祁连山不同年龄青海云杉径向生长对气候变化的响应*
2021-04-10于澎涛于艺鹏王彦辉万艳芳杨文娟王顺利刘贤德
王 彬 于澎涛 于艺鹏 张 雷 王彦辉 万艳芳 杨文娟 王顺利 刘贤德
1.中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林生态环境重点实验室 北京 100091; 2.中国科学院生态环境研究中心城市与区域生态国家重点实验室 北京 100085; 3.中国科学院大学 北京 100049; 4.中国林业科学研究院林业科技信息研究所 北京 100091; 5.甘肃省祁连山水源涵养林研究院 张掖 734000)
过去几十年全球经历了显著的气候变化,总体表现为上升趋势,尤其是1980年之后的几十年,可能是北半球近1 400年以来最热的一段时期 (IPCC, 2014); 然而自2000年起,全球快速增温趋势有所减缓(Fyfeetal., 2016; Xieetal., 2017)。我国的气候变化趋势与全球一致,但西北地区在增温的同时,降水也呈现出较大变化,是气候变化敏感区和生态环境脆弱区(商莎莎等, 2018)。
气候变化通过改变温度和水分的有效性对树木生长产生影响(Andreuetal., 2007),全球和区域尺度气候变暖均可能抑制树木生长或导致树木死亡(Jumpetal., 2006; Pengetal., 2011; Teetsetal., 2018; 韦景树等, 2018),尤其在半干旱地区,森林容易受到季节性和偶然性水分胁迫,而迅速增温进一步放大了水分胁迫对森林的影响,使得树木对急剧变化的气候更为敏感 (Allenetal., 2010; Liuetal., 2013; 詹思敏等, 2019),生长速率迅速下降(Adamsetal., 2017; Pengetal., 2011; Williamsetal., 2013); 增温趋缓后,快速增温的趋势减缓,伴随着降水增加,树木生长受到的干旱胁迫能够得到一定程度的缓解(Gaoetal., 2018; Wangetal., 2019a)。
树木的生理机能会随着年龄变化而改变(Carreretal., 2004),不同年龄树木在不利环境条件下的抗性不同(Wangetal., 2019b),研究发现,树木对气候干旱化的响应存在年龄差异(Prioretal., 2014; Wangetal., 2019b; 温晓示等, 2019),且在不同地区、不同树种间也不尽相同,如东北的红松(Pinuskoraiensis)小树对增温响应敏感(Wangetal., 2019b),而桉树(Eucalyptusrobusta)大树对增温响应敏感(Prioretal., 2014)。以往研究主要关注不同年龄树木生长对迅速增温的响应,然而自2000年之后全球增温趋缓,不同年龄树木生长对增温趋缓的响应是否存在差异仍不清楚,这严重限制了气候变化下森林动态的准确预测和精准化管理。采用全样地取样法研究森林内不同年龄树木生长对迅速增温和增温趋缓2种气候变化模式的响应差异,分析树木抵抗气候变化的能力与年龄的关系,对未来气候变化下森林的精准化管理和有效应对气候变化十分必要。
