近50年中国典型木本植物展叶始期温度敏感度变化及原因
2019-12-18徐韵佳葛全胜戴君虎王焕炯
徐韵佳,葛全胜,戴君虎,王焕炯,*
1 中国科学院地理科学与资源研究所, 陆地表层格局与模拟重点实验室, 北京 100101 2 中国科学院大学, 北京 100049
近几十年来,北半球温带地区大部分植物春季物候期明显提前,而秋季物候期的变化空间差异较大[1-3]。与其他气候因素(日照、降水量等)相比,温度升高是造成木本植物春季物候期提前的最主要因素[4-5]。因此,春季物候期对温度变化的响应程度受到广泛关注。温度每升高1℃,植物物候期变化的天数定义为温度敏感度[6-7]。研究表明,春季物候温度敏感度高的物种,在群落中的覆盖度和生物量占据优势[8]。因此,研究物候的温度敏感度变化有助于评估不同植物对气候变化的响应与适应能力,从而掌握未来生态系统中的种群动态[8-9]。
很多学者研究了植物春季物候温度敏感度在不同时期的变化特征。例如,在瑞士地区的研究表明,植物春季物候的温度敏感度在过去300年总体呈升高态势,而在1890—1950和1970—2007年两个典型暖期出现了降低趋势[10]。在中国,Dai等[11]分析了牡丹江地区40个木本植物1978—2014年春季展叶始期温度敏感度的变化特征,发现75%的植物温度敏感度呈上升趋势,25%呈降低趋势。在欧洲地区,Fu等[6]分析了1980—1994年和1999—2013年两个时段7种广泛分布的木本植物展叶始期对温度的敏感度变化,发现后时段的平均温度敏感度相较于前时段下降了40%。但当比较30年变化时,两时段(1951—1980年和1984—2013年)温度敏感度不存在显著差异[12]。
造成上述温度敏感度时间变化的原因仍有争议。一些学者认为植物春季物候期温度敏感度变化与植物在冬季受到的冷激量有关[6,13]。当冬季温度升高时,植物所经历的冷激量降低,从而使萌芽所需的积温量增加,降低春季物候期的温度敏感度[14-16]。也有学者认为光周期与植物春季物候期温度敏感度有关。一些树种对日照长度较为敏感,当展叶始期提前到一定程度,达到了植物开始发育昼长的临界值,展叶始期将不再提前[17]。另外,有研究表明春季气温变率与植物展叶始期的敏感度有关[18-20]。春季气温变率较大的地区更易发生霜冻害事件[21],植物通过降低温度敏感度这一适应策略来避免遭受霜冻灾害[22-23]。
国内对温度敏感度的研究主要集中在个别站点的种间差异和时间变化特征[11,24-25]或少数物种的空间差异方面[26-27],而对于中国近50年温度敏感度的变化情况尚无系统报导。为增强对未来物候变化的预测能力,亟需对过去50年来中国地区植物春季物候期的温度敏感度变化特征及影响因素进行深入研究。本文选择1963—2014年中国中东部地区10个站点163种植物的展叶始期资料,利用滑动分析法计算了每15a各植物的展叶始期温度敏感度。在此基础上,分析了温度敏感度的变化趋势及空间格局,并探讨了导致展叶始期温度敏感度变化的可能原因。
1 数据与方法
1.1 物候与气象数据
