γ-聚谷氨酸对冻藏谷蛋白水合及结构的影响
2021-04-22谢新华毋修远仵心军郝明远范逸超邢彩云薛振丹
谢新华,毋修远,仵心军,郝明远,范逸超,邢彩云,薛振丹
(1 河南农业大学食品科学技术学院 郑州450002 2 郑州市食品药品检验所 郑州450003)
小麦面筋蛋白又称谷朊粉,被认为是控制小麦制品的最重要的组成部分,并对其加工性能起主要作用[1]。谷蛋白和麦醇溶蛋白是小麦面筋蛋白中两种主要蛋白,占小麦总蛋白的85%以上,其充分吸水可形成蛋白网络结构,使其面筋蛋白具有弹性及黏性等特质,并直接影响面制品品质[2]。谷蛋白是由多个亚基通过链外二硫键聚合形成的聚合体蛋白,分子质量约40~300 ku,其中包含低分子质量(LMW)和高分子质量(HMW)谷蛋白亚基,同时谷蛋白在控制面团弹性和强度方面起着关键作用[3]。现阶段有研究表明,冷冻面团中面筋蛋白的劣变可能是谷蛋白聚合物的复杂结构和分子质量分布以及因冷冻引起的构象重排和水再分配等原因所致。先前的研究也表明冻藏期间谷蛋白大分子(GMP)会出现不同程度的解聚现象[4],从而影响面团特性及其制品品质[5]。
γ-聚谷氨酸(γ-PGA)是由D-谷氨酸和L-谷氨酸通过α-氨基和γ-羧基聚合而成的直链氨基酸聚合物,因具有良好的增稠性、吸水性、抗冻性及可食用性等特点而被广泛应用于食品工业。Lim等[6]在美式甜甜圈中添加γ-PGA 使得甜甜圈吸油量降低5 倍左右,且孔隙度及感官等多方面体现出更好的状态。姬晓月等[7]的研究发现γ-PGA 可减少可冻结水的增加,阻碍淀粉结晶,从而保护冷冻面团微观结构,使煮后的速冻水饺皮具有较佳的质构特性,改善速冻水饺的感官特性,γ-PGA还可抑制面筋蛋白在冻藏期间的劣变[8]。为明确γ-PGA 在冻藏期间在面筋蛋白中的作用机制,本试验通过核磁共振仪器 (NMR)、傅里叶红外仪(FITR)、差示扫描量热仪 (DSC)、动态流变仪(DHR)和扫描电子显微镜(SEM)测定冻藏过程中谷蛋白的保水力、水分子分布、蛋白二级结构、热力学及流变学特性、微观结构,研究γ-PGA 在冻藏期间对谷蛋白水合及结构的影响,为探究γ-PGA 在减缓面筋蛋白冻藏中劣变机理提供基础数据。
