栓皮栎叶片和枝条非结构性碳水化合物调配关系研究
2021-04-07魏龙鑫章异平李艺杰张玉茹
魏龙鑫,章异平,李艺杰,张玉茹
(河南科技大学林学院,河南 洛阳 471023)
植物通过光合作用产生的碳水化合物,按存在形式分为结构性碳水化合物(structural carbohydrates,SC)和非结构性碳水化合物(non-structural carbohydrates,NSC)[1]。其中后者的主要组分为可溶性糖和淀粉,可在植物碳库水平较低时起缓冲调节作用[2],能很好地表征树木整体的碳供应状况(不足或过剩)[3]。树木中NSC含量的大小是碳同化和碳消耗(即供应和需求)平衡关系的结果[4-5]。当碳同化速率低于碳消耗速率时,NSC含量下降,反之,NSC含量升高。Myers等[6]研究发现,NSC的储存有助于提高树木生存能力以及对外界干扰的缓冲能力。因此,掌握树体内不同器官间的NSC调配特征对于理解树木生长规律,预测生长趋势具有重要意义。
叶片是植物主要的光合作用器官,是重要的碳源,对树木生理状况及碳供应敏感,是较好的指示器官[7],也是目前树木NSC含量研究中最常采用的观测器官[8-9]。而枝条是植物碳供应器官(叶片)和碳需求器官(茎干、根等)的通道,起着运输营养物质的作用[10-11]。有研究指出,对于一些落叶树种,枝条中存储的NSC可以为树体发芽展叶提供能量[12-13]。对于一些径向生长早于展叶的树种,枝条存储的NSC可能还是茎干径向生长启动的主要能量来源[14]。在生长过程中,树体叶片和枝条间的NSC协调分配,使得叶片和枝条间的NSC含量很可能存在动态关联[15]。因此,研究两器官间NSC及其组分含量的动态变化对于了解树体内碳分配过程具有重要意义。
目前,已有研究多针对树体单个器官NSC含量开展[16-18],鲜见叶片和枝条间以及不同器官间NSC含量调配的相关分析报道。部分研究虽采集了叶片和枝条的同期NSC含量数据,但由于采样次数较少,无法进行器官间NSC含量的动态变化分析。此外,采样时间常集中于生长旺盛季,而忽略了非生长季和生长季初期(萌芽期)的重要变化。本研究拟以暖温带主要造林树种栓皮栎(Quercusvariabilis)为研究对象,在完整的生长季,通过半月-月尺度周期性采样,分析叶片和枝条NSC及其组分含量的动态变化特征,掌握NSC在叶片和枝条间的分配规律,加深对栓皮栎碳调配机制的理解,为进一步评估栓皮栎群落的固碳潜力提供基础数据。
1 材料与方法
1.1 研究区概况
研究样地设置在河南省洛宁县西南部的全宝山国有林场,位于豫西山区秦岭东段熊耳山北坡(111.38°~113.54°E,34.10°~34.18°N)。研究区年均气温10.5 ℃,年降水量863.6 mm,年蒸发量1 562.8 mm,全年无霜期186 d。主要植被类型有壳斗科(Fagaceae)、胡桃科(Juglandaceae)、禾本科(Gramineae)等。栓皮栎(Quercusvariabilis)在低海拔(<1 000 m)范围内常形成纯林[19]。
在栓皮栎为优势种分布的海拔下限(650 m)设置样地,样地坡度25°,林分郁闭度0.9。样树平均树龄26 a,树高16 m,胸径16 cm。从自动气象站监测数据可知,在观测采样期内(2016年5月—2017年6月),样地空气温度年均值为13.4 ℃,土壤温度年均值为13.4 ℃,土壤含水量年均值为12.6%。林下灌木层以栓皮栎幼树、连翘(Forsythiasuspensa)为主,草本层以羊胡子草属(Eriophorum)为主。