本文所用物候观测数据来自“中国物候观测网”(China Phenological Observation Network, http://www.cpon.ac.cn/)。根据观测序列较为连续、物种较丰富的原则,研究选取了中东部地区哈尔滨、北京、长沙等10个站点(表1)。物候数据时间跨度为1963—2014年,各站点观测年数在21—43a之间。为研究温度敏感度的时间变化,筛选出各站点观测年数超过20年的木本植物,共计163种、313条序列。这些植物隶属48个科,103个属,包括66种灌木,97种乔木。由于观测中断,1969—1972和1997—2002年两时段的物候数据缺失,所选序列主要分布于1963—1968、1973—1996和2003—2014年(图1)。各观测站点对应的气象资料从中国气象数据网(http://data.cma.gov.cn/)获取,包括1963—2014年的逐日平均气温数据。对由于迁站等原因造成气温数据不均一的站点(如贵阳、西安等),使用中国气象数据网提供的“中国近50年均一化历史气温数据集”。
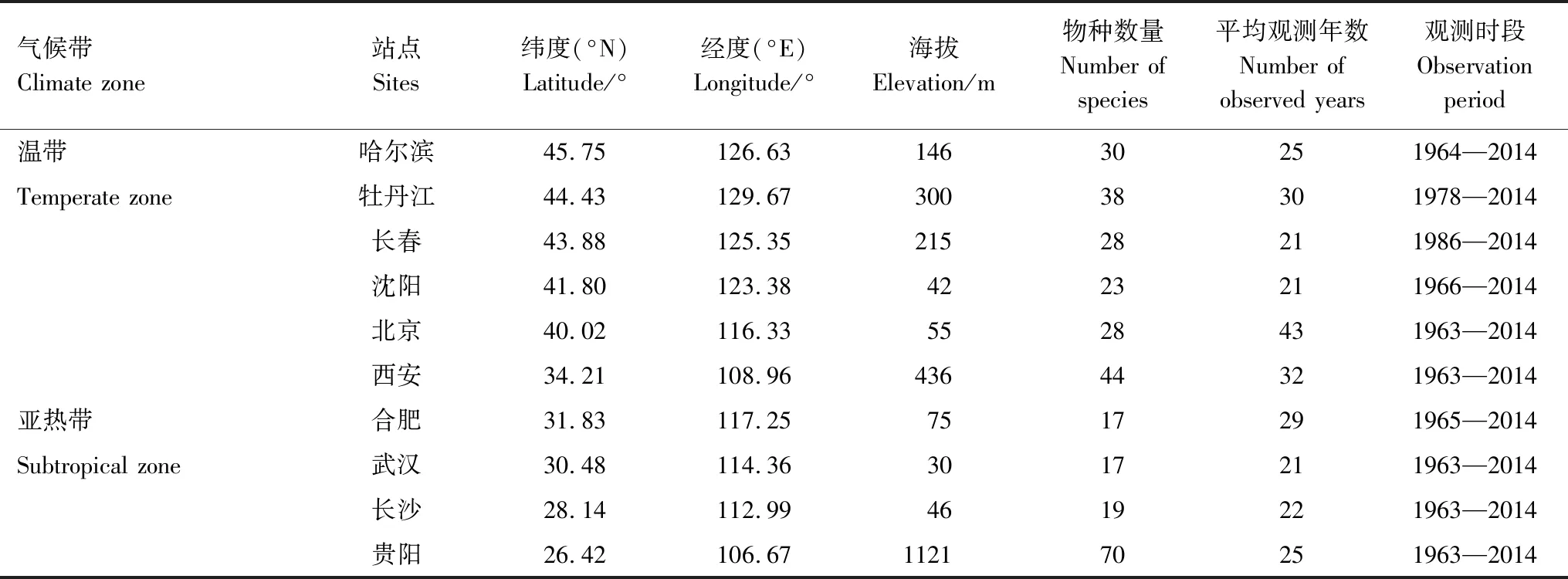
表1 研究站点地理位置及物候资料概况

图1 1963—2014年中国10个站点展叶始期逐年观测记录数量Fig.1 The number of the observation records for leaf unfolding date (LUD) at 10 sites of China from 1963 to 2014
1.2 研究方法
1.2.1温度敏感度计算方法
为估计温度敏感度,首先需要确定温度对植物展叶始期影响最为显著的时段[28-29],即在多年平均展叶始期前一段时间内,以15d为单位滑动计算每一时段(即展叶始期前15d,展叶始期前30d,直至前120d)的平均温度与展叶始期的相关系数,相关系数绝对值最大的时段作为响应时段。展叶始期与响应时段内平均气温的回归方程斜率,可用来表示展叶始期对温度变化响应的敏感度。
为评估展叶始期温度敏感度的变化趋势,本研究首先选择连续的15a时间窗口(例如1963—1977,1974—1988,…,2000—2014),然后对时间窗口内展叶始期序列与响应时段平均气温进行线性回归分析,回归系数用来表征每15a植物的展叶始期温度敏感度[6]。当某一时间窗口内观测年数不足8a时,不予计算。其次,对温度敏感度与时间窗口的终止年份进行线性回归分析,回归系数即为该物种展叶始期温度敏感度的变化速率。由于温度敏感度在大多数情况下为负值,因此温度敏感度在数值上越小意味着绝对值越大,敏感度越高。
1.2.2温度敏感度变化的归因分析
根据引言部分的文献综述,植物展叶始期温度敏感度可能与3个因子有关:植物在冬季受到的冷激量,光周期和春季气温变率。本研究分别用以下变量来表征这3个因子:
(1)冷激天数(CD),用前一年11月1日至平均展叶始期间的平均温度介于0到5℃之间的日数总和来表示[30]。根据本研究的假设,植物在冬季接受的冷激天数降低会导致植物展叶始期温度敏感度降低。
(2)平均展叶始期(LUD)。在固定站点,昼夜长短只与日期有关。当物候期提前到一定日期,昼长缩短,展叶可能受到限制而不再提前。因此,可以用植物平均展叶始期这一变量来表征植物物候发生时的光周期情况。根据本研究的假设,植物展叶始期提前会导致温度敏感度降低。
