稻蛙共作对水稻-紫云英轮作系统氨挥发的影响*
2021-05-07陈慧妍沙之敏吴富钧方凯凯徐春花杨晓磊朱元宏曹林奎
陈慧妍, 沙之敏, 吴富钧,2, 方凯凯, 徐春花, 杨晓磊, 朱元宏, 曹林奎**
稻蛙共作对水稻-紫云英轮作系统氨挥发的影响*
陈慧妍1, 沙之敏1, 吴富钧1,2, 方凯凯1, 徐春花3, 杨晓磊3, 朱元宏4, 曹林奎1**
(1. 上海交通大学农业与生物学院 上海 200240; 2. 福建农林大学农学院 福州 350002; 3. 上海市农业技术推广服务中心 上海 201103; 4. 上海青浦现代农业园区发展有限公司 上海 201717)
氨挥发是稻田氮素损失的主要途径之一, 探究稻田生态种养模式对稻田土壤氨挥发产生的影响, 可为该模式的生态环境效益评价提供理论依据。为评估稻蛙共作模式对水稻-紫云英轮作系统氨挥发的影响, 通过开展田间小区试验, 采用密闭式间歇抽气法采集氨气, 对水稻-紫云英轮作系统的土壤氨挥发及其影响因素进行研究。试验共设置3个处理: 空白对照(CK, 不施肥, 不放蛙)、常规水稻种植模式(CR, 施化肥, 不放蛙)、稻蛙共作模式(RF, 施化肥, 放蛙)。结果表明: 稻蛙共作模式水稻季氨挥发累积量为47.02 kg∙hm−2, 占当季施氮量12.9%; 其后茬紫云英季的氨挥发累积量为16.27 kg∙hm−2; 全年轮作系统的氨挥发累积量为63.29 kg∙hm−2,较常规水稻种植模式的氨挥发累积量降低15.3%。稻蛙共作模式全年水稻-紫云英轮作系统的氨挥发累积量占施氮量的比例为17.4%, 显著低于常规水稻种植模式所占比例(20.5%)。水稻田面水的铵态氮浓度是影响水稻季氨挥发的主要因素, 水稻田面水pH、水温、气温、风速等因素的影响次之, 随温度上升, 水稻田面水铵态氮浓度对氨挥发速率的影响逐渐增大。放蛙对水稻产量、水稻产量构成因素、氮肥利用效率及后茬作物紫云英产量的影响不显著。综上所述, 稻蛙共作模式在水稻-紫云英轮作系统中具备一定的氨减排潜力, 但该模式对稻田氨挥发影响的长期效应及其影响机理仍需进一步研究。
水稻-紫云英轮作; 稻蛙共作; 氨挥发; 产量
氨(NH3)是大气中一种主要的碱性痕量气体, 也是一种主要的大气污染物, 极易与空气中的酸性气体反应生成PM2.5的前体物质影响空气质量, 同时容易引起土壤酸化、水体富营养化等多种环境问题[1-2]。据报道, 我国每年的氨排放量约为14 Tg[3]。农业是主要的氨排放来源, 占全球人为源氨排放量的90%, 其中由于农田施肥导致的氨排放占农业源氨排放量的40%[4]。与小麦()田等旱地相比, 水稻()田的高温和淹水环境更有利于氨的挥发[5-6]。He等[7]研究表明稻田中施用的氮肥约有13.2%~47.0%通过氨挥发损失, 说明氨挥发是稻田氮素损失的主要途径之一。影响氨挥发的因素有很多, 包括氮肥管理技术(如肥料类型、肥料用量和施肥方式)[8-9]、土壤理化性质(如土壤pH和土壤类型)[10]、气象因素(如温度、降雨量和风速)[11]、作物生育期等, 其中水稻田面水铵态氮浓度和pH是稻田氨挥发的直接影响因素。
稻蛙共作模式是一种有利于实现农业可持续发展的生态农业模式。不少研究表明, 在稻田中引入蛙可以有效减少稻飞虱等水稻害虫的种群数量, 并有利于改善土壤的养分状况, 在一定程度上提高土壤微生物生物量和土壤脱氢酶、过氧化氢酶、酸性磷酸酶等多种土壤酶活性, 从而促进水稻生长, 提高水稻结实率与籽粒产量[12-18]。此外, 稻蛙共作模式还具备降低温室气体CH4排放的潜力[19]。研究表明多种稻田生态种养模式具备影响稻田生态系统中养分利用和氮素循环的潜力。Gao等[20]和Xu等[21]研究表明, 稻鸭共作模式可以减少稻田氮素的渗漏损失, 并提高水稻的产量, 但其排泄向稻田中添加了氮和有机碳底物, 促进了硝化和反硝化过程, 增加了N2O的排放。