1.2 叶片和枝条样品的采集及预处理
采样频率根据栓皮栎径向生长期确定[14]。生长前期每15~30天采样1次(2—6月),生长后期或非生长季每1~3个月采样1次(7月至次年2月),共采样13次(2016年6月由于道路原因未采样)。样地规格大小为10 m×10 m,在样地中,选取直立健康、非孤立、大小(年龄、树高、胸径)基本一致的栓皮栎优势木3株。使用高枝剪,剪取样树上部南向未遮阴、生长良好的1、2年生枝条,并筛选摘取当年生健康叶片。叶片和枝条的采集量以烘干粉碎后达到25 g以上为准,枝条较细的未去皮。将叶片和枝条样品分别统一标号后立即放入冷藏箱中保存。当天带回实验室放入烘箱杀青2 h,样品75 ℃烘干至质量恒定,将烘干样品粉碎过筛(孔径 <1 mm)。
1.3 叶片物候观测
2016年5月至2017年6月,对栓皮栎进行叶片物候观测。另外选取健康生长的栓皮栎优势木3株,做好标记并进行周期性观测,观测时间为2月下旬至5月下旬,观测频率为每周1次。通过测量叶片长度得到叶片生长百分比,并在此基础上,利用Gompertz生长模型[20-21]拟合叶片生长曲线,得到叶片物候主要阶段的具体日期。主要包括以下几个阶段:4月上旬展叶初期(展叶>5%),4月中下旬展叶中期(展叶>50%),5月上旬展叶末期(展叶>95%)[22]。
1.4 非结构性碳水化合物的测定
称取叶片和枝条样品粉末各(0.1±0.000 1 g),采用蒽酮浓硫酸法测定叶片和枝条中可溶性糖(soluble sugar,SS)和淀粉(starch,S)的吸光值[23-24]。根据葡萄糖标准曲线换算为相应的百分比含量(质量分数),可溶性糖与淀粉的含量之和即为非结构性碳水化合物的含量。单个采样日单株样树测定3个重复。
1.5 数据处理
采用多因素方差分析法,分析器官、日期及其交互作用对NSC及其组分含量影响的显著性;利用配对样本t检验,对比各采样日期叶片和枝条NSC组分含量及其差异显著性(显著水平α=0.05);利用Pearson相关系数,分析两器官间NSC组分含量及其相关性。以上分析均采用SPSS 20.0完成,图表制作利用Origin 9.0完成。
2 结果与分析
2.1 栓皮栎叶片和枝条中可溶性糖(SS)含量的时间动态变化
栓皮栎叶片和枝条中可溶性糖(SS)含量的影响因素分析结果见表1。
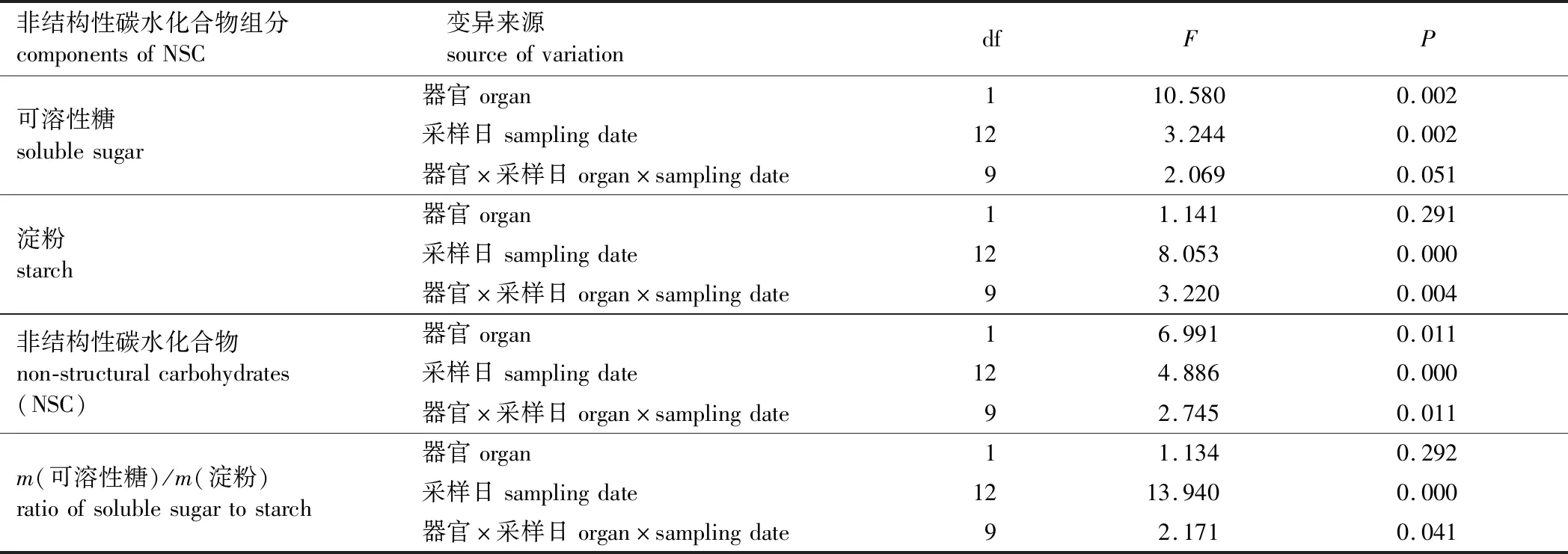
表1 栓皮栎非结构性碳水化合物及其组分含量影响因子方差分析Table 1 Analyses of variance of influence factors on the contents of non-structural carbohydrates (NSC) and their components
由表1可以看出,栓皮栎叶片和枝条的可溶性糖含量均随时间变化差异显著(df = 12,F= 3.244,P<0.05)。从时间动态可以看出(图1),叶片可溶性糖含量(质量分数)高于枝条,二者的均值分别为5.71%和4.78%。叶片可溶性糖含量最大值出现在4月下旬(7.68%),最小值出现在6月中旬(3.63%)。与叶片不同,枝条可溶性糖含量从10月开始上升,并于2017年2月中旬达到年内最大值(6.96%),之后总体上呈下降趋势,并于6月中旬达到年内最小值(2.35%)(图1)。