(3)春季气温标准差(SD),通过计算响应时段内气温标准差的多年平均值来计算[18,20]。根据本研究的假设,春季气温标准差增大,植物温度敏感度降低。
对每条展叶始期序列采用随机抽样法进行归因分析。具体做法是:对每条序列随机选择15a形成一组春季物候期和温度序列,计算该随机序列的温度敏感度以及3个变量(CD,LUD和SD)。将随机抽样实验重复1000次,对随机形成的各时段温度敏感度和各变量进行Pearson偏相关分析。通过分析偏相关系数的显著度和正负号,来判断各因子对温度敏感度的影响是否具有显著性和一致性。
2 结果分析
2.1 展叶始期温度敏感度变化的总体趋势

图2 中国地区313条展叶始期序列温度敏感度变化趋势的频率分布(1963—2014年)Fig.2 Frequency distribution of the trends in temperature sensitivity (ST) for 313 time series of leaf unfolding date (LUD) in China (1963—2014) 虚线表示变化趋势的中值(-0.15d ℃-1 (10a)-1)
从植物展叶始期温度敏感度变化的总体趋势来看,温度敏感度升高的序列明显多于降低的序列。313条展叶始期温度敏感度变化序列中,60.1%的序列呈升高趋势,其中显著的占40.0%(P<0.05);39.9%的温度敏感度序列降低,其中显著降低的占28.4%(P<0.05)(图2)。就变化幅度而言,展叶始期温度敏感度的变化趋势在-7.03d ℃-1(10a)-1(武汉的马尾松Pinusmassoniana)到4.96d ℃-1(10a)-1(长沙的枫香树Liquidambarformosana)之间,中值为-0.15d ℃-1(10a)-1。但81.2%的序列温度敏感度变化趋势绝对值小于1d ℃-1(10a)-1(图2)。
通过对所有展叶始期序列在各个时间窗口内的统计分析发现,1977—2014年展叶始期温度敏感度的平均值和中值呈波动变化。2004年以前主要呈升高趋势,温度敏感度中值从-2.43d/℃增加至-4.08d/℃。2004年后主要呈降低趋势,中值重新降低到约-3.46d/℃(图3)。从线性趋势来看,所有展叶始期序列温度敏感度的中值显著降低,但变化速率较小,仅-0.3d ℃-1(10a)-1(R2=0.64,P<0.05)。

图3 1977—2014年中国地区313条展叶始期序列15年滑动温度敏感度变化Fig.3 Temperature sensitivity of leaf unfolding date for each 15-year moving window of 313 time series in China (1977—2014) 箱底部和顶部代表四分位数;箱内横线代表中值;箱内圆点代表平均值;上下两个须的终端分别代表10和90分位数;红线为中值的线性拟合线:y=-0.03x+53.8, R2=0.64, P<0.05
2.2 展叶始期温度敏感度变化的站点差异

图4 中国10个站点植物展叶始期温度敏感度变化的空间分布Fig.4 The spatial patterns of trends in temperature sensitivity of leaf unfolding date at 10 sites in China