青海云杉(Piceacrassifolia)林是祁连山的主要森林类型,占祁连山森林总面积的75%(张立杰等, 2008),是重要的水源涵养林,对祁连山区的水分循环和水量平衡具有举足轻重的作用。本研究采用全样地取样法研究年龄对青海云杉树木生长的影响,探讨不同年龄的树木生长及其对气候变化的响应规律,以期为预测未来气候变化下的森林动态和应对气候变化的林业管理提供科学依据。
1 研究区概况
祁连山位于我国青海省东北部和甘肃省西部边境,地处蒙新荒漠和青藏、黄土两大高原的交汇地带,是石羊河、黑河、疏勒河等56条河流的源头。近几十年来祁连山地区的气温总体呈变暖趋势,尤其是1980—2000年,气温迅速增加而降水没有明显变化,形成了相对暖干的环境条件; 2000年之后,迅速增温趋势减缓伴随着降水增加,形成了相对暖湿的环境条件(Wangetal., 2019a)。
本研究以祁连山中段北坡排露沟小流域(100°17′—100°18′E,38°32′—38°33′N)为研究区,面积为2.91 km2。长期气象观测表明,该流域极端最高气温32.4 ℃,极端最低气温-29.0 ℃,年均降水量435.5 mm,年均蒸发量1 051.7 mm,平均相对湿度60%(张雷等, 2015)。青海云杉林主要分布在该区海拔2 600~3 300 m处的阴坡、半阴坡, 阳坡以草地为主,并分布有祁连圆柏(Sabinaprzewalskii)和灌丛,灌丛优势种包括金露梅(Potentillafruticosa)、鬼箭锦鸡儿(Caraganajubata)和吉拉柳(Salixgilashanica)等。土壤在该流域呈较明显的垂直带谱,自下而上主要有山地森林灰褐土、山地栗钙土、草甸土、亚高山灌丛草甸土和高山寒漠土等类型。
2 研究方法
2.1 样本采集与实验室处理
2014年夏季进行样本采集,在排露沟小流域海拔2 800和2 900 m的阴坡选择受人为干扰较小且生长良好的青海云杉林,设置2块20 m×20 m样地(表1)。对样地内所有树木进行每木检尺调查,并用内径5.15 mm的生长锥对胸径(DBH)大于5 cm且树高大于2 m的所有树木进行取样。取样位置距地面1.3 m,沿着平行于坡向和垂直于坡向钻取2个正交叉的样芯。样芯装入塑料吸管中记录编号,带回实验室处理。