Frei等[22]研究表明, 稻鱼共作模式中鱼会消耗水稻田面水中的溶解氧, 降低水稻田面水pH, 减少N2O的排放。然而, Li等[23]研究表明, 稻鸭共作模式和稻鱼共作模式在放鸭和放鱼后均能降低稻田氨挥发。类似地, 在稻蟹共作模式中, 蟹的投入提高了表层土壤酶活性, 蟹投入后能少量降低稻田氨挥发并提高水稻的氮素积累量[24-25]。稻蛙共作模式在我国南方地区有较为广泛的应用, 与其他生态种养模式不同的是, 蛙是稻田中原生的两栖动物, 在稻田中引入蛙是一种更加生态和自然的水稻种植模式, 但关于该模式的氨挥发影响还鲜见报道。本文通过田间小区试验, 比较稻蛙共作模式与常规水稻种植模式的稻田及全年水稻-紫云英()轮作系统的土壤氨挥发损失, 并比较氨挥发各影响因素的变化, 为评估稻蛙共作模式对生态环境的影响提供理论依据。
1 材料与方法
1.1 试验地点
试验于2019年在上海市青浦现代农业园区(121.03°E, 30.97°N)进行。该地区属于亚热带季风气候, 年平均气温15.5 ℃, 年平均降水量1056 mm, 年平均日照时数1960.70 h, 无霜期约247 d。2019年水稻生长期的日均气温、降雨量和风速如图1所示。本试验区的土壤类型为湖沼相沉积物起源的青紫泥水稻土, 试验前土壤基础理化性质为全氮1.70 g∙kg−1, 全磷0.95 g∙kg−1, 速效钾0.11 g∙kg−1, 有机质29.60 g∙kg−1, pH为7.02。
1.2 试验材料
供试水稻品种为‘青角307’, 供试蛙种为虎纹蛙(), 肥料为高塔尿基复合肥(28% N, 6% P2O5, 6% K2O)、尿素(46% N)、过磷酸钙(12% P2O5)和氯化钾(60% K2O)。
1.3 试验设计及田间管理
本试验设置3种处理: 空白对照(CK), 水稻-紫云英轮作, 水稻季不施肥, 不放蛙; 常规水稻种植模式(CR), 水稻-紫云英轮作, 水稻季施化肥, 不放蛙; 稻蛙共作模式(RF), 稻蛙共作-紫云英轮作, 水稻季施化肥, 放蛙。每个处理设3个重复, 共9个小区, 随机区组排列, 每个小区56 m2(7 m×8 m)。
每个小区有独立的进水口和排水口, 小区之间田埂用防渗膜包裹防止小区间的测渗, 稻蛙共作小区在田埂使用围蛙网防止蛙逃逸, 并在围蛙网田埂内堆砌部分小土堆供蛙栖息, 整个试验田块上方围防鸟网避免蛙被天敌捕食。在稻季, CR与RF处理采用完全相同的施肥方案, 共分3次施入等量等比例化肥, N、P2O5、K2O总投入量分别为300.00 kg∙hm−2、112.54 kg∙hm−2和90.04 kg∙hm−2,其中磷肥和钾肥在基肥时一次性施入, 氮肥分别在6月19日施基肥, 7月6日第1次追肥, 7月25日第2次追肥, 具体氮肥施用量见表1。6月23日进行人工插秧, 密度为16 cm株距×16 cm行距, 之后进行人工拔草, 不使用化学农药。7月23日, 按7500只∙hm−2蛙密度向稻蛙共作小区投放体重为20 g左右的虎纹蛙幼蛙。有研究[26]表明在自然状态下田间昆虫量可基本满足每公顷900~9000只投蛙量的捕食需求, 故本研究在放蛙后5 d内每天傍晚在田埂边上投放少量蛙饵助其适应田间环境, 随后蛙自主捕食田间昆虫, 不另外喂食饲料。通过观察蛙在田间的存活情况, 及时补充适量蛙苗, 以保证蛙在田间持续的活动影响。11月5日, 水稻人工收割, 投放的虎纹蛙不回收。在紫云英季, 3种处理的田间管理均相同, 10月18日以60 kg∙hm−2的播种量撒播紫云英种子, 不施肥, 水稻收获后在小区内开十字沟疏水, 次年4月23日对紫云英翻耕还田, 水稻季投放的虎纹蛙不回收。

表1 水稻-紫云英轮作系统不同处理水稻季氮肥纯氮施用量
1.4 样品采集与测定方法
1.4.1 NH3的采样与测定
