基于不同方法的毛茛族(毛茛科)导管穿孔板比较研究
2021-03-24罗敏蓉
罗敏蓉
摘 要: 导管分子的演化与植物的进化相关联。毛茛科是联系基部被子植物和核心真雙子叶植物的关键类群,对其导管穿孔板的研究具有重要的系统学意义。为研究毛茛科毛茛族不同属植物导管穿孔板类型及其与环境的相关性,该文利用新的扫描电子显微镜和半薄切片技术,对该族7属8种植物次生木质部导管状分子进行了比较观察。结果表明:木质部管状分子以单穿孔板为主,除了茴茴蒜、鸦跖花和角果毛茛外,其他类群均具有梯状穿孔板;侧金盏花属的蜀侧金盏花导管分子穿孔板的类型多样,有单穿孔板、梯形穿孔板、买麻藤式穿孔板、网状穿孔板和梯-网混合型穿孔板;碱毛茛属的圆叶碱毛茛具单穿孔板、梯形穿孔板和网状穿孔板;在水毛茛属水毛茛的根和茎中未离析出导管,半薄切片显示其具原始的环纹导管。对不同类型导管纹孔膜进行了观察,发现有对列纹孔、互列纹孔,纹孔膜完整或有残余。毛茛族植物导管长度的多样性较高。该文还对不同导管类型的适应性意义和毛茛族的系统位置进行了探讨,揭示了导管分子微形态结构特征与其所处的环境存在一定的适应性。太白美花草、蜀侧金盏花、鸦跖花的导管类型与高寒环境相关,角果毛茛、水毛茛的导管类型分别与其旱生、水生环境相关。
关键词: 毛茛族, 毛茛科, 导管, 穿孔板
Abstract: The evolution of different kinds of vessel molecules is always related to plant evolution. Ranunculaceae (Ranunculales) is a key family which links the basal angiosperm and the core-eudicots in the angiosperm phylogeny group system. Studies on the vessel perforation plates of this important family have great systematic significance, but few studies have been addressed till now. Here, the vessel elements and their perforation plates in the secondary xylem of Ranunculeae, including seven genera and eight species, were observed with the scanning electron microscope and the semithin section, in order to know the relationship betweenthe vessel with their environments in different species in Ranunculaceae of Ranunculeae. The results were as follows: Simple perforation plates were found in seven species, except for the Batrachium bungei; Scalariform perforation plates were found in four species, i.e. Callianthemum taipaicum, Adonis sutchuenensis, Ranunculus sceleratus and Halerpestes cymbalaria, but not for Ranunculus chinensis, Oxygraphis glacialis and Ceratocephalus orthoceras; Perforation plates of vessel elements of Adonis sutchuenensis were diverse, including simple perforation plate, scalariform perforation plate, reticulate perforation plate, gnetum type perforation plate, and scalariform-reticulate perforation plate; Simple perforation plate, scalariform perforation plate, and reticulate perforation plate were found in Halerpestes cymbalaria. In this study, we did not found vessel elements by dissociation the root and stem of Batrachium bungei. On the contrary, we found that there were annular vessels by semithin section. We also found that the side walls, and the pit membranes were unbroken or residual. The length diversities of those vessel elements of Ranunculeae were very high. There were significant differences in the average length of vessel elements in different species of Ranunculeae. At the same time, the differences were conspicious among different types of vessel elements even in the same species. We also discussed the adaptive significance of different types of vessel elements and the systematic position of Ranunculeae in the family Ranuculaceae. The results indicate that micro-structures of the vessel elements are also adapted to their growing environments. The vessel elements of Callianthemum taipaicum, Adonis sutchuenensis and Oxygraphis glacialis were related to their cold environments and those of Ceratocephalus orthoceras and Batrachium bungei were related to xerophytic and aquatic environment, respectively.
