采集到的蝽和甲虫分别置于装有95%酒精离心管中保存, 采集的蝴蝶标本置于三角纸袋中, 带回实验室整理鉴定。根据形态分类方法, 利用《中国蝶类志》[33]《云南森林昆虫》[34]《中国蝽类昆虫鉴定手册》[35-36]《海南森林昆虫》[37]进行分类鉴定, 对不能鉴定到种的标本, 以形态种进行区分统计。
1.3 数据分析
本次共采集昆虫标本13 000余号, 其中蝴蝶、蝽、甲虫3个类群全部标本2588号, 蝴蝶标本采集于108条样线, 蝽类110条样线, 甲虫112条样线, 每个类群采集的样线数量在不同类型生境中的分布情况见表1。从这些数量和分布来看, 采集较为充分和全面。进行数据分析的是截止到2020年1月份的鉴定数据, 其中未鉴定到种的均以形态种区分。
1.3.1 不同类型生境昆虫物种数和特有物种数
利用Excel的数据透视表, 统计不同类型生境中不同类群昆虫的物种数、特有物种数及其多度, 并计算在不同类型生境中不同类群昆虫物种数、特有物种数及其多度所占的比例。
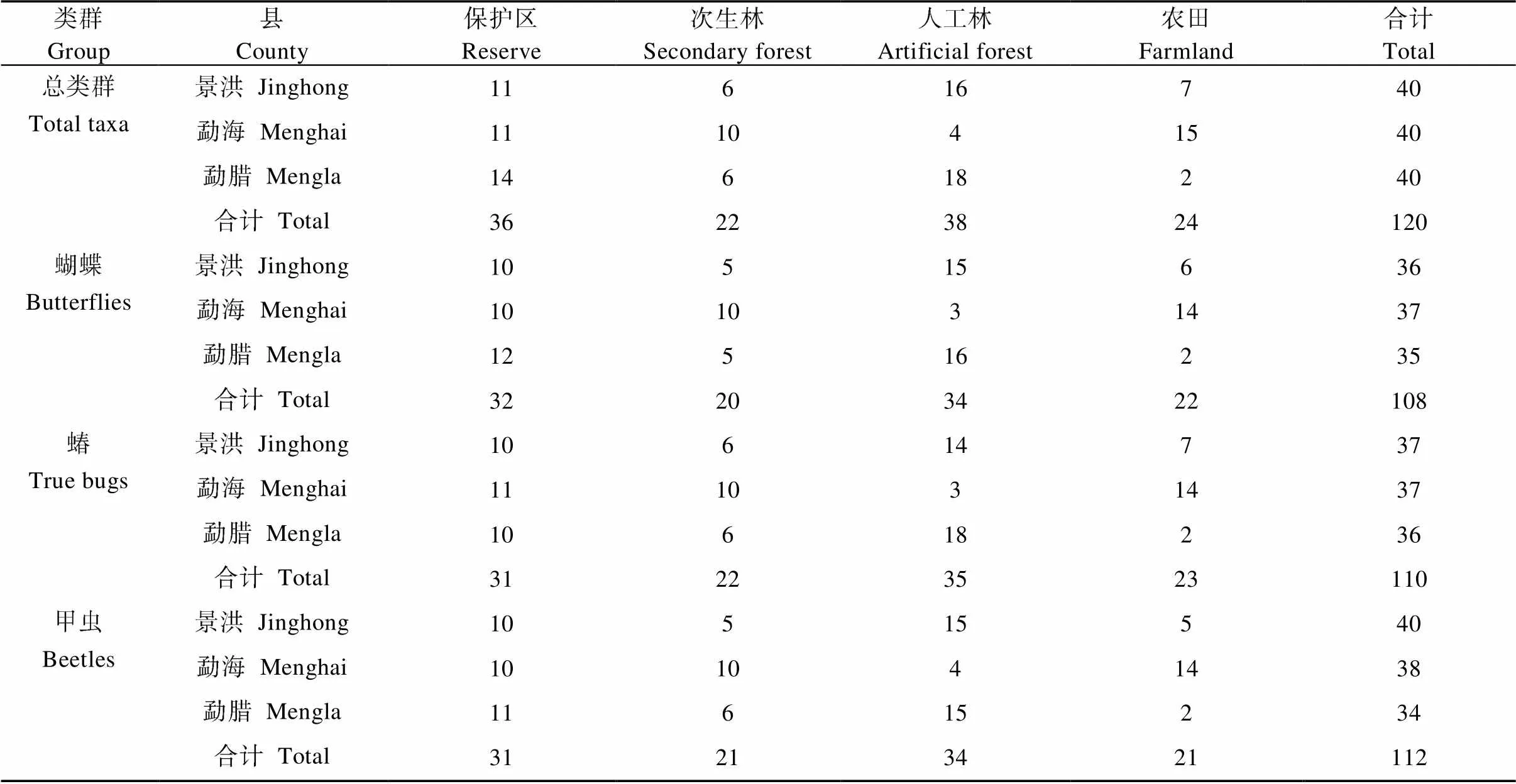
表1 西双版纳不同类型生境昆虫采集样线数量
1.3.2 不同类型生境昆虫群落的生境特异性分析
采用不同类群昆虫在不同类型生境的特异性指数, 比较不同类型生境之间对昆虫多样性影响的差异。计算昆虫群落的生境特异性指数, 计算公式如下[38]:
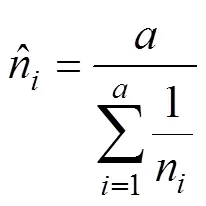
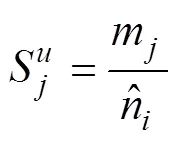
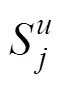
1.3.3 不同类型生境中昆虫群落结构分析
利用PRIMER7中CAP (canonical analysis of principle coordinates)主坐标规范分析对不同类型生境之间昆虫群落结构相似性分析(Clarke), 在分析前对数据进行平方根(square roots)转换处理, 并使用Permutation重复计算检验方法并获取值(999次重复)[39]。
1.3.4 不同类型生境中环境因子对昆虫物种丰富度的影响
