基因组印记的研究进展
2021-04-29苏鲁方李振庭郑瑜包纯刘小云
苏鲁方 李振庭 郑瑜 包纯 刘小云

摘要 基因组印记(genomicimprinting),又称遗传印记(geneticimprinting),是基因表达的一种调控机制,属于表观遗传学,包括DNA甲基化修饰和组蛋白甲基化修饰。阐述了印记基因的基本特征(可逆性、成簇分布、时空性、组织特异性和保守性),其可能的形成机制,为研究动、植物遗传育种奠定了分子基础。
关键词 基因组印记;印记基因;甲基化;H3K27me3
中图分类号 Q.75文献标识码A文章编号0517-6611(2021)07-0008-04
AbstractGenomicimprintingalsoknownasgeneticimprintingisaregulatorymechanismofgeneexpressionwhichbelongstoepigenetics,includingDNAmethylationandhistonemethylationmodification.Thispaperillustratedthebasiccharacteristics(reversibility,clustereddistribution,temporalandspatial,tissuespecificityandconservativeproperty)ofimprintedgenesanditspossibleformativemechanism,inordertostudymoleculargeneticfoundationinanimalsandplants.
KeywordsGenomicimprinting;Imprintedgene;Methylation;H3K27me3
基因組印记(genomicimprinting)指子代中来自亲本的特定基因或染色体在发育过程中产生特异性的加工修饰导致父源或母源一方等位基因表达,另一方等位基因沉默或表达量很少的可遗传表观修饰现象。其中,母本等位基因表达,而父本等位基因不表达的基因称为母源表达基因或父源印记(paternalimprinting);父本等位基因表达而母本等位基因不表达的基因则称为父源表达基因或母源印记(maternalimprinting)[1-2]。因此印记基因具有亲源特异性。近年来,研究调节印记基因的可能主要机制包括:DNA甲基化(DNAmethylation)修饰、组蛋白甲基化修饰(histonemethylationmodification)、染色体重塑修饰(chromatinremodelingmodification)和长非编码RNA:lncRNA(longnoncodingRNA)等,这些修饰模式易受环境因素的影响。基因组印记主要发生在动物胎盘、胚胎和开花植物的胚乳中,任何一种调节方式发生改变都可能导致基因功能的变化。因此,基因组印记对动、植物正常生长发育和分化等生命活动有重要的作用。
1印记基因概念的提出
基因印记概念首次提出于1960年,Crouse[3]在研究Sciara昆虫试验中发现后代2条X染色体中仅来自母方的X染色体具有活性,父方的X染色体处于静止状态,这说明一些等位基因的表达出现差异性,具有亲本选择性。1970年,Kermicle[4]研究报道玉米植株存在能够影响后代籽粒胚乳颜色产生显著差异的印记基因(R基因),首次提出了印记基因可能存在于植物。1984年,Mcgrath等[5]利用核移植技术发现小鼠受精发育过程中,雄原核替代雌原核即两原核同时来自父方或雌原核替代雄原核即两原核来自母方,发育主要形成胚胎组织或胎盘组织,以致发育后期将会死亡,证明了亲本等位基因在功能上具有不同等性,同样首次提出了哺乳动物也可能存在基因印记现象。