1 材料与方法
1.1 材料
面筋蛋白,购自一加一天然面粉有限公司,蛋白质含量82%;γ-聚谷氨酸,购自西安四季生物科技有限公司。
1.2 仪器与设备
Flexi-Dry 冷冻干燥机,美国FTSSYSTEM 公司;DHR-2 旋转流变仪,美国TA 仪器有限公司;Micro MR 型核磁共振仪,上海纽迈电子科技有限公司;QUANTAFEG250 型场发射扫描电镜,美国FEI 公司;TENSOR Ⅱ型傅里叶红外光谱仪(FITR),布鲁克科技有限公司;DSC-214 型差示扫描量热仪,耐驰科学仪器商贸(上海)有限公司。
1.3 方法
1.3.1 样品制备及冻藏 采用Wang 等[9]的方法制得谷蛋白,将其冷冻干燥后粉碎备用。将1 g 谷蛋白粉与1.5 mL 的水均匀混合(谷蛋白与水按4∶6 质量比混合),整个混合过程为避免蛋白局部结块现象采用混漩涡振荡的方式。试验设置空白组和γ-PGA 组,γ-PGA 组中加入占谷蛋白质量1%的γ-PGA。将制得的湿谷蛋白块置于4 ℃保温箱平衡1 h 后于-40 ℃低温速冻2 h,后置于-18 ℃下冻藏0,1,3,5 和7 周,即为冷冻谷蛋白。在冻藏不同周期后,部分样品于室温解冻2 h 后直接测定,部分样品采用冷冻干燥处理后经粉碎、研磨,过120 目筛后放置于干燥器中备用。
1.3.2 谷蛋白保水力测定
谷蛋白保水力计算公式为:M=m1-m2
式中M——谷蛋白保水力,g;m1——不同冻藏周期的谷蛋白质量,g;m2——不同冻藏周期谷蛋白在105 ℃烘箱内烘至恒重后质量,g。
1.3.3 谷蛋白中水分分布测定 谷蛋白弛豫时间T2测定参考Zhang 等[10]的方法稍作修改,参数设置:共振频率22 MHz,磁体温度32 ℃,90°脉冲时间P90 为18 μs,180°脉冲时间P180 为36 μs,采样点数TD 为416 616,累加次数NS 为32,回波个数NECH 为5 000,数字增益DRG 为3。