在实验室先将样品晾干,用乳胶将其固定在特定的木槽中,然后依次用不同粗糙度(120、240和600目) 的砂纸打磨,并目视交叉定年。采用测量精度为 0.01 mm 的LINTAB 树轮宽度量测系统测量年轮宽度,利用COFECHA 程序对所有样本目视定年的准确性进行检测 (Holmes, 1983)。根据COFECHA 检测结果,纠正目视定年过程中出现的错误,剔除与整个数据系列不吻合的样芯,共有335个样芯用于本研究分析(表1)。
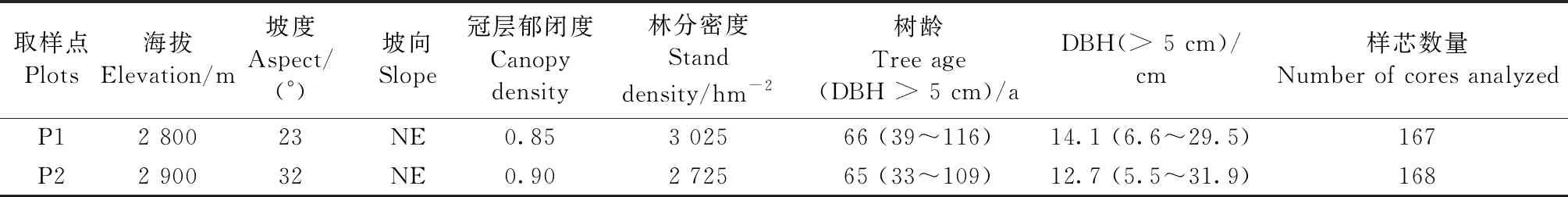
表1 祁连山中段取样点基本信息
2.2 胸高断面积年增长量(BAI)年表建立
随着树木胸径增加,年轮宽度通常呈下降趋势,从而导致年轮宽度随年龄增加而减小。将年轮宽度转化为胸高断面积年增长量(basal area increment, BAI),可以将年轮宽度随年龄增加而减小的趋势最小化,能够更准确地反映树木生长情况(Peuelasetal., 2008)。一般情况下,树木年轮学研究通过对年轮宽度系列进行去趋势处理,去除每株树由年龄造成的生长趋势,从而突出样点特征年的年轮宽度变化(吴祥定等,1996)。本研究重点关注树木生长趋势变化,因此以BAI序列反映树木生长的年际变化能更好反映树木的实际生长趋势;同时,保留气候条件对树木生长影响的信息(Jumpetal., 2006)。
BAI计算公式为:
式中:Rn为第n年树木半径;Rn-1为第n-1年树木半径(Monserudetal., 1996)。
2.3 气候数据与统计分析
气候数据(包括温度和降水)从中国气象科学数据网(http:∥data.cma.cn)获取。本研究使用距离采样点最近且海拔(2 787.4 m)相似的祁连气象站(100°9′E, 38°6′36″N)气候数据,该气象站位于采样点南50 km左右。标准化降水蒸散指数(standardized precipitation evapotranspiration index, SPEI)考虑蒸散和降水的影响,通过标准化降水与潜在蒸散的差值表征一个地区干湿状况偏离常年的程度(Vicente-serraonetal., 2010),可代表温度和降水综合影响下的干旱变化情况,SPEI越小,则越干旱。本研究基于全球SPEI数据库(http:∥spei.csic.es/index.html)中12个月尺度的SPEI数据(数据集版本为SPEIbase V2.5),并根据Wang等(2019a)研究结果,采用对研究区树木生长影响最为显著的气象要素进行树木生长影响因素分析,包括当年6月平均气温(C6-T)、上年7月到当年6月的总降水量(P7-C6-P)和当年5—8月的平均SPEI(C5-C8-SPEI)。