水稻季每次施肥后连续10 d采样, 此后1周采样1次; 紫云英季每2~4周采样1次。每次采样时间为7:00—9:00和15:00—17:00。NH3的采样使用改进的密闭室间歇抽气法[27]。采样装置由真空泵、流量计、洗气瓶、抽气室和换气杆等组成, 其中抽气室为直径20 cm、高30 cm、底部开放和顶部有两个通气孔的圆柱形有机玻璃罩。通过真空泵抽气经洗气瓶内的吸收液(100 mL 0.0005 mol∙L−1H2SO4)吸收氨气, 抽气速率为15~20 L∙min−1。采样时, 将抽气室置于每个小区正中央区域, 在抽气结束后, 将抽气室从田面移开, 以保证蛙能在采样点处进行活动, 消除抽气室内外环境的影响。采回的吸收液使用靛酚蓝分光光度法测定所吸收氨的含量。
1.4.2 其他样品的采样与测定
水稻季试验期间, 每次施肥后连续10 d上午8:00采集水稻田面水, 于冰箱保存至实验室分析。水稻收获时, 采用样方法收割水稻, 风干籽粒后称重测定水稻产量。紫云英翻耕前, 采用样方法收割紫云英地上部分, 风干后称重测定紫云英产量。

1.5 数据分析
对不同处理的差异显著性进行单因素方差分析,采用LSD法进行多重比较(<0.05), 采用逐步回归分析氨挥发对各影响因素的响应, 各相关性采用回归分析法。采用SPSS 25软件进行数据统计和分析, 用Origin 9.1绘图。
2 结果与分析
2.1 水稻-紫云英轮作系统的氨挥发量
在水稻季, 各处理的氨挥发累积量为37.08~55.72 kg∙hm−2, 其中CR和RF水稻季氨挥发累积量分别占当季施氮量的15.3%和12.9%, RF比CR的水稻季氨挥发累积量降低15.6%, 但无显著差异。在紫云英季, 各处理的氨挥发累积量为16.27~19.20 kg∙hm−2, 均无显著差异。全年水稻-紫云英轮作系统的氨挥发累积量为56.27~74.76 kg∙hm−2, RF较CR的全年轮作系统氨挥发累积量降低15.3%, RF和CR全年轮作系统氨挥发累积量分别占施氮量的17.4%和20.5%, RF显著低于CR(表2)。与CK相比, CR施肥显著增加了稻田氨挥发, 但RF全年氨挥发累积量与CK无显著差异。这说明放蛙能在一定程度上减少氨挥发, 尽管CR与RF的氨挥发累积量无显著差异。
在水稻季, CR和RF在3次施肥后的氨挥发速率迅速升高, 在施肥后1~3 d内达到峰值, CR和RF最高氨挥发速率峰值均在穗肥期间, 分别为3.97 kg∙hm−2∙d−1和3.10 kg∙hm−2∙d−1,随后逐步下降, 除基肥外其余两次追肥7 d后氨挥发速率降至与CK一致(图2)。CR和RF的氨挥发损失率分别为5.1%和2.7%, RF的氨挥发损失率略低于CR, 但无显著差异。在紫云英季氨挥发速率稳定在0.30 kg∙hm−2∙d−1以内, 各处理动态变化基本一致。

表2 水稻-紫云英轮作系统不同种植模式氨挥发累积量
各处理的含义见表1。不同小写字母表示处理间在<0.05水平差异显著。The meaning of each treatment is shown in the table 1. Different lowercase letters indicate significant differences among treatments at<0.05 level.
2.2 水稻田面水pH、和浓度变化
各处理的含义见表1。The meaning of each treatment is shown in the table 1.
各处理的含义见表1。The meaning of each treatment is shown in the table 1.
各处理的水稻田面水pH变化范围为6.97~8.27, CK、CR、RF水稻田面水的平均pH分别为7.43、7.37和7.41。在基肥和第1次追肥期间, 各处理水稻田面水pH的变化趋势较为一致; 而在第2次追肥期间不施肥处理CK的水稻田面水pH略高于施肥处理CR和RF(图3c)。在放蛙后, 稻蛙共作模式RF的水稻田面水pH略高于常规水稻种植模式CR, 但差异不显著。