利用ArcGIS获取调查样线的年均温、极端低温、海拔、植物均匀度指数、植物香农威纳指数、生境同质性指数, 并通过Minitab中的拟合Poisson模型对数据进行回归分析, 拟合物种丰富度与年均温、极端低温、海拔、植物均匀度指数、植物香农威纳指数、生境同质性指数、生境类型的相关性, 并得到值。
2 结果与分析
2.1 不同类型生境中昆虫物种数和特有物种数
本研究共采集3种类群昆虫个体2588头, 其中蝴蝶94种(744头), 蝽197种(1094头), 甲虫129种(750头)。不同类群的物种数和特有种数所占的比例均不同(表2), 不同类型生境中物种数和特有物种数都呈现出: 保护区>人工林>次生林>农田的趋势。在保护区生境中, 物种数和特有物种数均最多, 分别为235种和102种, 特有物种数占保护区总物种数的43.4%, 物种多度占保护区总物种多度的21.6%, 远大于其他3种类型生境中出现的特有物种数; 而在农田生境中出现的物种数和特有物种数均最少, 仅为143种和44种, 特有物种数占农田总物种数的30.8%, 物种多度占农田总物种多度的11.6%, 略高于人工林。
2.2 不同类型生境昆虫群落的生境特异性分析
合并3种类群昆虫比较其在不同类型生境中的差异发现, 3个类群昆虫群落的生境特异性指数差异不显著(3,57=2.054)(图2a), 但保护区与农田之间昆虫群落的生境特异性指数差异显著; 进一步比较不同类群昆虫在不同类型生境的差异发现, 甲虫的群落生境特异性指数在不同类型生境间差异显著(3,55=3.478)(图2d), 蝴蝶(3,38=1.504)(图2b)和蝽(3,53=1.153)(图2c)的群落生境特异性指数在不同类型生境间差异均不显著(图2)。

表2 不同生境类型蝴蝶、蝽、甲虫特有物种及物种数量及比例
不同类型生境对昆虫多样性的影响不同。保护区3种昆虫类群的群落生境特异性指数实测值高于期望值, 而其余生境类型中, 除了蝴蝶在人工林中的实测值高于期望值外, 其他昆虫群落生境特异性指数均低于实测值。将3种类群昆虫合并的昆虫群落生境特异性指数实测值从农田生境的10.8到保护区生境的20.1, 保护区最高, 农田最低; 而3个类群合并的昆虫群落生境特异性指数期望值波动较小, 说明保护区对昆虫多样性的保护具有积极作用。
R: 保护区; SF: 次生林; AF: 人工林; F: 农田。R: reserve; SF: secondary forest; AF: artificial forest; F: farmland.