1991年,Dechiara等[6]通过基因敲除技术证实了小鼠的第一个内源性印记基因为胰岛素生长因子Ⅱ(insulinlikegrowthfactor2,IGF2),属于母源印记基因。研究发现若敲除的等位基因来自父本,后代表现为动物体型较小,若敲除来自母本的等位基因,后代表现正常。1991年,Bartolomei等[7-8]进一步证实胰岛素生长因子Ⅱ受体(insulinlikegrowthfactortype-2receptor,IGF2R)和长链非编码RNA(LncRNA)H19基因在小鼠中为父源印记基因。随后,Rainier等[9]首次发现人类存在的印记基因:IGF2和H19。目前,哺乳动物、植物中已分别有约150、16个印记基因被鉴定[10-11]。与哺乳动物相比,针对植物印记基因的研究不多,拟南芥中已鉴定出10个印记基因,其中具有印记现象的FIS(fertilisation-independentseed)基因是在研究一系列胚乳突变体中最早发现的,它们具有抑制中央细胞分裂和调节早期胚乳发育的功能,包括MEDEA(MEA)/FIS1、FIS2和FIS3/FIE等,其中MEA和FIS2为母源印记基因[11-13]。fis突变体有2种表型,一是在未授精情况下,中央细胞自主发育为二倍体胚乳;二是授精条件下,形成非细胞化胚乳,均导致胚乳发育异常[12-14]。这些基因中的部分不仅自身具有印记基因的表达特征,还可通过修饰组蛋白抑制基因表达的方式来调节其他印记基因[15]。
20世纪90年代,人类对哺乳动物基因组印记的分子研究进入了活跃时期。之后,随着科技进步和科研经费的投入,科学家们发现牛、猪、羊等家畜,有袋类动物[16]和种子植物[17]存在印记基因,并预测有500~1000个,已经证实的有160个左右[18-20],但两栖类、爬行类、鱼类和鸟类中尚未发现基因印记现象的存在[21]。
2印记基因的特征
2.1可逆性
哺乳动物基因组印记一般经历3个过程:印记的建立、维持、去除[22]。生殖细胞形成早期,来自亲本双方的印记全部去除;亲本等位基因在生殖细胞形成精子或卵子时产生新的印记;精子和卵子受精后的个体发育过程维持印记[2]。大多数印记基因在印记消除和建立的过程中,伴随着明显的DNA甲基化消除和形成,认为DNA甲基化可能是基因组印记建立的主要机制之一[23]。最近另一印记调控机制通过亲本等位基因差异性组蛋白修饰,影响亲本等位基因表达不同,发现由于母本等位基因组蛋白H3第27个赖氨酸发生三甲基化(H3K27me3)的修饰,父本等位基因未修饰,因而母本等位基因表达受抑制,父本等位基因得到表达[24]。哺乳动物原始生殖细胞到成熟雌雄生殖细胞完成了染色体组蛋白甲基化或DNA甲基化的除去和重编程过程,类似于哺乳动物印记基因组蛋白甲基化或DNA甲基化的除去和重编程过程。因此,可以认为哺乳动物原始生殖细胞在胚胎发育早期的过程中,雌雄生殖细胞首先完成基因组组蛋白或DNA的除修饰,再重新对基因组进行组蛋白修饰或DNA修饰,直到产生稳定修饰状态的精原细胞和卵原细胞[24]。因此,基因组的印记在雌雄生殖细胞发育期通过印记的消除和建立,形成稳定、遗传且可具有发育潜能的细胞(图1)。