为了探讨1960年以来不同年龄树木生长对气候变化的响应差异,按30年划分树龄组(Qietal., 2015),将样地内所有树龄(DBH > 5 cm)为30~120年生的树木划分成3个树龄组: 小龄树(30~60年生)、中龄树(60~90年生)和大龄树(90~120年生)。本研究样地内小龄树、中龄树和大龄树的比例分别为22.52%、66.67%和10.81%,大部分青海云杉尚属于中龄林(刘兴聪, 1992)。计算每个树龄组所有树木的BAI均值,并采用最小二乘法线性回归方程拟合不同时段的BAI变化趋势,拟合线斜率为正值表示BAI呈增加趋势, 斜率为负值表示BAI呈减小趋势,斜率的绝对值越大,则树木逐年生长量增加或减少的越多。
按不同时段(1960—1980、1980—2000和2000—2013年)分析关键气象要素与单株树木BAI年表之间的Pearson相关系数,并计算不同树龄组呈显著相关的树木株数占该树龄组树木总株数的百分比,显著相关的株数占比越高,说明该树龄组树木生长受气象要素的影响越大。另外,通过对气象要素与BAI的拟合,计算不同树龄组树木对气象要素的敏感性,用BAI与气象要素的最佳拟合线性函数斜率表示,斜率绝对值越大则越敏感。
3 结果与分析
3.1 不同时段各树龄组BAI的差异
图1表明:迅速增温之前(1960—1980年),BAI呈增加趋势,随树龄增长,BAI拟合线斜率逐渐减小,小龄树BAI最大(9.83 mm2·a-1,P< 0.01),其次为中龄树(8.90 mm2·a-1,P< 0.01)和大龄树(7.07 mm2·a-1,P= 0.017);迅速增温期(1980—2000年),BAI拟合线斜率为负值,随树龄增大BAI拟合线斜率逐渐减小,小龄树、中龄树和大龄树BAI拟合线斜率分别为-5.91(P< 0.01)、-9.48(P< 0.01)和-14.97 mm2·a-1(P< 0.01);增温趋缓期(2000—2013年),BAI呈轻微增加趋势,中龄树和小龄树BAI拟合线斜率分别为2.27和2.45 mm2·a-1,大龄树BAI增加速率最小,拟合线斜率为1.02 mm2·a-1。
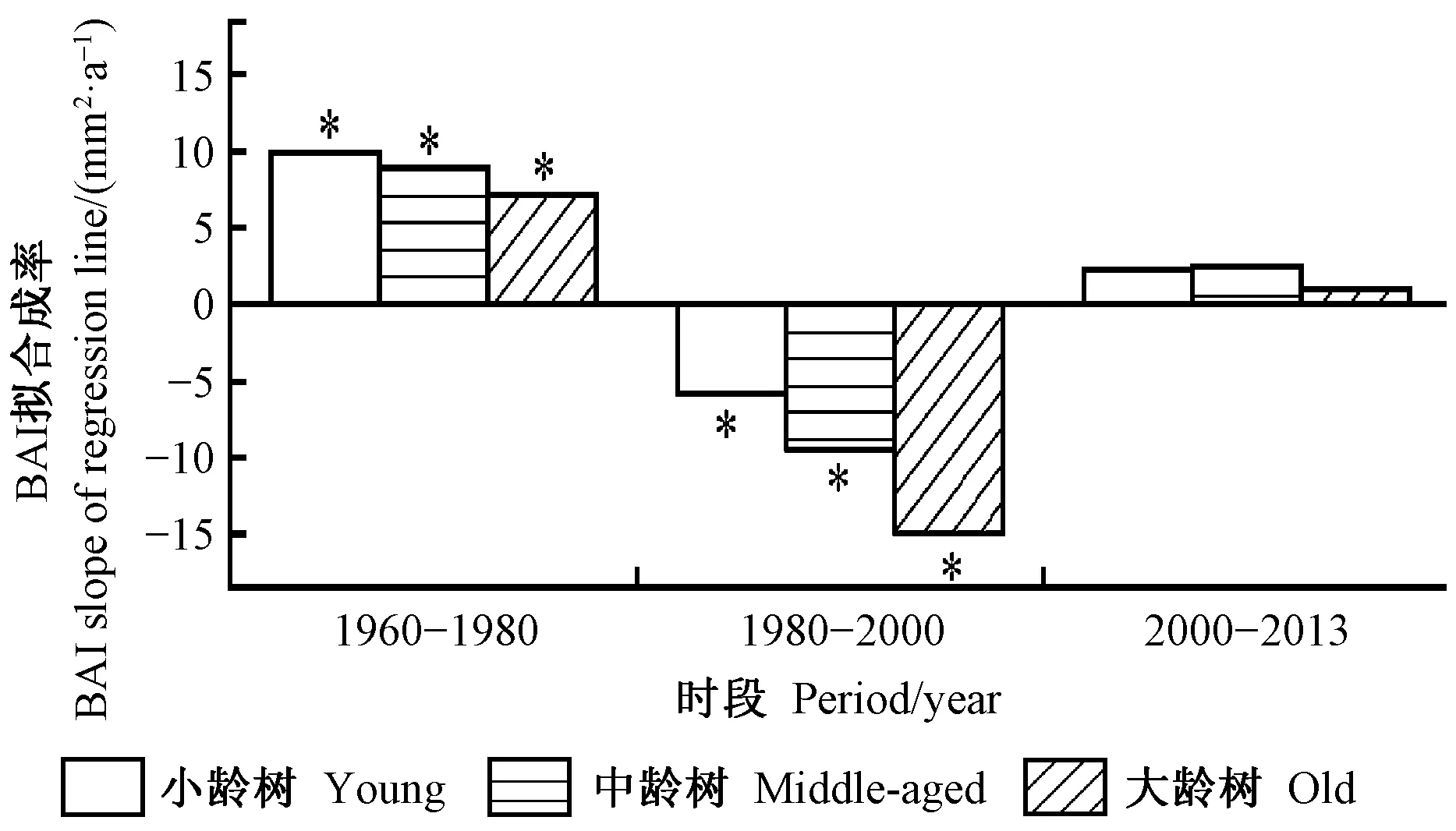
图1 不同时段不同树龄组青海云杉BAI拟合线斜率(*: P<0.05)
3.2 BAI与气象因子的相关性
迅速增温之前、迅速增温期和增温趋缓期,各树龄组BAI与C6-T主要呈负相关(图2),且呈负相关的大龄树株数百分比高于小龄树和中龄树。迅速增温期,BAI与C6-T呈显著负相关的株数百分比随树龄增增长逐渐增加,小龄树BAI与C6-T呈显著负相关的株数百分比为56.19%,中龄树为59.32%,大龄树为65.00%。增温趋缓期,各BAI与C6-T呈显著负相关的株数百分比相比迅速增温期均明显下降,但呈显著负相关的株数百分比仍随树龄增长而增加,其中小龄树呈显著负相关的株数百分比为2.86%,中龄树和大龄树分别为3.76%和10.00%。