*表示同一采样日叶片和枝条间差异性显著。下同。* indicated the significant differences between leaves and branches on the same sampling date.The same below.图1 栓皮栎叶片和枝条中可溶性糖(SS)含量随时间的动态变化Fig.1 Seasonal dynamics of soluble sugar content inleaves and branches of Q.variabilis
2.2 栓皮栎叶片和枝条中淀粉(S)含量(质量分数)的时间动态变化

图2 栓皮栎叶片和枝条中淀粉(S)含量随时间的动态变化Fig.2 Seasonal dynamics of starch content in leaves and branches of Q.variabilis
与可溶性糖相似,栓皮栎叶片的淀粉含量(质量分数)(3.23%)略高于枝条的淀粉含量(3.11%),特别是在生长旺盛期(5—8月,图2)。叶片和枝条的淀粉含量均随时间变化呈显著性差异(df = 12,F= 8.053,P<0.05)。叶片淀粉含量在生长季初期(3月—4月下旬)呈上升趋势,从5月开始呈下降趋势。枝条淀粉含量在3月到6月下降趋势明显,并在6月中旬达到年内最小值,仅为0.82%。生长季末(10月)枝条淀粉含量上升,达到年内最大值(5.38%)。
2.3 栓皮栎叶片和枝条中非结构性碳水化合物(NSC)含量(质量分数)的时间动态变化
栓皮栎叶片NSC含量(质量分数)均值(8.94%)高于枝条NSC均值(7.90%),但在展叶初期(2017年3—4月),叶片NSC含量低于枝条的(图3)。

图3 栓皮栎叶片和枝条中非结构性碳水化合物(NSC)含量随时间的动态变化Fig.3 Seasonal dynamics of non-structural carbohydrates (NSC) content in leaves and branches of Q.variabilis
叶片NSC含量(质量分数)随时间变化差异显著(df = 12,F= 4.886,P<0.01),最大值(12.29%)出现在5月,最小值(5.46%)出现在6月。其变化趋势与叶片可溶性糖(SS)、淀粉(S)含量高度正相关(叶片NSC与SS:R= 0.839,P<0.01。叶片NSC与S:R= 0.883,P<0.01,表2)。枝条NSC含量随时间变化亦存在显著性差异(df = 12,F= 4.886,P<0.01),最大值(10.96%)出现在3月底,最小值(3.17%)出现在6月中旬。同叶片类似,枝条NSC含量与枝条可溶性糖、淀粉均显著正相关(枝条NSC与SS:R= 0.830,P<0.01。枝条NSC与S:R= 0.895,P<0.01)。
2.4 栓皮栎叶片和枝条中可溶性糖与淀粉比值的时间动态变化