2.3 不同类型生境中昆虫群落结构比较
从整体上看, 4种类型生境中昆虫群落结构差异不显著(ANOSIM Global=0.035), 其中保护区和农田的昆虫群落结构差异显著(ANOSIM Global=0.081)(表3, 图3a)。从不同类群来看, 4种类型生境中蝴蝶群落结构差异不显著(ANOSIM Global=0.052), 其中次生林和人工林的蝴蝶群落结构差异显著(ANOSIM Global=0.128)(表3, 图3b); 蝽类群落结构整体上差异不显著(ANOSIM Global=0.026,=0.145)(表3, 图3c), 其中保护区和次生林的蝽类群落结构差异显著(ANOSIM Global=0.091)。甲虫群落结构差异不显著(ANOSIM Global=−0.003)(表3, 图3d)。
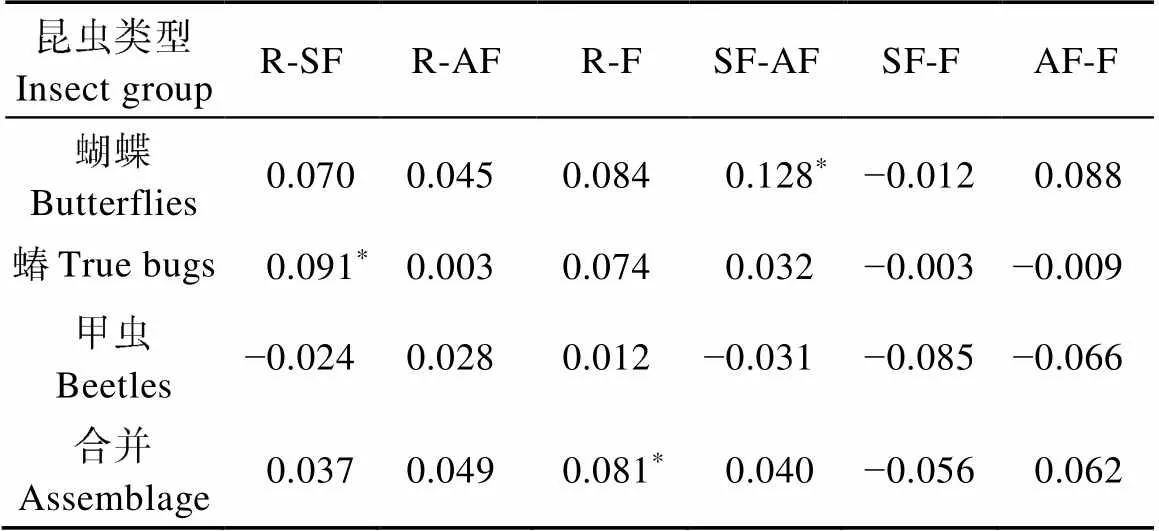
表3 不同类型生境间蝴蝶、蝽、甲虫组合和单一类群的群落结构差异显著性(ANOSIM Global R)
表中数据为不同类型生境间昆虫群落结构两两比较的统计值(ANOSIM Global R); R、SF、AF和F分别表示保护区、次生林、人工林和农田生境类型。*: 存在显著差异。Data in the table is the statistical value (ANOSIM Global R) of the comparison of insect community structure between two habitat types. R, SF, AF and F are habitat types of reserve, secondary forest, artificial forest and farmland, respectively. * means significant difference.