1.3.4 谷蛋白二级结构测定 采用傅里叶红外仪对蛋白二级结构进行测定,参数设置:扫描范围400~4 000 cm-1;扫描256 次;分辨率4 cm-1。其中1 615~1 637 cm-1和1 682~1 700 cm-1为β-折叠;1 646~1 664 cm-1为α-螺旋;1 637~1 645 cm-1为无规卷曲;1 664~1 681 cm-1为β-转角[11]。
1.3.5 谷蛋白热力学特性测定 准确称取5 mg谷蛋白冻干后的样品,密封在DSC 专用坩埚内,以空坩埚作为对照。程序设定为:初始温度20 ℃,升温速度5 ℃/min,最终温度100 ℃,面筋蛋白变性起始温度(T0)、变性峰值温度(Tp)及焓变(ΔH)通过耐驰公司系统软件分析计算。
1.3.6 谷蛋白流变学特性测定 将冻藏不同周期的谷蛋白于室温下解冻2 h 后,采用旋转流变仪测定动态流变学特性。平板直径为40 mm,测量间隙为1.2 mm,下压平板后用塑料刀对多余谷蛋白进行切除修边处理,表面涂硅油防止水分挥发。扫描频率范围为0.01~100 Hz,应力设置为1%,测定温度为25 ℃[10]。
1.3.7 谷蛋白微观结构观察 取冷冻干燥后样品的中心部位,制成5 mm×5 mm×5 mm 体积后表面喷金,在扫描电镜进行1 000 倍观察。
1.3.8 数据分析 用SPSS 22.0 对数据进行统计分析。
2 结果与分析
2.1 γ-PGA 对谷蛋白保水力的影响
γ-PGA 对谷蛋白保水力影响如图1所示。可知谷蛋白未冻藏时,其添加1%的γ-PGA 能有效提升其保水能力。随着冻藏时间延长,两组均呈现保水力下降,空白组在冻藏0 至1 周期间保水力急剧下降至0.003 g 左右,之后下降趋势趋于平缓。而γ-PGA 组的谷蛋白在冻藏7 周后其保水力仅下降了0.015 g 左右,相比空白组保留了更多水分。