栓皮栎叶片、枝条的可溶性糖与淀粉比值(SS/S)均随时间变化显著(df = 12,F= 13.940,P<0.01),两者的波动范围分别为1~7和1~5(图4)。叶片和枝条SS/S的最大值分别为6.89和4.83,出现时间相同(2017年5月中旬)。二者的SS/S变化趋势均与其淀粉含量呈显著负相关(叶片SS/S与S:R=-0.789,P<0.01。枝条SS/S与S:R=-0.764,P<0.05,表2)。相比而言,叶片SS/S的波动较枝条更剧烈。

图4 栓皮栎叶片和枝条中可溶性糖与淀粉的质量比值(SS/S)随时间的动态变化Fig.4 Seasonal dynamics of the mass ratio of soluble sugar to starch (SS/S) mass in leaves and branches of Q.variabilis
2.5 栓皮栎叶片和枝条间NSC及其组分含量的相关性
在整个观测期,利用Pearson相关性分析可知,叶片和枝条两器官间的NSC及其组分含量的相关性均未达到显著水平,但两器官的SS/S的值存在显著正相关(R= 0.713,P<0.05,表2)。但2017年生长前期(3—6月),叶片和枝条NSC含量变化趋势基本一致,存在叶片NSC含量变化滞后于枝条15~20 d的特征(R= 0.986,P<0.001)。
利用配对样本t检验,对比各采样日期两个器官间NSC及其组分含量的差异显著性。结果表明,两器官(叶片和枝条)的可溶性糖和NSC含量的显著性差异主要出现在生长季前期和后期(4—6月、10月,图1和图3),均为叶片含量高于枝条。而两器官的淀粉含量差异相对复杂,在生长旺盛季(5—8月),叶片淀粉含量显著高于枝条,但在生长初期(3—4月)和末期(11月),枝条淀粉含量显著高于叶片(图2)。相比而言,两器官间SS/S的值的显著性差异只出现在6月中旬(图4),叶片SS/S值高于枝条的。

表2 栓皮栎叶片与枝条间非结构性碳水化合物及其组分含量的相关性分析Table 2 Correlation analysis on the content of non-structural carbohydrates (NSC) and their components in leaves and branches of Q.variabilis
3 讨 论
3.1 栓皮栎叶片和枝条NSC含量的基本特征
本次研究结果显示,栓皮栎叶片和枝条的NSC含量波动范围分别为5%~13%和3%~11%,均值分别为8.94%和7.90%。相关研究发现,中国不同森林类型植物叶片的NSC含量呈现由北向南依次降低的趋势,其中温带阔叶林、亚热带常绿阔叶林和热带雨林分别为10.03%、9.12%和8.01%[17]。而笔者研究区位于温带和亚热带的过渡区域(北亚热带),栓皮栎叶片NSC含量与亚热带植物相接近,这说明本研究结果符合植物叶片NSC含量的区域地带性特征,与其他研究具有可比性。而枝条作为树体由叶片向茎干(或根部)的养分输送通道,通常认为枝条NSC含量的波动范围较小[25]。但也有研究指出,欧洲温带森林的3种常见树种欧洲鹅耳枥(Carpinusbetulus),欧洲水青冈(Fagussylvatica)和无梗花栎(Quercuspetraea)其枝条NSC组分含量随时间变化差异显著(P<0.01),枝条淀粉含量的波动范围为1.3%~11.9%[12]。