迅速增温之前、迅速增温期和增温趋缓期,各树龄组BAI与P7-C6-P主要呈正相关(图2)。迅速增温期,BAI与P7-C6-P呈显著正相关的株数百分比随树龄增长逐渐降低,小龄树BAI与P7-C6-P呈显著正相关的株数百分比为10.48%,中龄树为6.57%,大龄树BAI与P7-C6-P不存在显著相关关系。增温趋缓期,各BAI与P7-C6-P呈显著正相关的株数百分比较迅速增温期均明显增加,其中,小龄树BAI与P7-C6-P呈显著正相关的株数百分比最高(53.33%),其次为中龄树(36.62%)和大龄树(25.00%)。
迅速增温之前、迅速增温期和增温趋缓期,各树龄组BAI与C5-C8-SPEI主要呈正相关(图2)。迅速增温期,BAI与C5-C8-SPEI呈显著正相关的株数百分比随年龄增加逐渐增加,小龄树BAI与C5-C8-SPEI呈显著正相关的株数百分比为62.85%,中龄树为68.54%,大龄树为75.00%。增温趋缓期,SPEI增大,干旱程度得到缓解,BAI与C5-C8-SPEI呈显著正相关的株数百分比随年龄增加而降低,小龄树为88.57%,中龄树为84.98%,大龄树为70.00%。
3.3 BAI对气象因子的敏感性
迅速增温期,各树龄组BAI对C6-T的敏感性均大于迅速增温之前,大龄树对迅速增温最为敏感,(-84.1 mm2·℃-1),中龄树的敏感性次之(-56.2 mm2·℃-1),小龄树的敏感性最弱(-43.7 mm2·℃-1),仅为大龄树的1/2; 增温趋缓期,BAI对C6-T的敏感性明显减小,但C6-T对小龄树(2.9 mm2·℃-1)和中龄树(1.1 mm2·℃-1)的树木生长具有较弱的促进作用,对大龄树(-13.2 mm2℃-1)仍呈轻微的抑制作用(图3)。
迅速增温期,各树龄组BAI对P7-C6-P的敏感性均小于迅速增温之前,敏感系数为4.4~5.9 mm2·mm-1,尽管各树龄组的敏感性差异较小,但随着树龄增长敏感性逐渐增加,即大龄树>中龄树>小龄树; 增温趋缓期,各树龄组BAI对P7-C6-P的敏感性相比迅速增温期略有增加,但仍是大龄树的敏感性最大(图3)。
迅速增温期,各树龄组BAI对C5-C8-SPEI的敏感性总体比迅速增温之前略有下降,其中,大龄树的敏感性为603.8 mm2·SPEI-1,中龄树为414.4 mm2·SPEI-1,小龄树为317.6 mm2·SPEI-1,仅为大龄树的1/2;增温趋缓期,小龄树的敏感性略有上升(355.7 mm2·SPEI-1),而中龄树和大龄树的敏感性均有所下降(图3)。
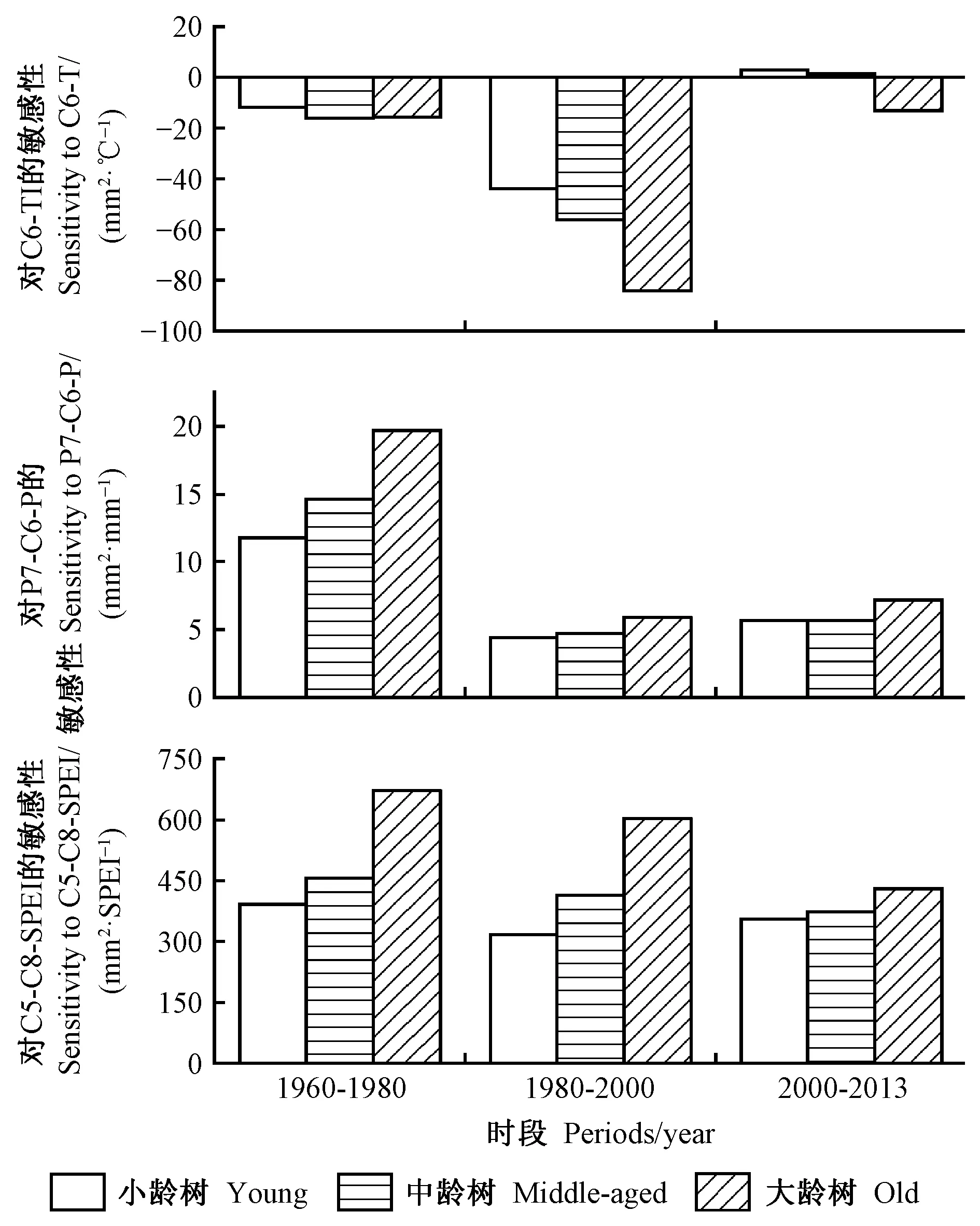
图3 不同树龄组BAI对气象因子的敏感性
4 讨论
4.1 不同时段各树龄组BAI差异的主要影响要素
