利用生物信息学探究IL4 在哮喘中的异常表达及其相关基因的功能作用
2021-04-30卓超洲沈观乐雷朝君余瑞林梁杰高妩媚魏艾
卓超洲,沈观乐,雷朝君,余瑞林,梁杰,高妩媚,魏艾
(深圳市龙华区人民医院呼吸内科,广东深圳518109)
支气管哮喘是一种常见的疾病,我国成年人患病率约为1.24%[1]。并且随着工业化发展,哮喘的发病率也在逐年上升[2]。哮喘的发病是多种炎症细胞(嗜酸性粒细胞、肥大细胞等)共同参与、相互作用的结果[3]。白细胞介素家族是一类与免疫调节相关的细胞因子,在炎症反应中起重要作用。白细胞介素家族至少有40个成员,IL4则是其中一个[4]。目前已经有研究发现,哮喘患者的血清中IL4 水平显著上升,可能与气道炎症相关[5]。但是目前对IL4 在哮喘中的调控机制尚不清楚,因此在本研究中,通过对GEO 数据库中哮喘患者的转录组及甲基化数据进行挖掘,探究了IL4 及其相关基因在哮喘发生中的作用机制,发现IL4可能是治疗哮喘的潜在靶点。
1 材料与方法
1.1 数据来源及预处理
本研究所用到的测序数据均来源于公共数据库——GEO(Gene Expression Omnibus,https://www.ncbi.nlm.nih.gov/geo/)。哮喘患者的mRNA表达数据集来源于:GSE27011和GSE40888[6-7]。DNA甲基化数据来源于GSE40736[8]。miRNA 表达数据来源于GSE142237。LncRNA表达数据来源于GSE106230[9]。
1.2 差异表达lncRNA和miRNA分析
使用GSE106230 数据筛选哮喘患者显著高表达的lncRNA,GSE142237 数据集筛选miRNA 显著低表达的基因。差异表达分析用R 语言(version 3.6)的limma 包[10],log2foldchange>1,P<0.05 的lncRNA被认为是显著高表达。log2foldchange<-1,P<0.05的miRNA被认为是显著低表达。
1.3 CeRNA网络构建
IL4 与miRNA 互作信息来源于miRWalk(http://mirwalk.umm.uni-heidelberg.de/),miRNA 与lncRNA互作信息来源于StarBase3(http://starbase.sysu.edu.cn/)。IL4 相关的ceRNA 调控网络图用Cytoscape 进行展示[11]。
1.4 基因富集分析
将GSE27011 数据集中的哮喘患者按照IL4 的表达量进行从高到低排序。按照IL4 表达量的中位数将患者分为IL4 高表达组和低表达组。本研究用GSEA(version,4.0.3)软件对表达矩阵进行分析,采用c2.cp.kegg.v7.1.symbols.gmt 数据集,按照缺省参数设置进行基因富集分析(gene set enrichment analysis,GSEA),设定随机组合次数为1 000,|NES|>1,P<0.05的基因集被认为是显著富集的。
1.5 基因功能分析
本研究将与IL4 呈正相关和负相关的基因分别进行基因本体(GO)和京都基因与基因组百科全书(KEGG)分析。GO 分析用R 语言(version 3.6)的clusterProfiler 包[12]。KEGG 分 析 使 用 的 在 线 分 析KOBAS3(http://kobas.cbi.pku.edu.cn/)。P<0.05 被认为是显著富集的功能或者通路。
1.6 统计学处理
用R 语言(version 3.6)进行统计学分析。哮喘患者和正常人的IL4 表达量及甲基化水平比较采用独立样本秩和检验。基因表达之间的关系采用Pearson 相关性分析。P<0.05被认为有统计学意义。
2 结果
2.1 GEO数据样本统计
GSE27011包含36例哮喘患者和18例正常人的白细胞mRNA 表达数据。GSE40888 包含65 例哮喘患者和40 例正常人的外周血单核细胞mRNA 表达数据。GSE40736包含97例哮喘患者和97例正常人的外周血单核细胞DNA 甲基化数据。GSE142237包含8 例哮喘患者和4 例正常人的支气管上皮miRNA表达数据。GSE106230包含9例哮喘患者和3例正常人的外周血lncRNA表达数据。
2.2 喘患者和正常人IL4差异比较
通过将哮喘患者和正常人的IL4 表达水平进行比较,发现患者中IL4 显著高表达(P=0.042 8,图1A)。哮喘患者和正常人的IL4 DNA 甲基化水平进行比较,发现患者中IL4 DNA 甲基化水平显著降低(P=0.028 3,图1B)。
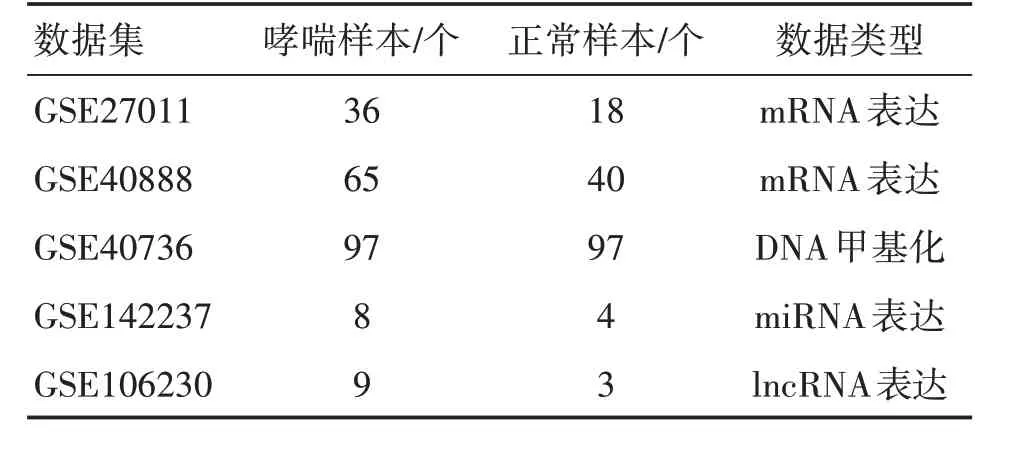
表1 数据集及样本分布情况Table 1 Data set and samples distribution
2.3 CeRNA网络构建
ceRNA 网 络 中,lncRNA 与mRNA 是 正 调 控 关系,miRNA与mRNA是负调控关系[13],因此本研究利用GSE142237 数据集筛选了在哮喘患者中显著下调的miRNA(50 个)。利用GSE106230 数据集筛选了在哮喘患者中显著上调的lncRNA(26 个)。通过预测IL4 与miRNA 的互作关系、miRNA 与lncRNA的互作关系,最终得到一个包含4个miRNA、6个lncRNA 和IL4 的ceRNA 调控网络(图2),表2 展示了这些lncRNA和miRNA的上调下调情况。
2.4 基因富集(GSEA)分析