2.4 不同类型生境中生态因子对昆虫物种丰富度的影响
拟合Poisson模型分析结果显示, 生境类型对昆虫群落物种丰富度的影响差异极显著(< 0.01), 其他因子对昆虫物种丰富度的影响差异不显著(表4)。
3 讨论与结论
生境结构和环境的变化可能影响昆虫的基因流动和扩散[40], 对区域景观中昆虫多样性的研究, 更应该关注昆虫的物种丰富度、均匀度和多度对环境变化的响应[41-42]。一些研究表明, 生境类型的差异导致不同类群昆虫群落结构发生变化[43-44]。本研究发现超过30%的蝴蝶、蝽类和甲虫在不同类型生境中是生境特异性物种, 但不同生境存在一定差异, 对昆虫多样性具有不同的影响。本调查还发现, 不同类群昆虫物种数和特有物种数都呈现出保护区>人工林>次生林>农田的趋势, 昆虫群落的生境特异性指数和昆虫群落结构在保护区和农田之间均存在显著差异。这可能是在保护区的生境异质性较高, 物种多样性较高, 对昆虫群落的生境特异性贡献度较高; 而在农田植被单一, 昆虫群落的生境特异性贡献度较低。这表明不同类型生境对不同类群昆虫的特异性贡献具有一定差异, 这与Filgueiras等[18]研究不同类群昆虫多样性对生境破碎程度响应的结果类似。另外, 在西双版纳热带雨林中, 受人为干扰的生境与自然生境的昆虫群落结构明显不同, 这表明昆虫物种组成随着土地使用强度的增加, 昆虫物种组成变得更加简单。这一结果与Püttker等[45]和Liu等[46]的研究中, 发现生境丧失和改变增加了生境同质化对昆虫多样性影响的结果一致。另外, 不同类型生境景观结构的变化, 对不同类群昆虫的群落产生不同的影响, 特别是可获得的食物资源对昆虫群落结构的影响[41]。例如, 农田生境主要由农田和小路组成, 由于人为干扰频繁, 小路边缘只能提供少量的结构单一的非永久性的食物资源, 维持的生物多样性水平低[47], 而蝴蝶[27]和蝽类[48]需要拥有可持续性食物资源的生境。除保护区外, 次生林和人工林中不同类群昆虫群落的生境特异性指数均低于期望值, 而蝴蝶在人工林却高于期望值, 这一结果与Vu等[49-50]的研究结果类似, 这可能是在西双版纳地区, 人工林主要由橡胶林、茶树等构成, 林下有一些开花植物, 且灌木较少, 景观结构的变化和连通性提升, 有利于一些蝴蝶觅食和飞行[51]。

表4 不同类型生境中环境因子对昆虫物种丰富度的影响
目前的观点认为, 昆虫群落不仅容易受到小尺度上生态因子的影响, 在较大空间尺度上, 由于昆虫的扩散能力有限, 昆虫群落受到生态因子变化的影响更大[52]。虽然温度、地形等生态因子对生物多样性影响较大, 不同的地形和地貌都可能造成物种不同的分布格局[24], 但本研究发现, 以植被为主导的生境类型对昆虫的物种丰富度具有极显著的影响,但海拔等生态因子则对昆虫的物种丰富度没有显著影响, 这说明生境类型是影响昆虫群落的重要因素, 这一结果与Miao等[27]研究气候和生境变化对蝴蝶多样性影响的研究结果类似, 造成这种差异的原因可能是在较大空间尺度上, 一些昆虫对生态因子的变化并不敏感, 而更容易受到生境丧失和改变的影响[46]。
本研究揭示了不同类型生境对不同类群昆虫多样性的影响, 不同类群昆虫对生境丧失和生态因子变化做出响应的经验数据, 在区域景观中对昆虫多样性的保护具有重要意义。保护区可以为昆虫的生长和发育提供更加丰富的食物资源和庇护所[25], 在维持生物多样性和物种分布格局方面具有重要作用。但人工林和农田由于受到人为干扰严重, 环境质量遭到破坏, 植被丰富度和昆虫多样性下降严重, 建议保护区外应适当减少人为干扰, 这不仅可以使昆虫多样性得到保护, 在一定程度上也可以增加农作物的产量。本研究中数据分析只采用3种类群昆虫的鉴定数据, 物种完全鉴定后, 利用更多类群更能够从科级和属级水平上反映出不同类型生境间昆虫多样性和群落结构的差异。另外, 我们的调查只涉及到4种类型生境, 以后应该调查更多的生境, 比如: 溪流、草地等, 以进一步探索不同景观间昆虫生境特异性和群落结构的差异, 这将有助于我们更加全面地了解特定生境对区域景观中昆虫多样性的影响。
[1] HANSEN M C, POTAPOV P V, MOORE R, et al. High-resolution global maps of 21st-century forest cover change[J]. Science, 2013, 342(6160): 850–853
[2] BROSE U, HILLEBRAND H. Biodiversity and ecosystem functioning in dynamic landscapes[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2016, 371(1694): 20150267
[3] BUTCHART S H M, WALPOLE M, COLLEN B, et al. Global biodiversity: indicators of recent declines[J]. Science, 2010, 328(5982): 1164–1168
[4] RICKLEFS R E. A comprehensive framework for global patterns in biodiversity[J]. Ecology Letters, 2004, 7(1): 1–15
[5] JETZ W, THOMAS G H, JOY J B, et al. The global diversity of birds in space and time[J]. Nature, 2012, 491(7424): 444–448
[6] MYERS N, MITTERMEIER R A, MITTERMEIER C G, et al. Biodiversity hotspots for conservation priorities[J]. Nature, 2000, 403(6772): 853–858
[7] DAVID L ORME C, DAVIES R G, BURGESS M, et al. Global hotspots of species richness are not congruent with endemism or threat[J]. Nature, 2005, 436(7053): 1016–1019
[8] BENNETT A F, RADFORD J Q, HASLEM A. Properties of land mosaics: Implications for nature conservation in agricultural environments[J]. Biological Conservation, 2006, 133(2): 250–264
[9] MEEUS J H A. The transformation of agricultural landscapes in western Europe[J]. Science of the Total Environment, 1993, 129(1/2): 171–190