基于树木生长理论,可将树木生长(本研究主要指BAI)分为3个时期:指数增长期、线性增长期和衰减期 (Colbertetal., 2004),健康树木BAI在指数增长期或线性增长期应呈增加或平缓增加趋势(Leblancetal., 1990; Dittmaretal., 2003)。青海云杉一般到120~140年树龄进入成熟阶段(刘兴聪,1992),树木年生长量才会减缓。本研究区青海云杉以中、幼龄林为主,所有树木的树龄至2013年均小于120年,同时所选样地近几十年没有明显的森林结构变化和人类活动干扰,因此, 1980—2000年各树龄组BAI均呈显著下降趋势可归因于外界环境条件的变化,尤其是1980年之后迅速增温所致。
近几十年来,迅速增温导致的干旱胁迫限制了红松、欧洲水青冈(Fagussylvatica)、挪威云杉(Piceaabies)和青海云杉等树木生长(Wangetal., 2019a; Jumpetal., 2006; Sidoretal., 2015; Liangetal., 2016; Zhangetal., 2018),其中青海云杉对水分条件尤为敏感(Liangetal., 2010),其生长和发育受可利用水分的限制。本研究中,迅速增温期间,各树龄组大多数青海云杉BAI与温度呈显著负相关,表明温度增加主导BAI的下降趋势。增温可导致潜在蒸散增加,树木生长需水量增大,在可利用水不增加的情况下,树木需水满足程度降低,受到的干旱胁迫加剧,从而导致生长减缓; 同时,该时段内SPEI下降(SPEI越小,表明干旱越严重),且大多数树木生长与SPEI呈显著正相关,进一步证明树木受到更强烈的干旱胁迫。
然而2000年之后,全球快速增温趋势有所减缓,进入变暖间歇期(Fyfeetal., 2016; Xieetal., 2017)。在本研究区,增温趋势减缓的同时伴随着降水增加(Wangetal., 2019a),形成了较1980—2000年湿润的气候条件,干旱胁迫得到了一定程度缓解(Gaoetal., 2018)。另外,增温可增加生长季长度,较长的生长季和较湿润的气候条件有利于树木生长,进而使得各树龄组BAI在2000年之后均呈增加趋势。Gao等(2018)的研究也表明2000年之后青海云杉BAI恢复增加趋势。
4.2 同一时段各树龄组BAI差异的主要影响要素
本研究发现,1980—2000年迅速增温期,大龄青海云杉的生长速率下降更严重。美国弗吉尼亚州中南部的白栎(Quercusfabri)和意大利东部阿尔卑斯山高海拔地区的欧洲山毛榉(Fagussylvatica)均为老树,对气象要素也具有较高敏感性(Carreretal., 2004; Copenheaveretal., 2011),与本研究结果基本一致。半干旱地区森林容易受到季节性和偶然性水分胁迫的影响,甚至对略有增加的水分亏缺响应敏感(Liuetal., 2013), 1980—2000年迅速增温和降水减少共同形成了暖干化的环境条件,水分亏缺抑制了树木生长(段洪浪等, 2015; Cabonetal., 2018; Zhangetal., 2018); 在此期间,大龄树BAI与SPEI呈显著相关的株数百分比和敏感性大于中龄树和小龄树,因此大龄树受到的干旱限制作用最大,生长下降趋势最大。