**表示在<0.01水平显著相关。** denotes significant correlation at<0.01 level.
为进一步分析氨挥发速率与水稻田面水pH的关系, 使用Growth模型分别拟合了水稻基肥期、第1次追肥期和第2次追肥期施肥后连续10 d的氨挥发速率与水稻田面水pH的关系(图5)。3个时期的模型均通过显著性检验。在3个时期中, 第1次追肥期的模型拟合优度最高, 该时期的模型说明当田面水pH分别在7.00~7.50、7.50~8.00和8.00~8.50范围内时, pH每增加0.5个单位, 氨挥发速率分别增加0.216 kg∙hm−2∙d−1、0.467 kg∙hm−2·d−1和1.012 kg∙hm−2∙d−1(图5b)。说明该阶段随着水稻田面水pH的逐渐增大, 稻田氨挥发速率也逐渐增大且增大的速率越来越高。
2.3 氨挥发及其影响因素的关系

表3 稻田氨挥发速率与各影响因素的逐步回归分析
2.4 不同种植模式的水稻和紫云英产量
常规水稻种植模式CR与稻蛙共作模式RF的水稻产量分别为8.33 t∙hm−2和8.32 t∙hm−2, 水稻氮素利用率分别为41.0%和35.3%。分别计算3种模式水稻单位产量的氨挥发累积排放量, RF的平均单位产量氨挥发累积排放量较CR降低16.2%, 但无显著差异,这与各模式稻季氨挥发累积量的结果一致。3种模式的紫云英鲜重产量为38.53~44.23 t∙hm−2, 风干后测得紫云英生物量为5.48~6.57 t∙hm−2, 均无显著差异。各处理水稻的收获指数为37.1%~41.9%, 分析不同模式水稻产量构成因素及植株性状, 施肥处理CR和RF较CK显著增加了单位面积有效穗数和株高, 而CR与RF间各产量构成因素无显著差异(表4)。
3 讨论
3.1 水稻-紫云英轮作系统的氨挥发量
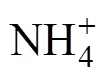
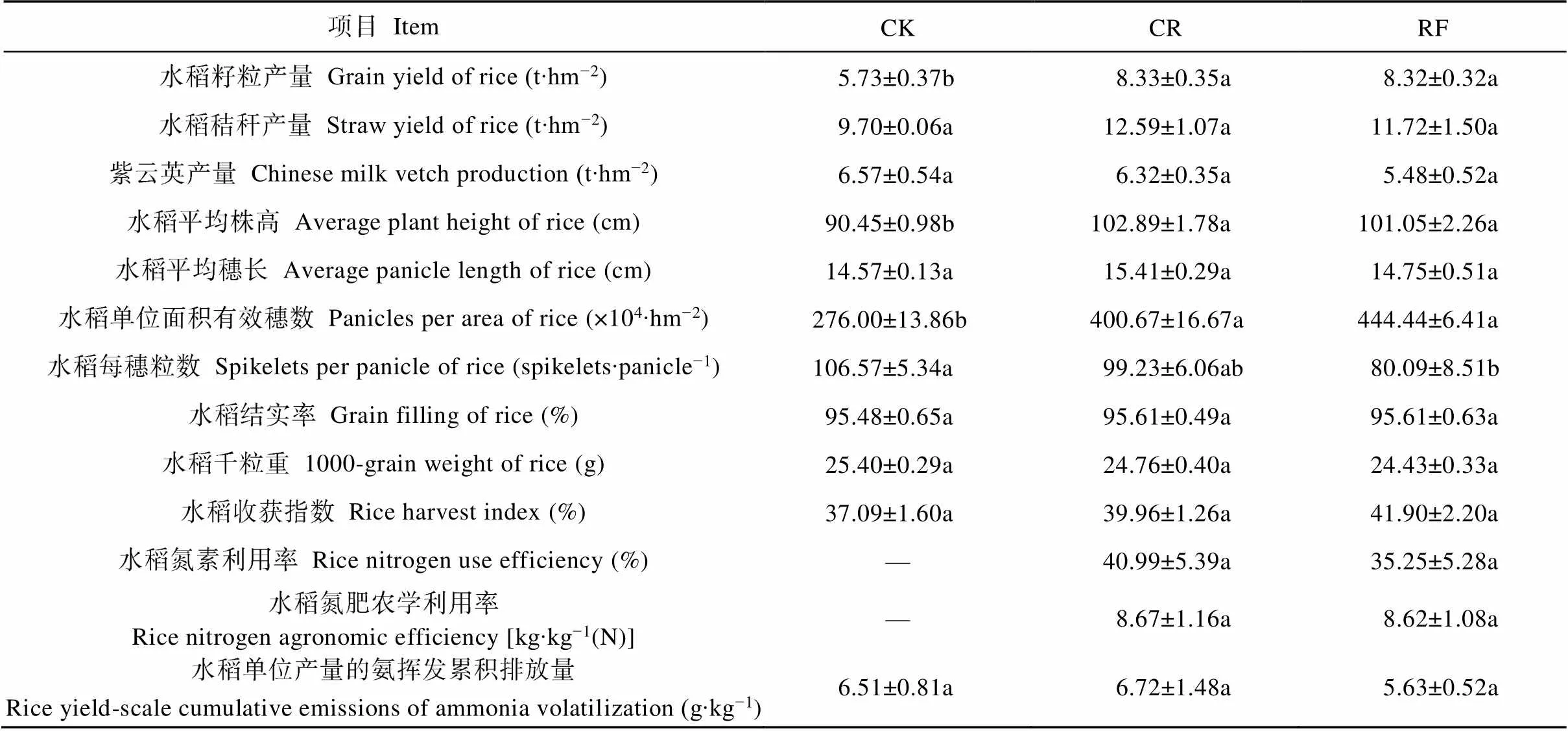
表4 不同种植模式水稻-紫云英轮作系统产量和氮肥利用效率
各处理的含义见表1。同行不同小写字母表示处理间差异显著。The meaning of each treatment is shown in the table 1. Different lowercase letters in the same line mean significant differences at<0.05 level.