这是因为γ-PGA 中大量羧基使其具有极强的吸水性,冻藏期间很好地锁住了水分,抑制了水分流失[12]。Shyu 等[13]通过在面团中添加γ-PGA 得知其可有效增强面团持水性能,在后续研究中发现面筋蛋白持水能力随γ-PGA 浓度的增大而提升。试验结果表明γ-PGA 可促使谷蛋白保留更多水分。
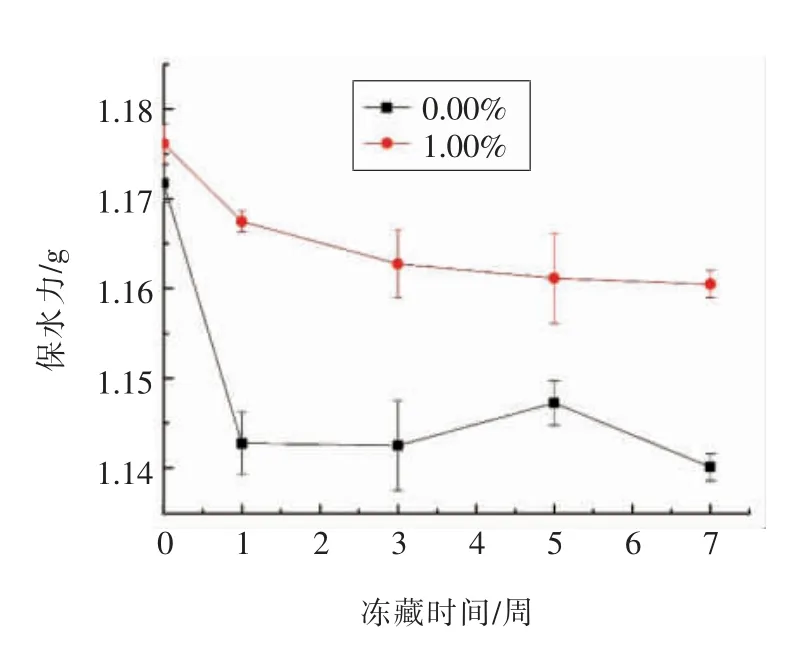
图1 γ-PGA 对谷蛋白保水力的影响Fig.1 Effect of γ-PGA on the water retention of glutenin
2.2 γ-PGA 对谷蛋白水分分布的影响
表1为γ-PGA 对谷蛋白水分子弛豫时间(T21、T22、T23)和对应峰面积的影响,谷蛋白中水分子是由结合水T21、弱结合水T22和自由水T23三部分水分组成。可知随冻藏时间增加,空白组中弱结合水呈下降趋势,而自由水含量却由6.7%左右显著增大至8.2%左右,这是因为冻藏期间冰晶的重结晶现象使得谷蛋白之间的交联作用减弱,导致蛋白与水的结合能力降低[14],引发部分弱结合水向自由水转化。而在γ-PGA 组,发现冻藏初期其自由水含量比空白组小2.5%,而弱结合水相对含量却变大,表明γ-PGA 的添加改变了谷蛋白的水分分布,较少的自由水使其更加稳定。同时γ-PGA 有效抑制了谷蛋白冻藏期间的水分迁移,冻藏7 周后其自由水含量较空白组低3.5%。对比γ-PGA 对面筋蛋白冻藏期间水分变化影响[8],试验发现谷蛋白与面筋蛋白冻藏期间弱结合水和自由水的变化趋势一致,由此表明γ-PGA 通过抑制谷蛋白的水分迁移从而影响了面筋蛋白冻藏期间的水分迁移。
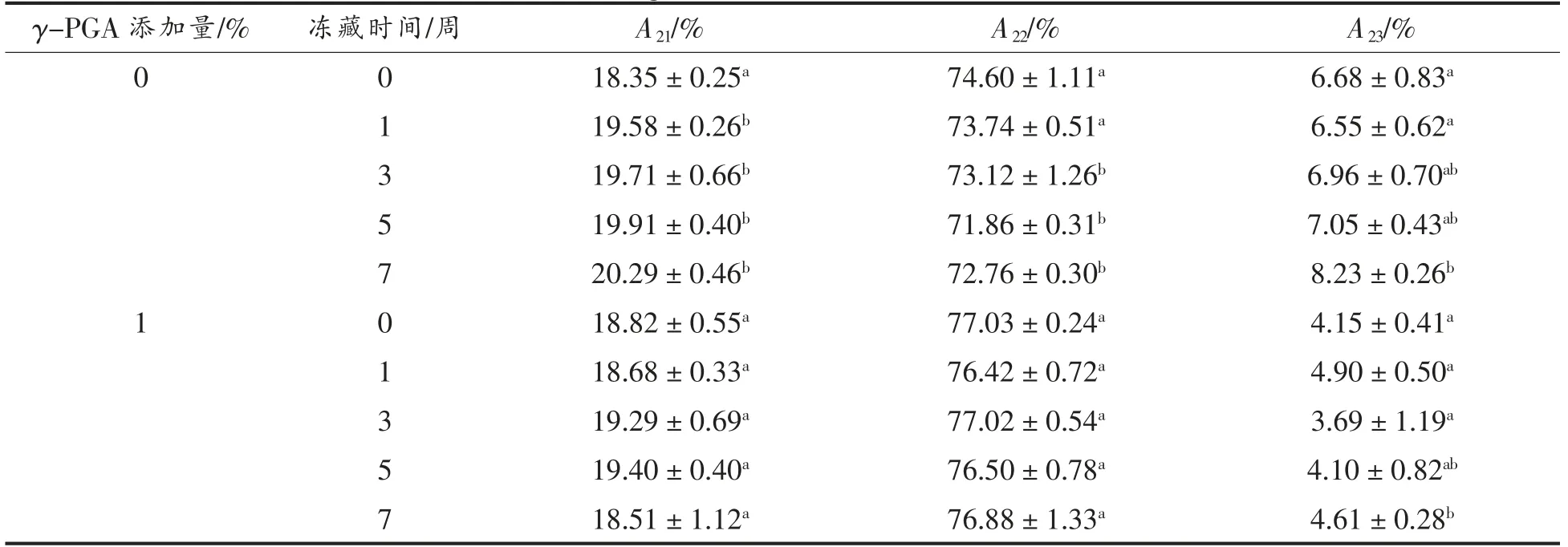
表1 γ-PGA 对谷蛋白水分子弛豫时间(T21、T22、T23)和对应峰面积的影响Table 1 Effect of γ-PGA on the relaxation time (T21,T22,T23)and corresponding peak area of glutenin water molecules