植物与动物基因组印记过程类似,研究发现拟南芥存在与哺乳动物相似功能作用的DNA甲基化转移酶(Methyltransferase1,AtMET1)、多梳抑制蛋白复合体2(polycombrepressivecomplex2,PRC2),分别主要负责维持CpG位点胞嘧啶的甲基化、H3K27三甲基化/H3K9二甲基化/CHG位点胞嘧啶的甲基化[25-26]。目前,植物基因组印记现象只存在被子植物三倍体胚乳中,拟南芥胚乳整个基因组是去甲基化的,并伴随着广泛的小干扰RNA在非CpG位点的高甲基化[27]。DNA糖基化酶(demeter,DME)是完成胚乳印记基因去甲基化的一种酶,只在雌配体中央细胞中表达,雄配体中不表达[28]。可以推测,雌雄配体在中央细胞形成前在AtMET1控制下为基因组甲基化状态,使这些印记基因转录沉默。拟南芥雌性配体的中央细胞和胚乳特异性存在另一母系表达抑制蛋白FIS-PRC2,使母本基因染色体附近发生H3K27me3修饰,同时也存在H3K9me2修饰,而且有CHG位点发生胞嘧啶甲基化的现象,从而母本基因表达沉默,父本基因表达[26]。因此,认为植物雌雄配子基因组在发育过程中出现一个母源特异性机制,使胚囊中央细胞中默认的沉默状态基因激活或抑制某个基因的表达,母本等位基因得到表达或抑制,而父源等位基因缺乏这一机制,相应地选择沉默或表达。这也与哺乳动物形成雌雄配子时的基因组甲基化模式相一致,而植物在形成雌雄配子前基因组甲基化/去甲基化方式仍不清楚。
2.2成簇分布
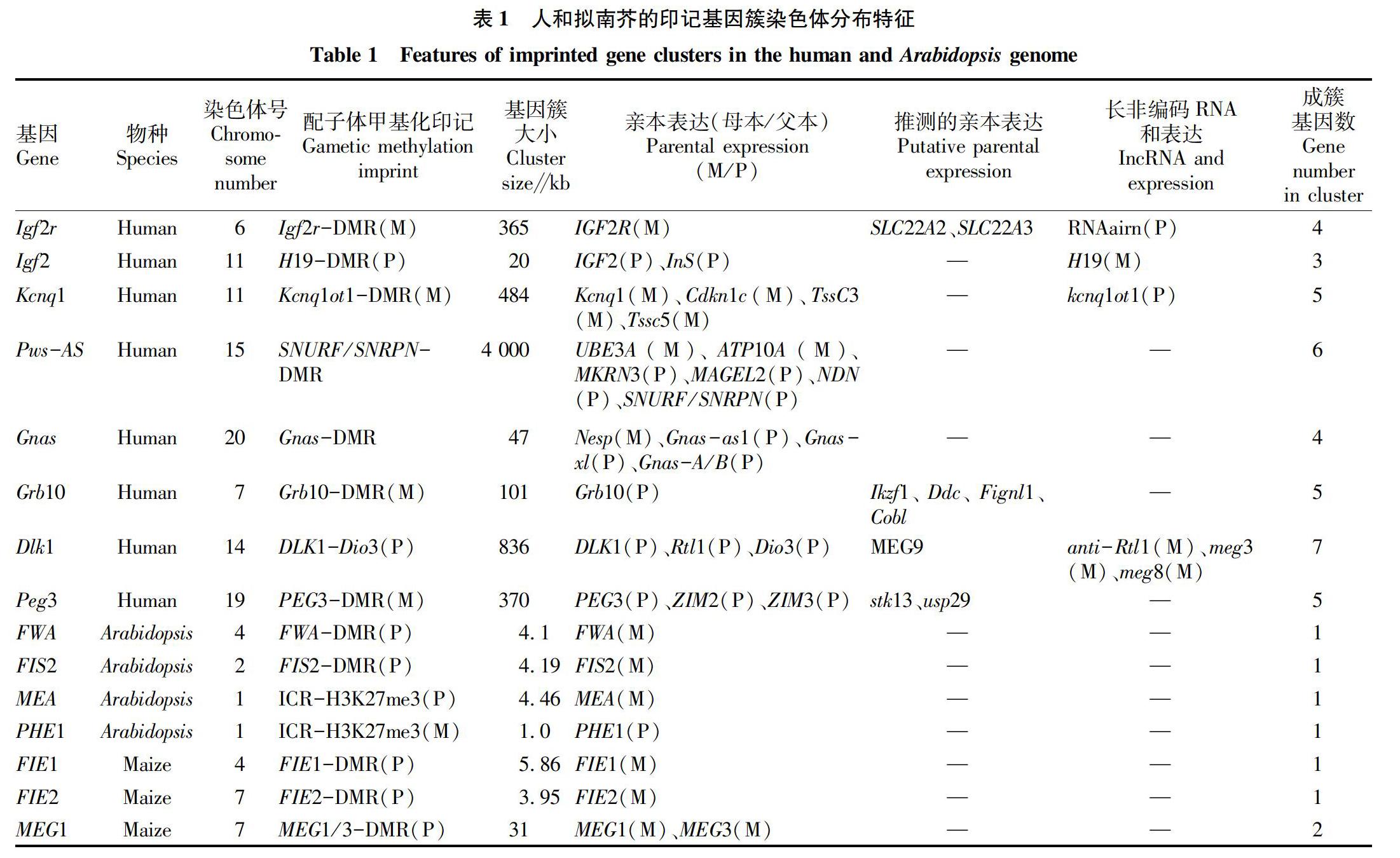
大多数印记基因的显著特征是在基因组中成簇分布,并构成印记域。一般情况下,父系印记基因和母系印记基因交替位于印记簇区域,它们翻译为蛋白质和转录为一些非编码RNA。人类和小鼠中已经发现100~200个印记基因,分别有70多个印记基因已证实,且约80%印记基因成簇存在于人和鼠基因组[29-30]。在小鼠2号、7号、9号、11号和17号染色体中发现较大的印记基因簇,这些印记基因簇包括IGF2、IGF2R、PWS、PEG3,DLK1、Grb10、Gna和Kcnq1印记域[31]。在人类7号、11号、14号、15号、17号、19号和20号染色体中发现7个较大的印记簇,分别位于人7q21、7q32、11p15、14q32、15q11-q13、19q13和20q13,另外这些印记簇包含基因较多(表1)[31]。
植物中发现的且已经证实的印记基因不多,主要发生在拟南芥和玉米中,无明显成簇分布现象,大多数印记基因为母系遗传表达即父系印记基因,印记方式有DNA甲基化和组蛋白H3K27me3。拟南芥中印记基因有MEDEA(MEA)/FIS1、FIS2、FIS3/FIE和FWA等,位于1號、2号和4号染色体中,且都为母源表达基因,在营养组织和胚中体现双等位基因表达[11-13,32]。在玉米的4号、7号和10号染色体上发现FIE1、FIE2、R、MEG1等印记基因,也都为母源表达基因[4,33-34]。小鼠中发现的依赖组蛋白甲基化修饰的印记基因无明显成簇现象,包括Grb10、Phf7、Smbt、2Slc38a4和smoc1,这些都为父源表达等位基因[24]。
2.3时空性、组织特异性和保守性
基因的印记遗传具有时空性,即不同发育阶段基因遗传印记表现不同。如鼠在交配后7.5d的滋养层细胞中Mash2基因表现为双亲基因的等位表达,即双等位基因表达,而8.5d之后则表现为只表达母方等位基因的遗传特性[35],即单等位表达;另外,鼠的Igf2r基因在妊娠期的第4.5天,胚胎未植入,所有细胞表现为双亲等位基因的表达,但到妊娠期的第6.5天至第13.5天,此时,Igf2r在已植入的胚胎细胞中仅表达母方等位基因[36]。