3.2 水稻-紫云英轮作系统氨挥发的影响因素
本研究中, 稻蛙共作模式的氨挥发累积量与常规水稻种植模式相比偏低, 其水稻氮素利用率与常规水稻种植模式相比也偏低, 尽管该偏低的结果无显著差异。研究学者通常认为较高氮素利用率的水稻更加具备减少稻田氮素损失的潜力, 但近年来Chen等[38]研究表明稻田氨挥发量与水稻氮素利用率两者间并无直接相关关系, 稻田氨挥发量越低, 并不意味着水稻氮素利用率越高, 稻田生态系统中的氮素可能通过其他途径损失。陈贵等[39]研究表明, 水稻在抽穗期之前的吸氮量超过全生育期吸氮量的80.0%, 推测较低氮素利用率的水稻可能在前期吸氮能力更强, 施肥引起的氨挥发较低, 但在后期通过渗漏、硝化和反硝化等途径引起的氮素损失量更大。有研究表明, 绿色稻蛙共作模式与有机稻蛙共作模式的渗漏水总氮浓度分别比常规水稻种植模式降低8.8%和18.0%, 但在相同施肥条件下稻蛙共作对稻田氮素渗漏损失的影响尚不清楚, 有待进一步研究[40]。
尽管本研究稻田中蛙的投入直接降低稻田氨挥发的效果有限, 但稻蛙共作模式下生产的稻米品质得到改善, 市场价格和市场需求可观, 在提高生态效益的同时也提高了经济效益[41]。Lia等[42]通过录像观察发现稻田中褐飞虱的主要捕食者是蛙类, 蛙在稻田中起到最直接的作用是捕食水稻害虫, 既加速了养分在稻田生态系统中的循环, 又减少了化学农药的使用, 提高了水稻的品质。然而, 稻蛙共作模式中蛙对稻田氮循环和土壤氨挥发的影响机理仍不清楚。采用长期定位研究, 设置不同的蛙投放密度, 结合盆栽试验分解研究蛙的扰动与蛙的排泄等行为对稻田生态系统氮循环产生的作用, 有利于全面评估稻蛙共作模式对稻田生态系统的影响。
4 结论
本研究采用密闭室间歇抽气法测得稻蛙共作模式中水稻季氨挥发累积量为47.02 kg∙hm−2, 占当季施氮量12.9%, 其后茬紫云英季的氨挥发累积量为16.27 kg∙hm−2, 全年轮作系统的氨挥发累积量为63.29 kg∙hm−2。与常规水稻种植模式相比, 稻蛙共作模式全年氨挥发累积量降低了15.3%, 其占该年施氮量的比例17.4%显著低于常规水稻种植模式所占比例20.5%。综上所述, 稻蛙共作模式在水稻-紫云英轮作系统中具备一定的氨减排潜力, 但该模式对稻田氨挥发影响的长期效应及其影响机理仍需进一步研究。
[1] LIAN Z M, OUYANG W, HAO F H, et al. Changes in fertilizer categories significantly altered the estimates of ammonia volatilizations induced from increased synthetic fertilizer application to Chinese rice fields[J]. Agriculture, Ecosystems & Environment, 2018, 265: 112−122
[2] GUO J H, LIU X J, ZHANG Y, et al. Significant acidification in major Chinese croplands[J]. Science, 2010, 327(5968): 1008−1010
[3] HUANG X, SONG Y, LI M M, et al. A high-resolution ammonia emission inventory in China[J]. Global Biogeochemical Cycles, 2012, 26(1): GB1030
[4] 王文林, 刘波, 韩睿明, 等. 农业源氨排放影响因素研究进展[J]. 生态与农村环境学报, 2016, 32(6): 870−878 WANG W L, LIU B, HAN R M, et al. Review of researches on factors affecting emission of ammonia from agriculture[J]. Journal of Ecology and Rural Environment, 2016, 32(6): 870–878
[5] JU X T, XING G X, CHEN X P, et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(9): 3041–3046
[6] DONG Y B, WU Z, ZHANG X, et al. Dynamic responses of ammonia volatilization to different rates of fresh and field-aged biochar in a rice-wheat rotation system[J]. Field Crops Research, 2019, 241: 107568
[7] HE T H, LIU D Y, YUAN J J, et al. A two years study on the combined effects of biochar and inhibitors on ammonia volatilization in an intensively managed rice field[J]. Agriculture, Ecosystems & Environment, 2018, 264: 44–53
[8] XIA L L, LI X B, MA Q Q, et al. Simultaneous quantification of N2, NH3and N2O emissions from a flooded paddy field under different N fertilization regimes[J]. Global Change Biology, 2020, 26(4): 2292–2303
[9] LIU X D, CHEN L Y, HUA Z L, et al. Comparing ammonia volatilization between conventional and slow-release nitrogen fertilizers in paddy fields in the Taihu Lake region[J]. Environmental Science and Pollution Research, 2020, 27(8): 8386–8394
[10] WEBB J, THORMAN R E, FERNANDA-ALLER M, et al. Emission factors for ammonia and nitrous oxide emissions following immediate manure incorporation on two contrasting soil types[J]. Atmospheric Environment, 2014, 82: 280–287
[11] LOURO A, SAWAMOTO T, CHADWICK D, et al. Effect of slurry and ammonium nitrate application on greenhouse gas fluxes of a grassland soil under atypical South West England weather conditions[J]. Agriculture, Ecosystems & Environment, 2013, 181: 1–11
[12] YI X M, YI K, FANG K K, et al. Microbial community structures and important associations between soil nutrients and the responses of specific taxa to rice-frog cultivation[J]. Frontiers in Microbiology, 2019, 10: 1752