2.3 γ-PGA 对谷蛋白二级结构的影响
表2为γ-PGA 对谷蛋白二级结构中各成分相对含量的影响,谷蛋白中α-螺旋、β-折叠、β-转角、无规则卷曲的相对含量分别为25.5%,40.8%,21.1%,12.5%。随冻藏时间延长,γ-PGA 组β-折叠相对含量由41.4%下降至37.4%左右,无规则卷曲相对含量显著增加1.5%。二级结构的改变一方面可能是因为随着冻藏时间的延长,谷蛋白中重结晶现象使得谷蛋白大分子因机械力而解聚,使其有序状态向无序状态转化,另一方面是因为冻藏导致蛋白分子间氢键的破坏,使得亲水与疏水基团暴露引发的蛋白分子间和分子内部的重新交联[15]。与空白相比,蛋白二级结构中作为无序结构的β-转角、无规则卷曲的增大幅度减缓,较为稳定的有序状态的α-螺旋、β-折叠降低幅度减小也表明γ-PGA 可以起到冰晶修饰作用,减少大冰晶的形成,从而保护谷蛋白大分子,削弱冻藏导致的有序向无序转化。
2.4 γ-PGA 对谷蛋白热力学的影响
变性温度(Tp)和焓变(ΔH)是蛋白在热变性过程中的主要参数,蛋白样品热力学特性如表3所示,因小麦蛋白对热敏感性较低,故只能看到较小的吸收峰值。可知随冻藏时间增加,谷蛋白的变性起始温度显著升高6.3 ℃,其变性温度Tp也由58.5 ℃显著升高至61.3 ℃,表明其低温稳定性升高,这可能与冻藏导致其疏水基团的暴露有关[16],同时其热焓值显著降低,表明蛋白有序程度降低,与谷蛋白二级结构的测定结果相一致。Wang 等[9]试验中面筋蛋白变性温度随冻藏时间延长而降低,同时焓变有下降趋势,对比结果推测谷蛋白对冻藏期间的热变性温度的提升起到主要作用。而γ-PGA 组有效抑制了冻藏导致的变性起始温度升高,冻藏7 周后使得变性温度Tp增大至65℃,较空白组提升4 ℃,并发现其热焓值并没有随冻藏时间延长而减小,这可能是γ-PGA 有效抑制了谷蛋白网络的劣变,从而减缓了热焓值的降低。