[10] STOATE C, BOATMAN N D, BORRALHO R J, et al. Ecological impacts of arable intensification in Europe[J]. Journal of Environmental Management, 2001, 63(4): 337–365
[11] BASSET Y, CIZEK L, CUENOUD P, et al. Arthropod diversity in a tropical forest[J]. Science, 2012, 338(6113): 1481–1484
[12] MISOF B, LIU S L, MEUSEMANN K, et al. Phylogenomics resolves the timing and pattern of insect evolution[J]. Science, 2014, 346(6210): 763–767
[13] NEMÉSIO A, SILVA D P, NABOUT J C, et al. Effects of climate change and habitat loss on a forest-dependent bee species in a tropical fragmented landscape[J]. Insect Conservation and Diversity, 2016, 9(2): 149–160
[14] WINDER L, ALEXANDER C J, HOLLAND J M, et al. Predatory activity and spatial pattern: the response of generalist carabids to their aphid prey[J]. Journal of Animal Ecology, 2005, 74(3): 443–454
[15] 洪雪萌, 戈昕宇, 李俊兰. 赛罕乌拉自然保护区蝶类多样性及其影响因素[J]. 生物多样性, 2018, 26(6): 590–600 HONG X M, GE X Y, LI J L. Butterfly diversity and its influencing factors in Saihanwula Nature Reserve[J]. Biodiversity Science, 2018, 26(6): 590–600
[16] 李巧, 陈又清, 陈彦林. 紫胶林-农田复合生态系统蝽类昆虫群落多样性[J]. 云南大学学报: 自然科学版, 2009, 31(2): 208–216 LI Q, CHEN Y Q, CHEN Y L. Diversity of heteropteran communities in lac plantation-farmland ecosystem[J]. Journal of Yunnan University: Natural Sciences Edition, 2009, 31(2): 208–216
[17] HIRAO T, MURAKAMI M, KUBOTA Y. Species abundance distributions of moth and beetle assemblages in a cool-temperate deciduous forest[J]. Insect Conservation and Diversity, 2013, 6(4): 494–501
[18] FILGUEIRAS B K C, MELO D H A, ANDERSEN A N, et al. Cross-taxon congruence in insect responses to fragmentation of Brazilian Atlantic forest[J]. Ecological Indicators, 2019, 98: 523–530
[19] 朱纪元, 李景科, 程赛赛, 等. 小兴安岭阔叶红松林局地尺度地表鞘翅目成虫群落结构[J]. 东北林业大学学报, 2016, 44(12): 57–63 ZHU J Y, LI J K, CHENG S S, et al. Community structure of adult Coleoptera on local scale in A mixed broadleaved Korean pine forest in the Xiaoxing’an mountains[J]. Journal of Northeast Forestry University, 2016, 44(12): 57–63
[20] FAHRIG L, BAUDRY J, BROTONS L, et al. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes[J]. Ecology Letters, 2011, 14(2): 101–112
[21] 陈又清, 李巧, 王莹, 等. 普洱市亚热带季风常绿阔叶林区蝽类昆虫多样性[J]. 西北林学院学报, 2010, 25(3): 137–142 CHEN Y Q, LI Q, WANG Y, et al. Diversity of true bugs in subtropical monsoon evergreen broadleaved forest in Pu’er City, Yunnan[J]. Journal of Northwest Forestry University, 2010, 25(3): 137–142
[22] GESSÉ F, MONLEÓN-GETINO T, GOULA M. Biodiversity analysis of true bug assemblages (Hemiptera, Heteroptera) in four habitats in the garraf natural park (Barcelona, Spain)[J]. Journal of Insect Science, 2014, 14(1): 283
[23] 戈昕宇. 赛罕乌拉自然保护区蝽类昆虫群落多样性研究[D]. 呼和浩特: 内蒙古大学, 2019 GE X Y. A study on the species diversity of true bugs community in Saihanwula Nature Reserve[D]. Hohhot: Inner Mongolia University, 2019
[24] BIRKHOFER K, GOSSNER M M, DIEKÖTTER T, et al. Land-use type and intensity differentially filter traits in above- and below-ground arthropod communities[J]. The Journal of Animal Ecology, 2017, 86(3): 511–520