基因的印记遗传具有组织特异性和物种差异性。在1月龄猪中MEST基因在心、胃、肌肉、肾脏、肺、膀胱、舌头和脂肪中,MEST表达为父源等位基因即为母源印记,而在肝脏、小肠和脾中为双等位表达[37];人MEST基因有2种剪接体形式存在,分别是MEST1和MEST2,在所有组织中MEST1基因剪接体只表达父本的等位基因,而剪接体MEST2在心、脑、脾、胃、肾上腺等11个组织表达双亲等位基因,在胎儿的肾和胎盘组织只表达父方等位基因[38-39]。
印记基因的遗传具有保守性。如在小鼠中发现印记基因,人类这些基因也被证实具有类似的印记现象,发现有100~200个印记基因[29],说明物种间印记基因遗传具有保守性。目前为止,人和鼠中已被鉴定的印记基因分别约为45个和74个,其中在人和小鼠中共同鉴定的印记基因有34个[29]。在这34个人和小鼠同时鉴定的印记基因中,人和小鼠中印记方向一致有26个基因,2个基因表现为印记的组织和发育阶段特异性,6个基因表现为印记方向不同或一方印记而另一方非印记[29]。
在植物拟南芥和玉米中证实的印记基因不多,印记现象只发现在胚乳组织中具有组织印记特异性。拟南芥中FIS和FWA基因只在胚乳中表现印记现象,且都只表现为母方等位基因[11-13,32];玉米中FIE1、FIE2和R基因也都在胚乳组织中表达母本等位基因,胚中无此现象发生[4,33-34]。
因此,哺乳动物印记基因的表达具有时空性、组织特异性和保守性,而目前发现的植物印记基因不多,印记基因表现的特点为组织特异性,还未发现印记基因应有的其他特点。
3基因组印记与生长发育
基因组印记现象与动植物的生长发育、疾病、哺乳行为和重要经济性状息息相关。PHLDA2(TssC3)是第一个与细胞凋亡相关的印记基因[40],在人和鼠中分别位于11p15和7号染色体,其母源等位基因表达方式相继在人、鼠、牛、猪中得到证实,大量表达于胎盘滋养细胞层细胞,对调控胎盘和胎儿生长发育方面有着重要功能[41-43]。Igf2-Cdkn1印记域在人和鼠中基因序列和印记表达状态高度保守,这些基因是最早发现的印记基因,调控机体的发育和分化,已证实贝金威思-威德曼综合征(Beckwith-Wiedemannsyndrome,BWS)与该印记域内印记基因的突变和缺失相关[44]。人的15q11-q13印记域包含SNRPN、IPW、ZNF127、NDN的父源表达基因和UBE3A的母源表达基因,该区域印记状态的变化受SNRPN上游印记中心(imprintingcentre,IC)微缺失和突变影响,从而导致天使综合征(angelmansyndrome,AS)和普拉德-威利综合征(Prader-Willisyndrome,PWS)的发生[45-49]。鼠的父源表达基因PEG3被证实有养育行为的功能,与人的PEG3基因同源性高度相似,也为父源表达基因,它们都在脑组织中表达量高,推测可能同样具有哺育后代行为的功能[50-51]。家畜中存在很多调控性状的印记基因,包括绵羊的后群肌肉肥大母系印记基因callipyge[52]、调控蒙古羊胸椎数量的母系印记基因Homebox[53]和调控猪背膘厚度、眼肌高度和肌内脂肪含量的印记QTL[54]。在植物拟南芥中发现的一系列FIS印记基因具有调控中央细胞分裂和调节早期胚乳发育的功能,相应的突变体表现为胚和胚乳发育不全现象[15]。
4展望
基因組印记现象广泛存在于哺乳动物和开花植物体内,对动植物的正常生长和发育发挥着重要作用,包括动物胚胎、胎盘的发育和生长以及开花植物种子胚乳的发育。基因组的印记还与人的肿瘤形成相关,而且基因组印记与DNA的甲基化或组蛋白的修饰因果关系尚不明确,深入研究基因组印记机制,将会为肿瘤的诊断和治疗提供新的方法。目前,基因组印记现象存在于植物种子胚乳发育的过程中,这种方式也可能参与植物抗逆境胁迫的机理,可能包括DNA甲基化、组蛋白甲基化或乙酰化或磷酸化或泛素化和IncRNAs(longnoncodingRNAs)以及染色质重塑等,这些修饰对基因组印记调控起着重要作用。