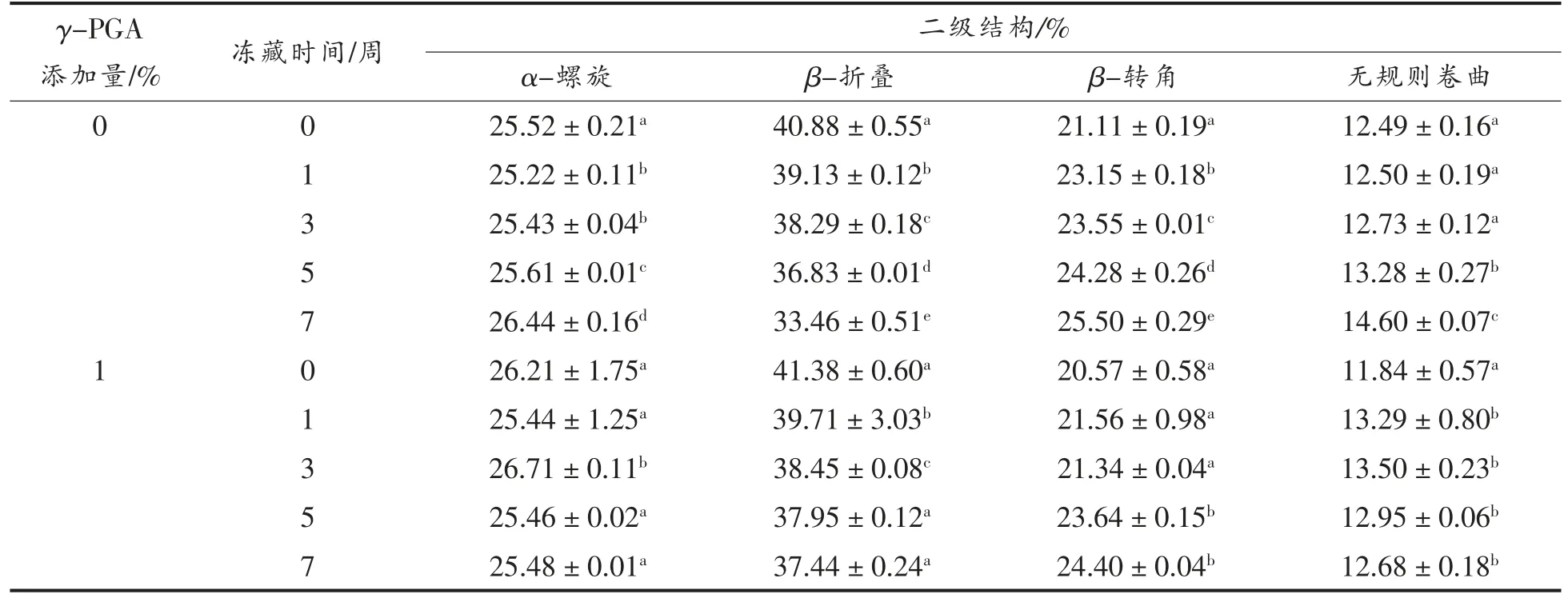
表2 γ-PGA 对谷蛋白二级结构的影响Table 2 Effect of γ-PGA on secondary structure of glutenin
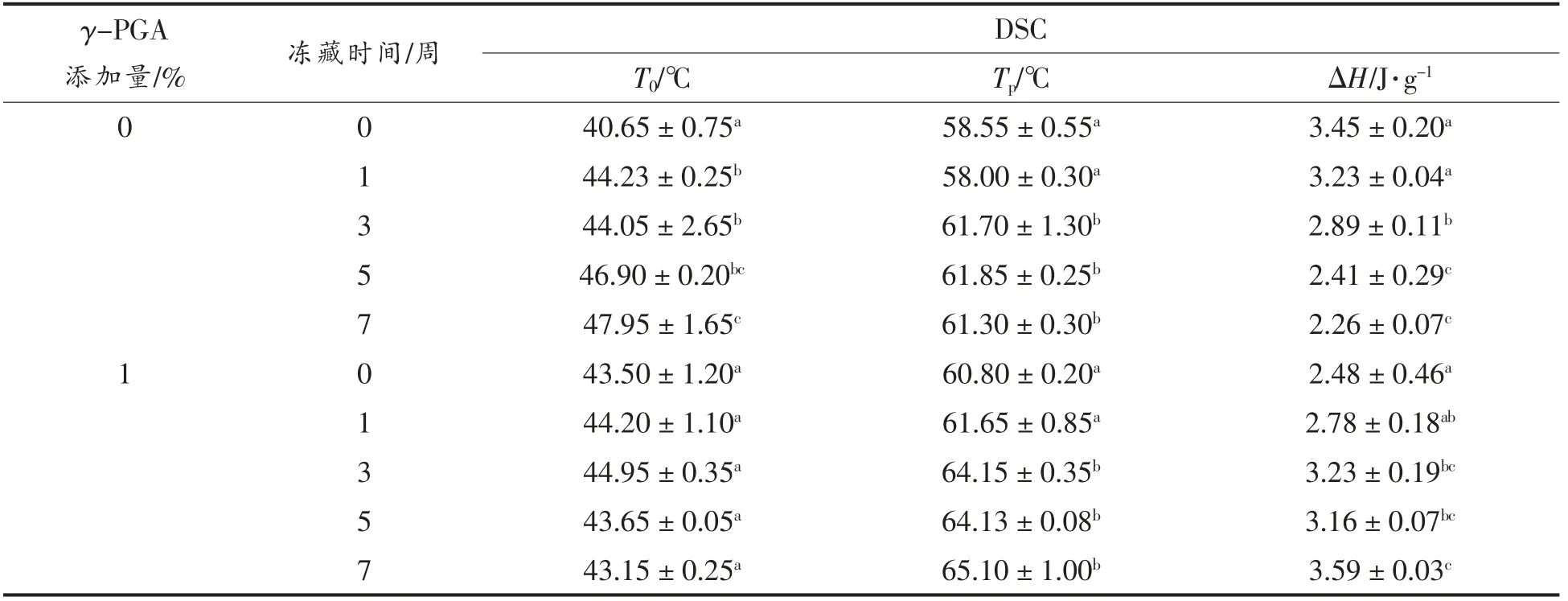
表3 γ-PGA 对谷蛋白热力学特性的影响Table 3 Effect of γ-PGA on thermodynamic properties of glutenin
2.5 γ-PGA 对谷蛋白流变学特性的影响
黏弹性被认为是评价面团品质的重要指标之一,而谷蛋白是弹性的主要贡献因素。谷蛋白流变学测量的弹性模量(G′)和黏性模量(G″)如图2所示。与面筋蛋白相一致,谷蛋白在频率0.1~100 Hz范围内其G′和G″都随着频率的增加逐渐增大,且G′始终大于G″。当冻藏时间延长,谷蛋白的弹性模量(G′)和黏性模量(G″)均出现下降的趋势,这与Xuan 等[17]研究结果保持一致。而添加了1%的γ-PGA 后,虽然冻藏初期谷蛋白的弹性模量(G′)低于空白组,但其受到冻藏影响的程度较小,冻藏7 周后其弹性模量(G′)比空白组高,表明其有效抑制了冻藏导致的谷蛋白弹性的降低,而黏性模量(G″)却与之相反,在冻藏前后其黏性性能均低于空白组,这表明γ-PGA 并没有对谷蛋白的黏性性能产生积极作用,前人研究也发现面团及面筋蛋白的弹性性能主要由谷蛋白起主要贡献[18],故试验结果表明γ-PGA 并不能通过改变谷蛋白的黏性性能来减缓冻藏期间面筋蛋白黏性性能的劣变,但可以通过对谷蛋白弹性的影响来抑制面筋蛋白弹性的减弱。
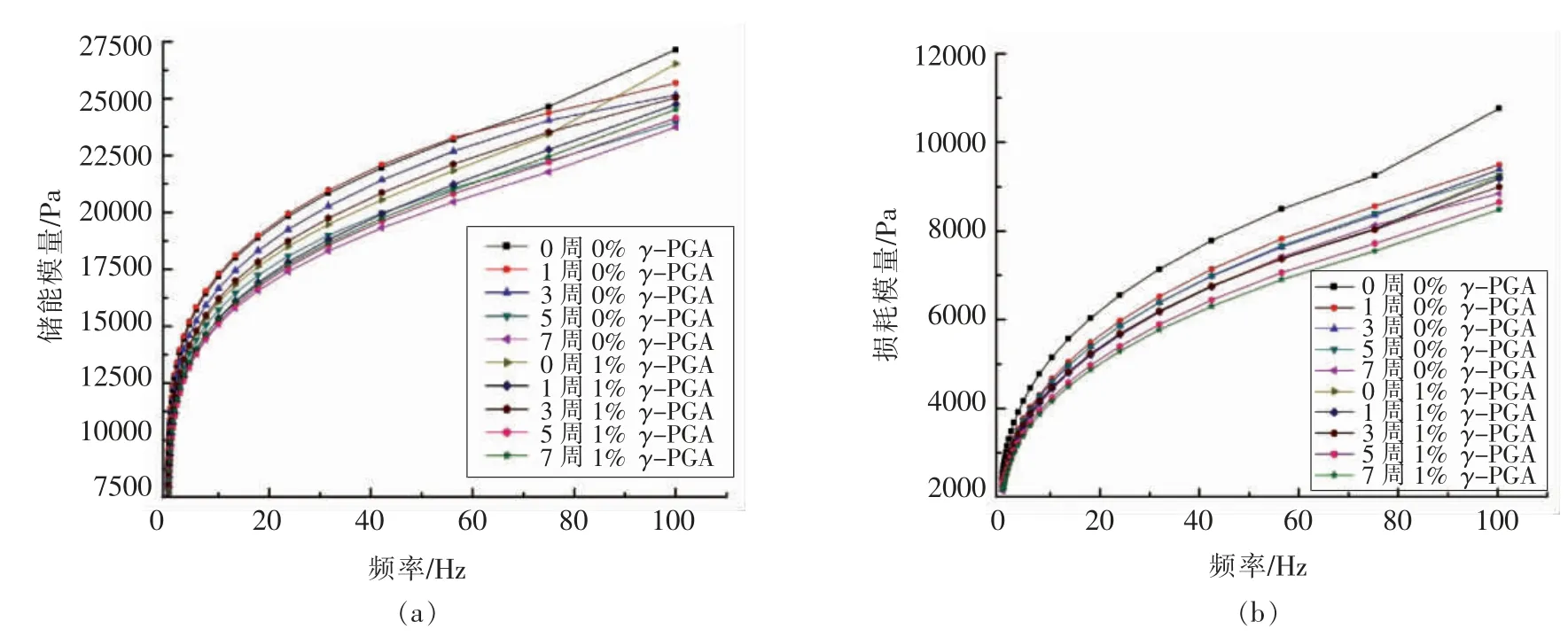
图2 γ-PGA 对谷蛋白流变学特性的影响Fig.2 Effect of γ-PGA on the rheological properties of glutenin