图1 哮喘患者和正常人IL4差异比较结果。Figure 1 Difference of IL 4 between asthmatic patients and normal people
GSEA 分析发现有19 条通路显著富集在IL4 高表达患者中。如表3所示,高IL4患者富集的通路主要涉及代谢类通路和一些受体信号通路。对排名为前2的通路进行了展示(图3)。


表2 CeRNA网络中miRNA和lncRNA差异表达分析结果Table 2 Differential expression analysis of miRNA and lncRNA in ceRNA network
2.5 IL4相关基因筛选
利用GSE27011 和GSE40888 两个数据集中的哮喘患者基因表达矩阵,分别计算IL4 与其他基因表达的相关性。其中GSE27011 筛选到1 584 个显著与IL4表达相关的基因(816个正相关,768个负相关)。GSE40888 筛选到7 690 显著与IL4 表达相关的基因(5 119 个正相关,2 571 个负相关)。图4 和图5 分别对GSE27011 和GSE40888 2 个数据集相关性最强的前4 个基因进行了展示。将2 个数据集筛选到的相关基因取交集,一共得到210 个基因(144正相关,66负相关,图6)。

表3 IL4高表达患者显著富集的通路Table 3 Pathways enriched by IL4-overexpression patients

图3 IL4高表达患者富集的top2通路Figure 3 Top 2 pathways enriched by IL4-overexpression patients
2.6 IL4功能及通路分析
选取与IL4 正相关的基因集进行GO 和KEGG富集分析,以此来分析IL4 表达所促进的功能(图7A)或通路(图7C)。在分子功能(biological process,BP)方面,主要与一些蛋白结合或者磷酸酶活性有关,例如:蛋白激酶B结合、激活素结合、跨膜受体蛋白酪氨酸磷酸酶活性、跨膜受体蛋白磷酸酶活性等;在细胞成分(cellular component,CC)方面,参与构成一些细胞器,例如反式高尔基网囊泡的网格蛋白外套、跨高尔基网络转运囊泡膜、内溶酶体膜等;在生物学过程(molecular function,MF)方面,主要参与一些通路的调节作用,如:辅助性T-helper2 细胞分化的调控、前列腺素生物合成过程的调控、谷氨酸分泌的正调节等。在KEGG 通路方面,主要参与一些代谢通路,如甘油磷脂代谢、花生四烯酸代谢;一些重要物质的合成,如:氨酰-tRNA 生物合成、甾体激素生物合成、泛酸和辅酶A 生物合成和初级胆汁酸生物合成;另外还参与一写信号传导通路,如:mTOR 信号通路、幽门螺杆菌感染中的上皮细胞信号转导和甲状腺激素信号通路。选取与IL4 负相关的基因集进行富集分析,以此来分析IL4 表达所抑制的功能(图7B)或通路(图7D)。在分子功能方面,主要与一些蛋白结合或者磷酸酶活性有关,如泛素化类修饰依赖蛋白质结合、长春新碱结合、泛素-泛素连接酶活性、磷脂酰肌醇磷酸激酶活性和NAD 依赖性组蛋白脱乙酰酶活性(H3-K14 特异性)等。在细胞成分方面,参与构成DNA 复制因子A 复合体、核酸复制颗粒、树突棘膜、黑素体膜和壳质体等。在生物学过程方面,主要参与一些通路的调节作用,如:eIF2α 磷酸化对翻译起始的调控、ATF6 介导的未折叠蛋白反应、内质网蛋白输出的负调控和蛋白质多泛素化的负调控等。在KEGG 通路方面,主要参与一些基本生命活动的通路,如内质网的蛋白质加工、醛固酮合成与分泌、泛素介导的蛋白质水解和加压素调节的水重吸收和粘蛋白型O-聚糖生物合成等。