另外,不同年龄树木的根冠比存在差异,对逆境的适应能力也不同,根冠比越大,抗旱能力越强。有研究发现,随着年龄增加,根冠比先减小后趋于稳定(Perietal., 2006),因此,年龄较大的青海云杉抗逆性较差,抗旱能力小于小龄树和中龄树。
2000年之后增温趋缓并伴随着降水增加形成了相对湿润的环境,各树龄组BAI均转为增加趋势,小龄树和中龄树BAI增加趋势略高于大龄树。小龄树和中龄树BAI与SPEI呈显著相关的株数百分比大于大龄树,表明增温趋缓后小龄树和中龄树受到的干旱胁迫缓解程度大于大龄树。这主要是因为大龄树处于林冠上层,受到更强的光照和较大的风速影响,而中龄树和小龄树树高一般较大龄树小,处于大龄树下面,受到的光照较大龄树少,因此大龄树需水量高于中龄树和小龄树(Zhouetal., 2013),尽管该时段干旱在一定程度上得到了缓解,但仍不能满足大龄树的需水量,对大龄树来说需水量缺口较中龄树和小龄树大,因此大龄树的生长速率在增温趋缓后恢复程度小于小龄树和中龄树。
另外,与年龄相关的树木生长对气候的响应差异与树木次生生长的时间变化有关。有研究认为,春季小龄树形成层细胞开始分裂的时间比大龄树早(Lietal., 2013),且生长素产生于活化的芽中并沿茎向基部输出(Ugglaetal., 1998),因此与大龄树相比,小龄树的茎基部更接近树冠,也更接近生长素的来源(Rossietal., 2008)。基于上述2点,可推断年龄较小的树木具有更长的生长季,产生的细胞数量比大龄树多,故迅速增温时小龄树生长速率下降较大龄树少。而且,根系结构也会影响树木生长,大龄树可在更深的土壤层中有更发达的根系(Børjaetal., 2008),从而获得更多的水分和养分,然而本研究样地位于山坡上,土壤较薄(王彬等,2016),且青海云杉为浅根系树种,主根系通常分布在0~60 cm土层,这导致大龄树在水分吸收方面并不比小龄树和中龄树更有优势。
5 结论
本研究采用全样地取样法研究迅速增温期和增温趋缓期不同年龄的青海云杉树木生长及其对气候变化的响应规律。得到以下结论: 1) 1980—2000年迅速增温期,水分亏缺导致所有年龄的青海云杉BAI均呈减小趋势,但大龄树更敏感,减小趋势比小龄树和中龄树更严重; 2) 2000—2013年增温趋缓期,形成了相对湿润的环境,所有年龄青海云杉受到的干旱胁迫均减缓,其中小龄树和中龄树的缓解程度大于大龄树,该时段小龄树和中龄树BAI拟合线斜率大于大龄树; 3) 青海云杉大龄树对干旱更敏感且抗旱性差,迅速增温导致的干旱胁迫对大龄树影响最大,而增温趋缓后小龄树和中龄树恢复生长的能力更强。因此,在未来气候变化下的森林管理中,应考虑树木生长对气候响应的年龄差异。