2.6 γ-PGA 对谷蛋白微观结构的影响
图3为不同冻藏周期谷蛋白微观结构图,冻藏初期谷蛋白通过链内和链外二硫键形成了高度网络化结构,使其具有连续的结构及较小的孔洞。随冻藏时间增加,发现其微观结构受到破坏。冻藏5 周后,谷蛋白网络趋于断裂,出现明显结块现象,主要原因可能是冻藏期间冷冻收缩作用使得相邻的小冰晶聚合形成大冰晶引发的结构断裂[19-20]。与此相比,γ-PGA 组在冻藏初期谷蛋白网络孔洞变大,但仍然呈现较好的网络结构,这是因为γ-PGA 具有良好的吸水性而与谷蛋白之间形成了竞争性吸水,导致其孔洞直径变大,但与空白组不同,随冻藏时间延长γ-PGA 组的谷蛋白网络的连续性比空白组好,且断裂得较少,γ-PGA 良好的增稠性能产生水分滞留效果,使得水分流动性降低可能是其中的主要原因[6]。较连续的网络结构也使谷蛋白的热稳定性增加,与上述试验结果相吻合。同时表明γ-PGA 有效缓解了冻藏期间谷蛋白的劣变,其效果可能在延缓面筋蛋白劣变中做出了贡献。

图3 γ-PGA 对谷蛋白微观结构的影响Fig.3 Effect of γ-PGA on the microstructure of glutenin
3 结论
研究表明添加γ-PGA 可以抑制冻藏期间谷蛋白水分的流失,促使谷蛋白中自由水向弱结合水转换,增强其水分稳定程度,并有效减缓冻藏期间自由水增大及弱结合水含量的减少。同时γ-PGA 有效延缓冻藏导致的谷蛋白热焓值降低,提升谷蛋白变性温度,表明γ-PGA 有效地保护了谷蛋白结构。并较好地抑制了冻藏引发的结构断裂和结块现象,有效地保护了蛋白网络。冻藏导致了谷蛋白弹性及黏性性能都随冻藏时间延长而降低,添加γ-PGA 减弱了其弹性性能的下降趋势,但对黏性性能并没有产生相应作用。且γ-PGA 有效改善了冻藏导致β-折叠相对含量降低,无规则卷曲相对含量增大趋势,减弱了蛋白二级结构由有序向无序结构的转化程度。