[25] SWART R C, SAMWAYS M J, ROETS F. Latitude, paleo-history and forest size matter for Afromontane canopy beetle diversity in a world context[J]. Biodiversity and Conservation, 2021, 30(3): 659–672
[26] 郝淑莲, 薛琪琪, 冯丹丹, 等. 山西南部山地蝴蝶多样性与生态位差异比较研究[J]. 生态与农村环境学报, 2019, 35(10): 1314–1321 HAO S L, XUE Q Q, FENG D D, et al. Comparative study on butterfly diversity and niche difference in mountainous region of southern Shanxi Province[J]. Journal of Ecology and Rural Environment, 2019, 35(10): 1314–1321
[27] MIAO B G, PENG Y Q, YANG D R, et al. Climate and land-use interactively shape butterfly diversity in tropical rainforest and savanna ecosystems of southwestern China[J]. Insect Science, 2020. DOI: 10.1111/1744–7917.12824
[28] ANTÃO L H, MCGILL B, MAGURRAN A E, et al. Β-diversity scaling patterns are consistent across metrics and taxa[J]. Ecography, 2019, 42(5): 1012–1023
[29] 朱华, 王洪, 李保贵, 等. 西双版纳森林植被研究[J]. 植物科学学报, 2015, 33(5): 641–726 ZHU H, WANG H, LI B G, et al. Studies on the forest vegetation of Xishuangbanna[J]. Plant Science Journal, 2015, 33(5): 641–726
[30] 刘晓娜, 封志明, 姜鲁光, 等. 西双版纳土地利用/土地覆被变化时空格局分析[J]. 资源科学, 2014, 36(2): 233–244 LIU X N, FENG Z M, JIANG L G, et al. Spatial-temporal pattern analysis of land use and land cover change in Xishuangbanna[J]. Resources Science, 2014, 36(2): 233–244
[31] 杜加强, 舒俭民, 王跃辉, 等. 青藏高原MODIS NDVI与GIMMS NDVI的对比[J]. 应用生态学报, 2014, 25(2): 533–544 DU J Q, SHU J M, WANG Y H, et al. Comparison of GIMMS and MODIS normalized vegetation index composite data for Qinghai-Tibet Plateau[J]. Chinese Journal of Applied Ecology, 2014, 25(2): 533–544
[32] 刘晓娜, 封志明, 姜鲁光, 等. 西双版纳橡胶林地的遥感识别与数字制图[J]. 资源科学, 2012, 34(9): 1769–1780 LIU X N, FENG Z M, JIANG L G, et al. Rubber plantations in Xishuangbanna: Remote sensing identification and digital mapping[J]. Resources Science, 2012, 34(9): 1769–1780
[33] 周尧. 中国蝶类志修订本上下册[M]. 郑州: 河南科学技术出版社, 2000 ZHOU Y. Monograph of Chinese Butterflies (Revised Edition) (Two volumes)[M]. Zhengzhou: Henan Scientific and Technological Publishing House, 2000
[34] 云南省林业厅, 中国科学院动物研究所. 云南森林昆虫[M]. 昆明: 云南科技出版社, 1987 Forest Department of Yunnan Province; Institute of Zoology, Chinese Academy of Sciences. Forest Insects of Yunnan[M]. Kunming: Yunnan Science and Technology Press, 1987
[35] 萧采瑜. 中国蝽类昆虫鉴定手册(半翅目异翅亚目)第一册[M]. 北京: 科学出版社, 1977 XIAO C Y. Identification Handbook of Chinese True Bugs (Volume Ⅰ) [M]. Beijing: Science Press, 1997
[36] 萧采瑜. 中国蝽类昆虫鉴定手册(半翅目异翅亚目)第二册[M]. 北京: 科学出版社, 1981 XIAO C Y. Identification Handbook of Chinese True Bugs (Volume Ⅱ) [M]. Beijing: Science Press, 1981
[37] 黄复生. 海南森林昆虫[M]. 北京: 科学出版社, 2002 HUANG F S. Forest Insects of Hainan[M]. Beijing: Science Press, 2002
[38] HALVORSEN R, EDVARDSEN A. The concept of habitat specificity revisited[J]. Landscape Ecology, 2009, 24(7): 851–861
[39] ANDERSON M J, WILLIS T J. Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology[J]. Ecology, 2003, 84(2): 511–525
[40] BALAKRISHNAN S, SRINIVASAN M, MOHANRAJ J. Diversity of some insect fauna in different coastal habitats of Tamil Nadu, southeast Coast of India[J]. Journal of Asia-Pacific Biodiversity, 2014, 7(4): 408–414