[13] YI X M, YUAN J, ZHU Y H, et al. Comparison of the abundance and community structure of N-cycling bacteria in paddy rhizosphere soil under different rice cultivation patterns[J]. International Journal of Molecular Sciences, 2018, 19(12): 3772
[14] TENG Q, HU X F, LUO F, et al. Influences of introducing frogs in the paddy fields on soil properties and rice growth[J]. Journal of Soils and Sediments, 2016, 16(1): 51–61
[15] 林海雁, 黄倩霞, 邵紫依, 等. 养殖虎纹蛙稻田土壤酶活性及主要养分含量特征[J]. 核农学报, 2018, 32(4): 802–808 LIN H Y, HUANG Q X, SHAO Z Y, et al. Characterization of soil enzymatic activities and main nutrient contents in paddy field for eco-planting and breeding rice and frog[J]. Journal of Nuclear Agricultural Sciences, 2018, 32(4): 802–808
[16] 郭文啸, 赵琦, 朱元宏, 等. 蛙稻生态种养模式对土壤微生物特性的影响[J]. 江苏农业科学, 2018, 46(5): 57–60 GUO W X, ZHAO Q, ZHU Y H, et al. Effect of frog and rice ecological farming model on soil microbial characteristics[J]. Jiangsu Agricultural Sciences, 2018, 46(5): 57–60
[17] SHA Z M, CHU Q N, ZHAO Z, et al. Variations in nutrient and trace element composition of rice in an organic rice-frog coculture system[J]. Scientific Reports, 2017, 7(1): 15706
[18] 刘功朋, 张玉烛, 黄志农, 等. 水稻牛蛙生态种养对稻飞虱防效及水稻产量的影响[J]. 中国生物防治学报, 2013, 29(2): 207–213 LIU G P, ZHANG Y Z, HUANG Z N, et al. Effects of rice-bullfrog mixed cultivation on rice planthoppers and rice yield[J]. Chinese Journal of Biological Control, 2013, 29(2): 207–213
[19] FANG K K, YI X M, DAI W, et al. Effects of integrated rice-frog farming on paddy field greenhouse gas emissions[J]. International Journal of Environmental Research and Public Health, 2019, 16(11): 1930
[20] GAO H, SHA Z M, WANG F, et al. Nitrogen leakage in a rice-duck co-culture system with different fertilizer treatments in China[J]. Science of The Total Environment, 2019, 686: 555–567
[21] XU G C, LIU X, WANG Q S, et al. Integrated rice-duck farming mitigates the global warming potential in rice season[J]. Science of The Total Environment, 2017, 575: 58–66
[22] FREI M, BECKER K. Integrated rice-fish production and methane emission under greenhouse conditions[J]. Agriculture, Ecosystems & Environment, 2005, 107(1): 51–56
[23] LI C F, CAO C G, WANG J P, et al. Nitrogen losses from integrated rice-duck and rice-fish ecosystems in southern China[J]. Plant and Soil, 2008, 307(1/2): 207–217
[24] 罗喜秀, 李成芳, 陈灿, 等. 稻蟹共作模式生态效应研究进展[J]. 作物研究, 2019, 33(5): 352–355 LUO X X, LI C F, CHEN C, et al. Research status of ecological effect under the co-production mode of rice and crabs[J]. Crop Research, 2019, 33(5): 352–355
[25] 王昂, 马旭洲, 于永清, 等. 北方稻蟹共作系统氨挥发损失的研究[J]. 浙江农业学报, 2018, 30(4): 622–631 WANG A, MA X Z, YU Y Q, et al. Ammonia volatilization from rice-crab culture systems in northern China[J]. Acta Agriculturae Zhejiangensis, 2018, 30(4): 622–631
[26] 刘功朋. 不同蛙类及密度对水稻主要害虫、天敌及其产量的影响[D]. 长沙: 湖南农业大学, 2013: 11–16 LIU G P. Effects of different frog species and densities on main insect pests, natural enemies and rice yield[D]. Changsha: Hunan Agricultural University, 2013: 11–16
[27] KISSEL D E, BREWER B L, ARKIN G F. Design and test of a field sampler for ammonia volatilization[J]. Soil Science Society of America Journal, 1977, 41(6): 1133–1138
[28] 白志刚. 氮肥运筹对水稻氮代谢及稻田氮肥利用率的影响[D]. 杭州: 中国农业科学院(中国水稻研究所), 2019: 18 BAI Z G. Effects of N management strategy on N metabolism in rice plant and N use efficiency in paddy soil[D]. Hangzhou: Chinese Academy of Agricultural Sciences (China National Rice Research Institute), 2019: 18