从植物展叶始期温度敏感度变化的空间格局来看,分布在温带地区的6个站点温度敏感度升高的物种数量均超过60%(图4)。其中,北京植物展叶始期温度敏感度升高最为普遍,升高的物种比例达到89.3%。其中显著升高的物种占75.0%(P<0.05),平均变化速率为-0.43d ℃-1(10a)-1。而分布在亚热带温度带的4个站点,除合肥植物展叶始期温度敏感度升高(-0.51d ℃-1(10a)-1)外,其他站点(武汉、长沙和贵阳)均有超过65%的物种展叶始期温度敏感度降低,且显著降低的物种比例在45.7%到68.4%之间。
2.3 展叶始期温度敏感度变化的影响因子
对温带地区站点而言,有66%的展叶始期序列温度敏感度与冷激天数呈负相关关系,其中偏相关系数显著的序列占58.3%(P<0.05);只有10%的展叶始期序列温度敏感度与冷激天数呈显著正相关关系(P<0.05)。这表明,对温带地区大部分植物而言,当冷激天数减少时,展叶始期温度敏感度降低,符合预期假设。对亚热带地区的站点,则有超过一半(58.9%)的序列温度敏感度与冷激天数呈正相关关系,显著的序列达46.8%(P<0.05);只有38.7%序列温度敏感度与冷激天数呈显著负相关关系(P<0.05)。这表明,对亚热带地区大部分植物而言,随着冷激天数减少,展叶始期温度敏感度反而升高,不符合预期假设(图5)。
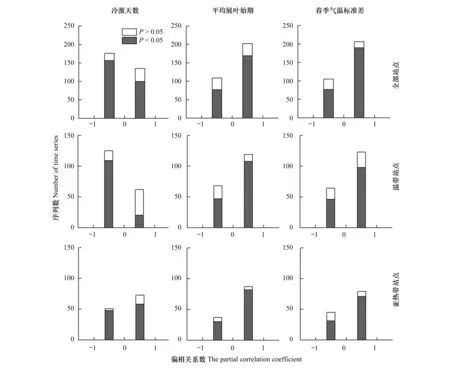
图5 中国地区全部站点以及分区的植物展叶始期温度敏感度与3个因子的偏相关系数频率分布Fig.5 Frequency distributions of the partial correlation coefficients between temperature sensitivity of leaf unfolding date and three factors
在温带和亚热带地区,分别有63.7%和70.2%的序列展叶始期温度敏感度与平均展叶始期呈正相关关系,其中显著的序列分别有57.8%和66.1%(P<0.05)。两个地区分别只有25.1%和24.2%的序列温度敏感度与平均展叶始期呈显著负相关(P<0.05)。因此,无论是在温带地区还是亚热带地区,对大部分植物而言,展叶始期越早,温度敏感度越高,不符合本研究的预期假设(图5)。
与气温标准差的偏相关分析表明,在温带和亚热带地区,分别有65.8%和63.7%的序列展叶始期温度敏感度与春季气温标准差呈正相关关系,其中偏相关系数显著的序列数分别有52.4%和57.2%(P<0.05)。两个地区分别只有24.6%和25%的序列展叶始期温度敏感度与春季气温标准差呈显著负相关关系(P<0.05)。该结果表明,当春季气温变率增大时,大部分植物展叶始期温度敏感度降低,符合预期假设,并且在两个区域的结果一致(图5)。
3 讨论
本研究系统探讨了1963—2014年中国中东部地区10个站点313条展叶始期序列的温度敏感度变化趋势及可能原因。其中,60.1%的序列温度敏感度呈升高趋势,39.9%的温度敏感度序列呈降低趋势。该结果与欧洲地区的研究结果不同。欧洲7种广布种展叶始期平均温度敏感度1999—2013年相较于1980—1994年降低了40%[6]。这表明植物春季物候期的温度敏感度的变化存在显著的区域差异。