[41] MAJEED W, RANA N, QAMAR S U R, et al. Diversity of foliage insects around different canal territories: a case study of Dingroo and Kamal Pur canal, Faisalabad, Pakistan[J]. GSC Biological and Pharmaceutical Sciences, 2019, 6(1): 7–15
[42] RAMZAN U, MAJEED W, RANA N, et al. Occurrence of different insect species with emphasis on their abundance and diversity in different habitats of Faisalabad, Pakistan[J]. International Journal of Tropical Insect Science, 2020. DOI: 10.1007/s42690-020-00314-5.
[43] BARTON P S, CUNNINGHAM S A, MANNING A D, et al. The spatial scaling of beta diversity[J]. Global Ecology and Biogeography, 2013, 22(6): 639–647
[44] DRISCOLL D A, BANKS S C, BARTON P S, et al. Conceptual domain of the matrix in fragmented landscapes[J]. Trends in Ecology & Evolution, 2013, 28(10): 605–613
[45] PÜTTKER T, DE ARRUDA BUENO A, PRADO P I, et al. Ecological filtering or random extinction? Beta-diversity patterns and the importance of niche-based and neutral processes following habitat loss[J]. Oikos, 2015, 124(2): 206–215
[46] LIU C, GUÉNARD B, BLANCHARD B, et al. Reorganization of taxonomic, functional, and phylogenetic ant biodiversity after conversion to rubber plantation[J]. Ecological Monographs, 2016, 86(2): 215–227
[47] REBERG-HORTON S C, MUELLER J P, MELLAGE S J, et al. Influence of field margin type on weed species richness and abundance in conventional crop fields[J]. Renewable Agriculture and Food Systems, 2011, 26(2): 127–136
[48] KŐRÖSI Á, BATÁRY P, OROSZ A, et al. Effects of grazing, vegetation structure and landscape complexity on grassland leafhoppers (Hemiptera: Auchenorrhyncha) and true bugs (Hemiptera: Heteroptera) in Hungary[J]. Insect Conservation and Diversity, 2012, 5(1): 57–66
[49] VU L V. Diversity and similarity of butterfly communities in five different habitat types at Tam Dao National Park, Vietnam[J]. Journal of Zoology, 2009, 277(1): 15–22
[50] VU L V, QUANG VU C. Diversity pattern of butterfly communities (Lepidoptera, Papilionoidae) in different habitat types in a tropical rain forest of southern Vietnam[J]. ISRN Zoology, 2011, 2011: 1–8
[51] ÖCKINGER E, VAN DYCK H. Landscape structure shapes habitat finding ability in a butterfly[J]. PLoS One, 2012, 7(8): e41517 DOI:10.1371/journal.pone.0041517
[52] HENDRICKX F, MAELFAIT J P, DESENDER K, et al. Pervasive effects of dispersal limitation on within- and among-community species richness in agricultural landscapes[J]. Global Ecology and Biogeography, 2009, 18(5): 607–616
Habitat-specific influences on insect diversity in regional landscapes: A case study of Xishuangbanna*
ZHANG Xiang1, LU Zhixing1, WANG Qing1, GAO Shutong1, TANG Chunying2, LI Qiao2, CHEN Youqing1**
(1. Research Institute of Resources Insect, Chines Academy of Forestry, Kunming 650224, China; 2. Southwest Forestry University, Kunming 650224, China)