参考文献
[1]周泉勇,唐艳强,赵书红.猪印记基因的研究现状及其应用前景[J].猪业科学,2010,27(7):30-33.
[2]REIKW,WALTERJ.Genomicimprinting:Parentalinfluenceonthegenome[J].NatRevGenet,2001,2(1):21-32.
[3]CROUSEHV.ThecontrollingelementinsexchromosomebehaviorinSciara[J].Genetics,1960,45(10):1429-1443.
[4]KERMICLEJL.DependenceoftheRmottledaleuronephenotypeinmaizeonmodeofsexualtransmission[J].Genetics,1970,66(1):69-85.
[5]MCGRATHJ,SOLTERD.Completionofmouseembryogenesisrequiresboththematernalandpaternalgenomes[J].Cell,1984,37(1):179-183.
[6]DECHIARATM,ROBERTSONEJ,EFSTRATIADISA.ParentalimprintingofthemouseinsulinlikegrowthfactorIIgene[J].Cell,1991,64(4):849-859.
[7]BARTOLOMEIMS,ZEMELS,TILGHMANSM.ParentalimprintingofthemouseH19gene[J].Nature,1991,351(6322):153-155.
[8]BARLOWDP,STGERR,HERRMANNBG,etal.Themouseinsulinlikegrowthfactortype2receptorisimprintedandcloselylinkedtotheTmelocus[J].Nature,1991,349(6304):84-87.
[9]RAINIERS,JOHNSONLA,DOBRYCJ,etal.Relaxationofimprintedgenesinhumancancer[J].Nature,1993,362(6422):747-749.
[10]SCOTTRJ,SPIELMANM.Deeperintothemaize:Newinsightsintogenomicimprintinginplants[J].Bioessays,2006,28(12):1167-1171.
[11]BERGERF,CHAUDHURYA.Parentalmemoriesshapeseeds[J].TrendsPlantSci,2009,14(10):550-556.
[12]KINOSHITAT,YADEGARIR,HARADAJJ,etal.ImprintingoftheMEDEApolycombgeneintheArabidopsisendosperm[J].PlantCell,1999,11(10):1945-1952.
[13]BERGERF,GRINIPE,SCHNITTGERA.Endosperm:Anintegratorofseedgrowthanddevelopment[J].CurrOpinPlantBiol,2006,9(6):664-670.