研究结果表明,展叶始期温度敏感度与冷激天数的相关关系存在区域差异。在温带地区,66%的植物展叶始期序列的温度敏感度与冷激天数呈负相关关系,表明在温带地区,冷激量的变化能够解释大部分植物温度敏感度变化。然而,在亚热带地区,大部分植物可能不需要或只需要很小的冷激量即可解除芽在秋冬季的休眠[31],冬季低温不足不会使展叶始期温度敏感度降低,表现为58.9%的序列展叶始期温度敏感度与冷激天数呈正相关关系。对于冷激条件变化的响应差异可能是导致温带和亚热带植物展叶始期温度敏感度变化趋势不同的原因。当前,冷激条件对植物叶芽发育的影响仍缺乏深刻认识,需要进一步研究。
根据光周期限制的假设,在气温较高的年份,展叶始期对光周期敏感的植物由于昼长不足的限制而停止提前。因此,展叶始期提前,温度敏感度应降低。但仅有36.4%的温带序列和29.8%的亚热带序列支持这一结论,对于其余63.7%和70.2%的序列而言,随着展叶始期提前,温度敏感度反而升高。因此,光周期限制不能解释大部分植物春季展叶始期温度敏感度的变化。可能的原因是所选植物的春季物候期对光周期变化并不敏感。另一个可能原因是在当前气候状态下,大部分植物春季展叶始期尚未受到光周期限制。在未来气候持续升温的情景下,对光周期敏感的植物可能因到达昼长的阈值而不再提前[32]。
本研究结果还表明无论在温带还是亚热带地区,超过一半的展叶始期序列温度敏感度与春季气温标准差呈显著正相关关系,说明春季气温变率对展叶始期温度敏感度有显著影响,且区域差异性较小。可能的机制是春季温度波动较大的区域更易发生霜冻事件,植物通过降低对气候变化响应的敏感度以减小遭受霜冻灾害的风险[18]。该结果也与温度敏感度空间分布的影响因素一致。Wang等[18]研究发现欧洲不同站点间植物春季物候期温度敏感度与春季气温标准差存在显著正相关关系,在春季气温变率较大的地区植物春季物候敏感度较低。Zhang等[20]通过分析中国59个站点植物花期温度敏感度分布规律也得到类似结论。
4 结论
本研究定量分析了近50年中国中东部地区10个站点163种植物展叶始期温度敏感度的变化趋势及空间格局,并探讨了导致展叶始期温度敏感度变化的可能原因,主要结论为:
(1)在全部313条展叶始期时间序列中,60.1%的序列温度敏感度呈升高趋势,其中显著升高的占40.0%(P<0.05)。39.9%的序列温度敏感度降低,其中显著降低的占28.4%。所有植物温度敏感度的中值在1963—2014年显著升高。
(2)在空间分布上,温带地区的6个站点展叶始期温度敏感度平均呈升高趋势。其中,北京植物展叶始期温度敏感度升高最为普遍,显著升高的物种比例达到75.0%(P<0.05)。而亚热带站点(除合肥外)的展叶始期温度敏感度主要呈降低趋势。其中,长沙植物展叶始期温度敏感度显著降低的物种比例最高,达68.4%。
(3)冬季冷激量和春季气温变率是影响植物展叶始期温度敏感度随时间变化的主要因素。冬季冷激量降低将导致植物展叶始期温度敏感度降低,而春季气温变率降低将导致植物展叶始期温度敏感度升高。