张海燕[2]研究也发现中国温带树种老枝NSC含量变化范围在4.1%~15.0%。栓皮栎叶片NSC及其各组分含量均值均高于枝条,这与Martínez-Vilalta等[26]利用Meta分析全球121个研究案例得到的研究结果是一致的。因为叶片是树木的“碳源”器官,通过光合作用实现碳同化,为树木生长提供碳水化合物,叶片较高的代谢速率需要其保持正常的细胞膨压,因此NSC含量特别是可溶性糖含量相对较高。而枝条是“碳汇”器官,是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者,起着树体内传输碳水化合物的作用[10-11]。与茎干/根部相比,枝条在生长季并不储存大量的NSC[27]。
栓皮栎叶片和枝条NSC组成均以可溶性糖为主。可溶性糖和淀粉在植物生长过程中发挥着不同的作用,一般认为,可溶性糖可快速为植物供能[28],而淀粉是植物的长期贮能物质[1]。两者组成比例的高低,除受树种特性(常绿、落叶)等内在因素的影响外[8,15],温度[29]、水分[30]、海拔[10,31]等外在因素对其组成比例都有不同程度的影响。通常认为,在寒冷地区,为适应低温环境,植物会合成更多的可溶性糖来应对极端环境[3];在湿润地区,植物不需要通过增加可溶性糖类的积累来增强渗透压以获取更多的水分,因此较干旱区域,植物体内淀粉含量会相对更高。本次研究区地处北亚热带山地区域,年均气温7.7 ℃,土壤含水量12.6%。在这种暖干生境下,栓皮栎树体内可溶性糖含量比例更高或许是其对环境适应性的一种综合表现。
3.2 栓皮栎生长中叶片和枝条NSC含量的动态协调变化
栓皮栎叶片和枝条NSC含量随时间的变化趋势并不一致,且极值大小及出现时间亦不相同。叶片和枝条两器官间NSC含量的显著性差异主要出现在生长季前期和后期。因为不同生长阶段,叶片和枝条的功能性差异导致不同器官在碳同化和碳消耗上并不同步。作为落叶树种,栓皮栎需要储存较多NSC为来年展叶提供能量。Klein等[12]研究也证实,枝条中的C存储对叶片重建具有重要作用,新叶生长所需要的C几乎全部来自于枝条淀粉的分解。因此4月中旬,展叶盛期,枝条NSC被显著消耗为叶片生长供应能量,同期叶片通过光合作用增加碳同化产物,叶片NSC含量开始升高。5月,随着展叶完成,叶片碳同化速率趋于稳定。但随着树木生长速率的加快,叶片新生成的碳更多地被输送到树木碳需求器官(茎干、根)。6月,栓皮栎茎干进入径向生长快速阶段[14],同期叶片和枝条NSC含量同时达到年内最小值。在生长季末,在满足茎干、根等器官的能量需求后,栓皮栎枝条开始积累NSC,这主要是为来年发芽展叶、茎干径向生长做准备。因为来年春季,落叶树种栓皮栎需要碳水化合物来重建冠层。此外,作为环孔材,栓皮栎茎干径向生长早于叶片生长[14],需要大量的碳水化合物为茎干形成层生长启动提供能量。这种不同器官间的碳供需协同变化,有利于栓皮栎生长,增强其应对极端环境(如干旱)的抵抗力和适应性。
综上可知,栓皮栎叶片和枝条两器官间的碳存储存在互相平衡、互相调配的关系。但这种关系并非简单的线性相关,而是表现出NSC含量变化叶片滞后于枝条15~20 d的特征。因此,今后在使用模型估算栓皮栎群落固碳潜力时,需综合考虑碳分配在不同器官间的非线性关系、碳源-碳汇器官间NSC含量变化的时滞性等因素。