[14]LUOM,BILODEAUP,DENNISES,etal.ExpressionandparentoforigineffectsforFIS2,MEA,andFIEintheendospermandembryoofdevelopingArabidopsisseeds[J].PNAS,2000,97(19):10637-10642.
[15]SCOTTRJ,SPIELMANM.Deeperintothemaize:Newinsightsintogenomicimprintinginplants[J].Bioessays,2006,28(12):1167-1171.
[16]KILLIANJK,BYRDJC,JIRTLEJV,etal.M6P/IGF2Rimprintingevolutioninmammals[J].MolCell,2000,5(4):707-716.
[17]ALLEMANM,DOCTORJ.Genomicimprintinginplants:Observationsandevolutionaryimplications[J].PlantMolBiol,2000,43(2/3):147-161.
[18]NIKAIDOI,SAITOC,MIZUNOY,etal.Discoveryofimprintedtranscriptsinthemousetranscriptomeusinglargescaleexpressionprofiling[J].GenomeRes,2003,13(6B):1402-1409.
[19]LUEDIPP,HARTEMINKAJ,JIRTLERL.Genomewidepredictionofimprintedmurinegenes[J].GenomeRes,2005,15(6):875-884.
[20]GLASERRL,RAMSAYJP,MORISONIM.Theimprintedgeneandparentoforigineffectdatabasenowincludesparentaloriginofdenovomutations[J].NucleicAcidsRes,2006,34:D29-D31.
[21]REIKW,SANTOSF,DEANW.Mammalianepigenomics:Reprogrammingthegenomefordevelopmentandtherapy[J].Theriogenology,2003,59(1):21-32.
[22]BENDERJ.DNAmethylationandepigenetics[J].AnnuRevPlantBiol,2004,55(1):41-68.
[23]BRANDEISM,KAFRIT,ARIELM,etal.Theontogenyofallelespecificmethylationassociatedwithimprintedgenesinthemouse[J].EMBOJ,1993,12(9):3669-3677.
[24]INOUEA,JIANGL,LUF,etal.MaternalH3K27me3controlsDNAmethylationindependentimprinting[J].Nature,2017,547(7664):419-424.
[25]JULLIENPE,KINOSHITAT,OHADN,etal.MaintenanceofDNAmethylationduringtheArabidopsislifecycleisessentialforparentalimprinting[J].PlantCell,2006,18(6):1360-1372.
[26]CAOR,ZHANGY.ThefunctionsofE(Z)/EZH2mediatedmethylationoflysine27inhistoneH3[J].CurrOpinGenetDev,2004,14(2):155-164.
[27]HSIEHTF,IBARRACA,SILVAP,etal.GenomewidedemethylationofArabidopsisendosperm[J].Science,2009,324(5933):1451-1454.
[28]CHOIY,GEHRINGM,JOHNSONL,etal.DEMETER,aDNAglycosylasedomainprotein,isrequiredforendospermgeneimprintingandseedviabilityinArabidopsis[J].Cell,2002,110(1):33-42.
[29]FALLSJG,PULFORDDJ,WYLIEAA,etal.Genomicimprinting:Implicationsforhumandisease[J].AmJPathol,1999,154(3):635-647.
[30]WANLB,BARTOLOMEIMS.Regulationofimprintinginclusters:NoncodingRNAsversusinsulators[J].AdvGenet,2008,61:207-223.
[31]MORISONIM,RAMSAYJP,SPENCERHG.Acensusofmammalianimprinting[J].TrendsGenet,2005,21(8):457-465.
[32]KINOSHITAT,MIURAA,CHOIY,etal.OnewaycontrolofFWAimprintinginArabidopsisendospermbyDNAmethylation[J].Science,2004,303(5657):521-523.
[33]GUTIRRZMARCOSJF,COSTALM,DALPRM,etal.Epigeneticasymmetryofimprintedgenesinplantgametes[J].NatGenet,2006,38(8):876-878.
[34]GUTIRREZMARCOSJF,COSTALM,BIDERREPETITC,etal.Maternallyexpressedgene1isanovelmaizeendospermtransfercellspecificgenewithamaternalparentoforiginpatternofexpression[J].PlantCell,2004,16(5):1288-1301.
[35]GUILLEMOTF,CASPARYT,TILGHMANSM,etal.GenomicimprintingofMash2,amousegenerequiredfortrophoblastdevelopment[J].NatGenet,1995,9(3):235-242.