Habitat-specific plays an important role in the maintenance of biodiversity. This study aimed to understand the influence of habitat type on the diversity of different insect groups and the differences in community structure across the regional landscape. This study also explored the distribution patterns and maintenance mechanisms of insect diversity on a large scale. From August to September 2019, the sample line survey method was used in the Xishuangbanna region to estimate the number of species and endemic species of butterflies, true bugs, and beetles in four different habitat types (reserve, secondary forest, artificial forest, and farmland) and to analyze the habitat specificity index of the insect communities, the community structure similarity, and the relationships between species richness and the ecological factors. A total of 2588 insects were collected, including 94 butterfly species (744 heads), 197 bug species (1094 heads), and 129 beetle species (750 heads). The number of insect species and endemic species in the reserve was higher than those in the other habitats, and the observed habitat specificity index of the insect community was higher than the expected value in the reserve. The observed habitat specificity index values for the other habitat types were lower than the expected values. There was no significant difference in the habitat specificity index of the insect communities in different habitat types (3,57=2.054), but there was a significant difference in the habitat specificity index of beetles (3,55=3.478). The habitat specificity index of butterflies (3,38=1.504) and true bugs (3,53=1.153) did not differ. Analysis of the community structure similarity showed that the insect community structure of the three groups significantly differed between the reserve and farmland. There was a significant difference in the community structure of butterflies between secondary forests and artificial forests (=0.037), and there was a significant difference in the community structure of true bugs between the reserve and secondary forests (=0.029). There was no difference in the insect community structure of beetles (=0.507), and only habitat type had a significant effect on the species richness of the insect community. The other ecological factors did not have a significant effect. In large-scale regional landscapes, the reserve had the greatest impact on diversity, and the habitat types were closely related to the species richness of the insect communities. Well-protected natural forests are important for maintaining the distribution and diversity of insect communities in regional landscapes.
Regional landscape; Insect community habitat-specific; Insect diversity; Community structure; Xishuangbanna
10.13930/j.cnki.cjea.200684
张翔, 卢志兴, 王庆, 高舒桐, 唐春英, 李巧, 陈又清. 区域景观中生境特异性对昆虫多样性的影响——以西双版纳为例[J]. 中国生态农业学报(中英文), 2021, 29(5): 771-780
ZHANG X, LU Z X, WANG Q, GAO S T, TANG C Y, LI Q, CHEN Y Q. Habitat-specific influences on insect diversity in regional landscapes: A case study of Xishuangbanna[J]. Chinese Journal of Eco-Agriculture, 2021, 29(5): 771-780
Q968.1

* 生态环境部生物多样性调查、观测和评估项目(2019—2023年)资助
陈又清, 主要从事昆虫生态学研究。E-mail: cyqcaf@126.com
张翔, 研究方向为昆虫生态学。E-mail: m18213456905@163.com
2020-08-20
2021-01-27
* This study was supported by the Biodiversity Survey, Observation and Assessment Program of the Ministry of Ecology and Environment, China (2019−2023).
, E-mail: cyqcaf@126.com
Aug. 20, 2020;
Jan. 27, 2021