图4 GSE27011数据集中与IL4相关性最强的前四个基因。Figure 4 Strongest four genes correlated with IL4 in GSE27011 data set

图5 GSE40888数据集中与IL4相关性最强的前4个基因。Figure 5 Strongest four genes correlated with IL4 in GSE40888 data set
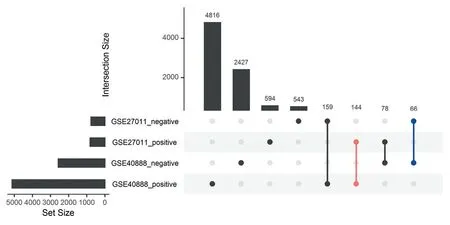
图6 GSE27011和GSE40888两个数据集中与IL4呈正相关和负相关基因交集Figure 6 Intersection of positive and negative correlation genes with IL4 between GSE27011 and GSE40888 data sets

图7 基因本体(GO)和京都基因与基因组百科全书(KEGG)分析Figure 7 Gene Ontology(GO)and Kyoto Encyclopedia of Genes(KEGG)analysis
3 讨论
DNA 的甲基化修饰可引起染色质结构改变,从而调控基因的表达,在疾病的发生发展中发挥重要的作用[14]。本研究中,通过比较哮喘患者和正常人的IL4 基因位点上DNA 甲基化水平,发现哮喘患者的IL4甲基化水平显著降低,而IL4在哮喘患者中的表达水平显著上升,因此推测,IL4 DNA 甲基化水平的降低促进了IL4的表达。
目前越来越多研究表明,lncRNA 可以与mRNA竞争性与miRNA 结合,从而调控mRNA 的表达[13]。这类调控机制的失衡可能导致疾病的发生。因此,本研究建立了IL4 相关的ceRNA 网络,以探究相关的调控机制。网络中的OIP5-AS1 已被报道可以作为哮喘的诊断标记物[15],hsa-miR-125b-5p 也被报道与IL4的表达具有一定的相关性[16]。网络中lncRNA和miRNA 的调控可能导致了哮喘患者IL4 的异常表达。
GSEA 分析发现高IL4 表达水平的患者富集的大多为代谢相关的通路,其中糖代谢、亚油酸、花生四烯酸、谷胱甘肽及各种氨基酸代谢已被报道过在哮喘患者中及健康对照中存在显著差异[17]。
通过对IL4 正相关的mRNA 进行通路分析,发现这些mRNA 所在的一些通路与哮喘发生相关,例如:花生四烯酸代谢通路被发现与哮喘急性发作相关[18];内吞作用,细胞膜上重要结构蛋白caveolin-1参与细胞的内吞作用,该蛋白也被报道可作用于中气道平滑肌细胞,与哮喘的发病相关[19];幽门螺杆菌感染中的上皮细胞信号转导通路,幽门螺杆菌感染被发现与儿童哮喘发病呈正相关性[20];甘油磷脂代谢通路,甘油磷脂被发现在哮喘组小鼠与对照组小鼠有显著差异[21];mTOR 信号通路在哮喘鼠模型中可参与肺组织炎性浸润和气道重塑等病理过程而介导哮喘的发生[22];甲状腺激素信号通路,甲状腺激素被报道可增强哮喘患者的气道平滑肌重构[23]。对于与IL4 呈负相关的mRNA,其富集的通路,也与哮喘相关,例如:自噬,自噬在支气管哮喘的发病过程中可能既有保护作用,又有损害作用[24];泛素介导的蛋白水解通路,paucigranulocytic 哮喘中发现一些显著高甲基化的基因,这些基因富集于泛素介导的蛋白水解通路[25]。
本研究通过应用生物信息学方法分析哮喘患者和正常人的转录组及甲基化数据,发现IL4 在哮喘中显著高表达,并探究了其异常表达的调控机制,同时也发现IL4 的高表达可能导致与哮喘相关的代谢异常。另外本研究还挖掘出与IL4 表达相关的mRNA,发现这些mRNA 可能通过调控一些相关通路参与哮喘的发病。本研究从多组学多角度研究了IL4 的异常表达及其相关基因对哮喘的调控作用,研究显示IL4可能可以作为哮喘治疗的靶点。