[36]LERCHNERW,BARLOWDP.PaternalrepressionoftheimprintedmouseIgf2rlocusoccursduringimplantationandisstableinalltissuesofthepostimplantationmouseembryo[J].MechDev,1997,61(1/2):141-149.
[37]李顺,蒋曹德,石萍,等.猪COPG2和MEST克隆、印记状况和组织表达分析[J].中国农业科学,2010,43(23):4901-4909.
[38]KOSAKIK,KOSAKIR,CRAIGENWJ,etal.IsoformspecificimprintingofthehumanPEG1/MESTgene[J].AmJHumGenet,2000,66(1):309-312.
[39]MCMINNJ,WEIM,SADOVSKYY,etal.ImprintingofPEG1/MESTisoform2inhumanplacenta[J].Placenta,2006,27(2/3):119-126.
[40]LEEMP,FEINBERGAP.Genomicimprintingofahumanapoptosisgenehomologue,TSSC3[J].CancerRes,1998,58(5):1052-1056.
[41]QIANNF,FRANKD,O′KEEFED,etal.TheIPLgeneonchromosome11p15.5isimprintedinhumansandmiceandissimilartoTDAG51,implicatedinFasexpressionandapoptosis[J].HumMolGenet,1997,6(12):2021-2029.
[42]SIKORAKM,MAGEEDA,BERKOWICZEW,etal.PHLDA2isanimprintedgeneincattle[J].AnimGenet,2012,43(5):587-590.
[43]HOUSP,CHENYM,LIANGJ,etal.DevelopmentalstagespecificimprintingofIPLindomesticpigs(Susscrofa)[J].JBiomedBiotechnol,2010,2010:1-6.
[44]OHLSSONR,NYSTMA,PFEIFEROHLSSONS,etal.IGF2isparentallyimprintedduringhumanembryogenesisandintheBeckwithWiedemannsyndrome[J].NatGenet,1993,4(1):94-97.
[45]WEVRICKR,KERNSJA,FRANCKEU.IdentificationofanovelpaternallyexpressedgeneinthePraderWillisyndromeregion[J].HumMolGenet,1994,3(10):1877-1883.
[46]GLENNCC,NICHOLLSRD,ROBINSONWP,etal.Modificationof15q11q13DNAmethylationimprintsinuniqueAngelmanandPraderWillipatients[J].HumMolGenet,1993,2(9):1377-1382.
[47]REEDML,LEFFSE.MaternalimprintingofhumanSNRPN,agenedeletedinPraderWillisyndrome[J].NatGenet,1994,6(2):163-167.
[48]SUTCLIFFEJS,HANM,CHRISTIANSL,etal.Neuronallyexpressednecdingene:AnimprintedcandidategeneinPraderWillisyndrome[J].Lancet,1997,350(9090):1520-1521.
[49]ALBRECHTU,SUTCLIFFEJS,CATTANACHBM.ImprintedexpressionofthemurineAngelmansyndromegene,Ube3a,inhippocampalandPurkinjeneurons[J].NatGenet,1997,17(1):75-78.
[50]KUROIWAY,KANEKOISHINOT,KAGITANIF,etal.Peg3imprintedgeneonproximalchromosome7encodesforazincfingerprotein[J].NatGenet,1996,12(2):186-190.
[51]KIMJ,ASHWORTHL,BRANSCOMBE,etal.Thehumanhomologofamouseimprintedgene,Peg3,mapstoazincfingergenerichregionofhumanchromosome19q13.4[J].GenomeRes,1997,7(5):532-540.
[52]KOOHMARAIEM,SHACKELFORDSD,WHEELERTL,etal.Amusclehypertrophyconditioninlamb(callipyge):Characterizationofeffectsonmusclegrowthandmeatqualitytraits[J].JAnimSci,1995,73(12):3596-3607.
[53]張立岭,菊林花,杨丽君.蒙古羊胸椎数的亲本印记遗传研究[J].内蒙古农业大学学报(自然科学版),2000,21(2):1-6.
[54]DEKONINGDJ,RATTINKAP,HARLIZIUSB,etal.Genomewidescanforbodycompositioninpigsrevealsimportantroleofimprinting[J].PNAS,2000,97(14):7947-7950.