[29] SUN H J, ZHANG H L, POWLSON D, et al. Rice production, nitrous oxide emission and ammonia volatilization as impacted by the nitrification inhibitor 2-chloro-6-(trichloromethyl)- pyridine[J]. Field Crops Research, 2015, 173: 1–7
[30] 陈宇眺. 栽培模式对水稻产量和氮肥利用率的影响及生理机制的研究[D]. 武汉: 华中农业大学, 2016: 62–63 CHEN Y T. Effects of cultivation patterns on rice yield and nitrogen use efficiency and its mechanisms[D]. Wuhan: Huazhong Agricultural University, 2016: 62–63
[31] 余姗, 薛利红, 花昀, 等. 水热炭减少稻田氨挥发损失的效果与机制[J]. 环境科学, 2020, 41(2): 922–931 YU S, XUE L H, HUA Y, et al. Effect of applying hydrochar for reduction of ammonia volatilization and mechanisms in paddy soil[J]. Environmental Science, 2020, 41(2): 922–931
[32] SUN H J, FENG Y F, XUE L H, et al. Responses of ammonia volatilization from rice paddy soil to application of wood vinegar alone or combined with biochar[J]. Chemosphere, 2020, 242: 125247
[33] LIN K M, WU J P. Effect of introducing frogs and fish on soil phosphorus availability dynamics and their relationship with rice yield in paddy fields[J]. Scientific Reports, 2020, 10(1): 21
[34] WYNGAARD N, CABRERA M L, SHOBER A, et al. Fertilization strategy can affect the estimation of soil nitrogen mineralization potential with chemical methods[J]. Plant and Soil, 2018, 432(1/2): 75–89
[35] WHALEN J K, CHANG C, CLAYTON G W, et al. Cattle manure amendments can increase the pH of acid soils[J]. Soil Science Society of America Journal, 2000, 64(3): 962–966
[36] WYNGAARD N, FRANKLIN D H, HABTESELASSIE M Y, et al. Legacy effect of fertilization and tillage systems on nitrogen mineralization and microbial communities[J]. Soil Science Society of America Journal, 2016, 80(5): 1262–1271
[37] 何一帆. 云南8种无尾两栖类呼吸和水分调节器官的环境适应性研究[D]. 昆明: 云南师范大学, 2018: 2–6 HE Y F. Studies on the environmental adaptability of respiratory and water regulation structures of 8 species of Anura Ampthibians in Yunnan Province[D]. Kunming: Yunnan Normal University, 2018: 2–6
[38] CHEN G, ZHAO G H, CHENG W D, et al. Rice nitrogen use efficiency does not link to ammonia volatilization in paddy fields[J]. Science of the Total Environment, 2020, 741: 140433
[39] 陈贵, 施卫明, 赵国华, 等. 太湖地区主栽高产水稻品种对土壤和肥料氮的利用特性研究[J]. 土壤, 2016, 48(2): 241–247 CHEN G, SHI W M, ZHAO G H, et al. Characteristics of utilization of n sources from soil and fertilizer by rice varieties with high yield in Taihu Lake Area[J]. Soils, 2016, 48(2): 241–247
[40] 郭文啸. 不同种植模式对稻田土壤养分和微生物特性的影响[D]. 上海: 上海交通大学, 2017: 38–41 GUO W X. Response of soil nutrient and microbiological characteristics to different rice cultivation patterns[D]. Shanghai: Shanghai Jiao Tong University, 2017: 38–41
[41] 鲁艳红, 廖育林, 聂军, 等. 稻-蛙生态种养技术模式研究与展望[J]. 湖南农业科学, 2017, (3): 74–76 LU Y H, LIAO Y L, NIE J, et al. Research and prospect of rice-frog ecological cultivation and breeding technology mode[J]. Hunan Agricultural Sciences, 2017, (3): 74–76
[42] LIA H, Felix B, INGE V D W, et al. Survival analysis of brown plant hoppers () in rice using video recordings of predation events[J]. Biological Control, 2018, 127: 155–161
Effect of rice-frog cultivation on ammonia volatilization in rice-Chinese milk vetch rotation system*
CHEN Huiyan1, SHA Zhimin1, WU Fujun1,2, FANG Kaikai1, XU Chunhua3, YANG Xiaolei3, ZHU Yuanhong4, CAO Linkui1**