Key words: Ranunculeae, Ranunculaceae, vessel elements, perforation plates
植物关键形态解剖学性状的发掘、分析、重新认识与评价一直是植物系统分类学研究的重要内容之一。导管位于维管植物的木质部中,是适应陆生环境而形成的输导水分和矿质元素的植物组织。科学家普遍认为原始被子植物的木质部没有导管,具有导管是被子植物的典型特征 (Young, 1981; Li et al., 2011)。导管在长期的进化过程中,产生了很多不同于管胞的特征:纹孔膜消失,长度缩短,直径增大,大量的导管分子互相连接,穿孔板出现分化等 (Field et al., 2000)。因此,导管比管胞的输导能力更强 (Medeiros et al., 2019)。其中,穿孔板的出现是导管与管胞的关键区别特征,纹孔中纹孔膜的消失是重要的特征 (Bailey, 1944)。
Fahn (1982) 认为植物管状分子的进化与植物类群的进化相关联,导管及其穿孔板特征是显示一个类群系统演化地位的重要解剖学指标之一,具有不同类型穿孔板的导管的输导能力不同。因此,管状分子的结构在分类学和系统学研究中具有重要的意义。Carlquist & Schneider (1998) 曾对天南星科 (Araceae) 植物根和茎的导管进行研究,发现该科植株的生活习性及生态型和导管分子的特化相关联。Carlquist (1980) 曾指出长的导管和湿润的生境相关,短的导管和干旱的生境相关。刘树焕等 (2015)发现苹果亚科不同属植物导管分子形态与环境存在适应性。郑宝江和李国秀 (2015) 研究了茶藨子属不同物种导管分子形态与系统位置及生境的关系。
穿孔板可分为单穿孔板、梯状穿孔板、网状穿孔板、麻黄式和买麻藤式穿孔板等类型,同时,在不同类型穿孔板之间还存在过渡类型。关于被子植物导管穿孔板的进化途径,Thompson (1923) 曾认为单穿孔板起源于梯状穿孔板,由梯状穿孔板失去横隔进化而来,并提出了网状穿孔板的概念,同时指出梯状穿孔板与单穿孔板之间具有过渡类型。Frost (1930a, b)认为穿孔板的进化方向为梯状穿孔板、梯状和网状混合穿孔板、端壁倾斜的单穿孔板,端壁水平的单穿孔板是最高级的类型。Bailey (1944)则认为导管的进化顺序为由横隔多的梯状穿孔板到横隔少的梯状穿孔板,再到单穿孔板。20世纪后期,人们发现了网状穿孔板向单穿孔板的过渡类型。
毛茛科(Ranunculaceae)位于真双子叶植物的基部,是联系基部被子植物和核心真双子叶植物的关键类群 (APG, 2016; Zhai et al., 2019)。对该科木质部导管分子的研究,有助于理解真双子叶植物的基本特征以及科内的系统发育演化关系。毛茛族(Ranunculeae)隶属于毛茛科(Ranunculaceae),包括了约18个属,是该科包含属数较多,也是进化地位较为高级的族(Tamura, 1995; Zhai et al., 2019)。