(1. School of Agriculture and Biology, Shanghai Jiaotong University, Shanghai 200240, China; 2. College of Agriculture, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 3. Shanghai Agricultural Technology Extension and Service Center, Shanghai 201103, China; 4. Shanghai Qingpu Modern Agricultural Park Development Co., Ltd, Shanghai 201717, China)
Ammonia (NH3)volatilization is one of the main mechanisms of nitrogen loss in paddy fields. Studying the impact of the ecological cultivation model in paddy fields on ammonia volatilization can provide a theoretical basis for its ecological and environmental benefits. To evaluate the effects of rice-frog cultivation on ammonia volatilization in a rice-Chinese milk vetch (CMV) rotation system, the continuous airflow enclosure method was used to collect ammonia in a field plot experiment to study soil ammonia volatilization and its related factors. The experiment included three treatments: control check (CK, no fertilization, no frogs), conventional rice cultivation (CR, fertilization, no frogs), and rice-frog cultivation (RF, fertilization, frogs released). The results showed that the cumulative amount of ammonia volatilization in the rice-frog cultivation treatment was 47.02 kg·hm–2, accounting for 12.9% of the nitrogen application rate in the current season. The subsequent cumulative amount of ammonia volatilization in the Chinese milk vetch season was 16.27 kg·hm–2. The cumulative ammonia volatilization in the annual rotation system was 63.29 kg·hm–2, which was 15.3% lower than that of conventional rice planting. The cumulative amount of ammonia volatilization produced by rice-frog cultivation in the annual rice-Chinese milk vetch rotation system accounted for 17.4% of the annual nitrogen application, which was significantly lower than that of conventional rice cultivation (20.5%). The ammonium nitrogen concentration in the floodwater was the main factor affecting ammonia volatilization in the rice season, followed by the pH and temperature of the floodwater, air temperature, and wind speed. As the temperature increased, the influence of the ammonium nitrogen concentration in the floodwater on ammonia volatilization increased. Frogs did not affect the rice yield, rice yield components, nitrogen fertilizer efficiency, or Chinese milk vetch yield. Therefore, rice-frog cultivation has the potential to reduce ammonia in the rice-Chinese milk vetch rotation system, but the long-term effects of this model on ammonia volatilization in paddy fields and its mechanisms require further study.
Rice-Chinese milk vetch () rotation; Rice-frog cultivation; Ammonia volatilization; Yield
10.13930/j.cnki.cjea.200657
陈慧妍, 沙之敏, 吴富钧, 方凯凯, 徐春花, 杨晓磊, 朱元宏, 曹林奎. 稻蛙共作对水稻-紫云英轮作系统氨挥发的影响[J]. 中国生态农业学报(中英文), 2021, 29(5): 792-801
CHEN H Y, SHA Z M, WU F J, FANG K K, XU C H, YANG X L, ZHU Y H, CAO L K. Effect of rice-frog cultivation on ammonia volatilization in rice-Chinese milk vetch rotation system[J]. Chinese Journal of Eco-Agriculture, 2021, 29(5): 792-801
X171.3

* 上海市科技兴农推广项目[沪农科推字(2019)第2-1号]和国家自然科学基金项目(31770482)资助
曹林奎, 主要研究方向为生态农业与面源污染防控。E-mail: clk@sjtu.edu.cn
陈慧妍, 主要研究方向为稻田生态系统氮素循环。E-mail: huiyanchen@sjtu.edu.cn
2020-08-06
2020-11-12
* This study was supported by Shanghai Science and Technology Promotion Project [(2019) No. 2-1] and the National Natural Science Foundation of China (31770482).
, E-mail: clk@sjtu.edu.cn
Aug. 6, 2020;
Nov. 12, 2020