关于毛茛科植物的管状分子,Metcalf & Chalk (1950) 指出,毛茛属(Ranunculus)的维管束被厚壁组织包围,导管具单穿孔板。陈永喆和李正理 (1990) 采用常规压片法,观察了毛茛科的导管分子形态,涉及到毛茛族7属17种植物。其结果表明:单穿孔板普遍存在;除水毛茛属外,均存在梯状穿孔板;仅在侧金盏花属中观察到了买麻藤式穿孔板。Tamura (1995) 指出,毛茛科的茎中导管一般较大、成簇,通常为单穿孔板,有时为梯状穿孔板;在毛茛属中观察到了梯状穿孔板横隔的融解现象,有时单穿孔和梯状穿孔在同一导管的两端出现。
由于研究方法的限制,以往的研究结果均采用常规压片和光学显微镜观察。扫描电子显微镜技术相对于普通显微镜具有分辨率更高、放大倍数更大、更逼真的特点,可以展示导管分子精细的三维结构,而半薄切片技术可以展示導管清晰的切面,对于导管的研究具有重要的意义。在水青树、昆栏树等无导管的类群中发现了导管的存在 (Li et al., 2011)、番荔枝科(Annonaceae)的导管处于较高的演化水平 (李红芳和张小卉, 2018)。目前在毛茛科中利用扫描电子显微镜 (SEM)只对类叶升麻属(Actaea)(张阿娟等, 2005)、黄三七属(Souliea)(宋春慧等, 2006)根木质部中的管状分子进行了详细的报道,阻碍了我们对全科管状分子多样性和演化规律的理解。毛茛族植物导管分子的特征是什么,该族不同植物的管状分子类型与其生长环境以及系统位置是否存在相关性,是我们关心的两个科学问题。本文利用扫描电子显微镜法和半薄切片法,对毛茛族7属8种植物的管状分子进行比较观察,以期获得该类群较全面的导管分子特征,为该类群的系统演化演化提供解剖学证据。
1 材料与方法
实验材料采于2006—2010年,将采集的成熟的根和茎以FAA 固定液 (甲醛∶冰醋酸∶50%乙醇=5∶5∶90) 固定,采集信息如表1。
将用作扫描电镜观察的材料首先用KQ-500DE超声波清洗仪清洗,Jeffrey 离析液 (10%铬酸∶10%硝酸=1∶1) 室温下离析24~38 h,离析材料涂片,自然干燥后利用JS-1600镀膜仪喷金镀膜,在日立HITACHI S-3400扫描电子显微镜 (SEM) 下观察照相。将用作解剖学的材料以酒精系列脱水,环氧丙烷置换,环氧树脂包埋,超薄切片机切片,甲苯胺兰染色,环氧树脂封片,Nikon SMZ25体式显微镜观察并照相。每种植物测量120个导管分子的长度, 并计算平均值和标准误差。
2 研究结果
2.1 太白美花草
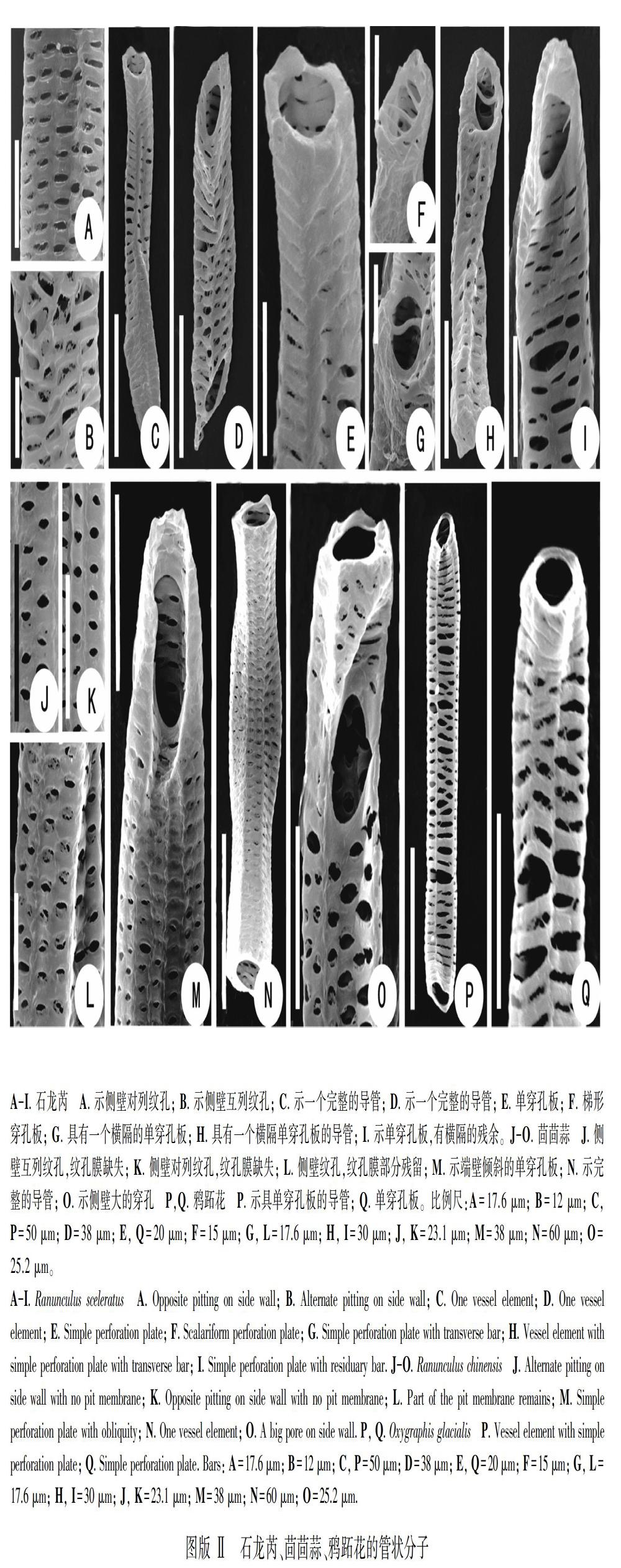
太白美花草导管端壁穿孔板主要有两种类型:梯形穿孔板 (图版Ⅰ:A) 和单穿孔板 (图版Ⅰ:B-D),单穿孔板比例较高,约占79%,平均长度也较长 (162.97 μm)。其端壁倾斜 (图版Ⅰ:B) 或端壁水平 (图版Ⅰ:C, D);个别单穿孔板具较长的喙 (图版Ⅰ:B)。
2.2 蜀侧金盏花
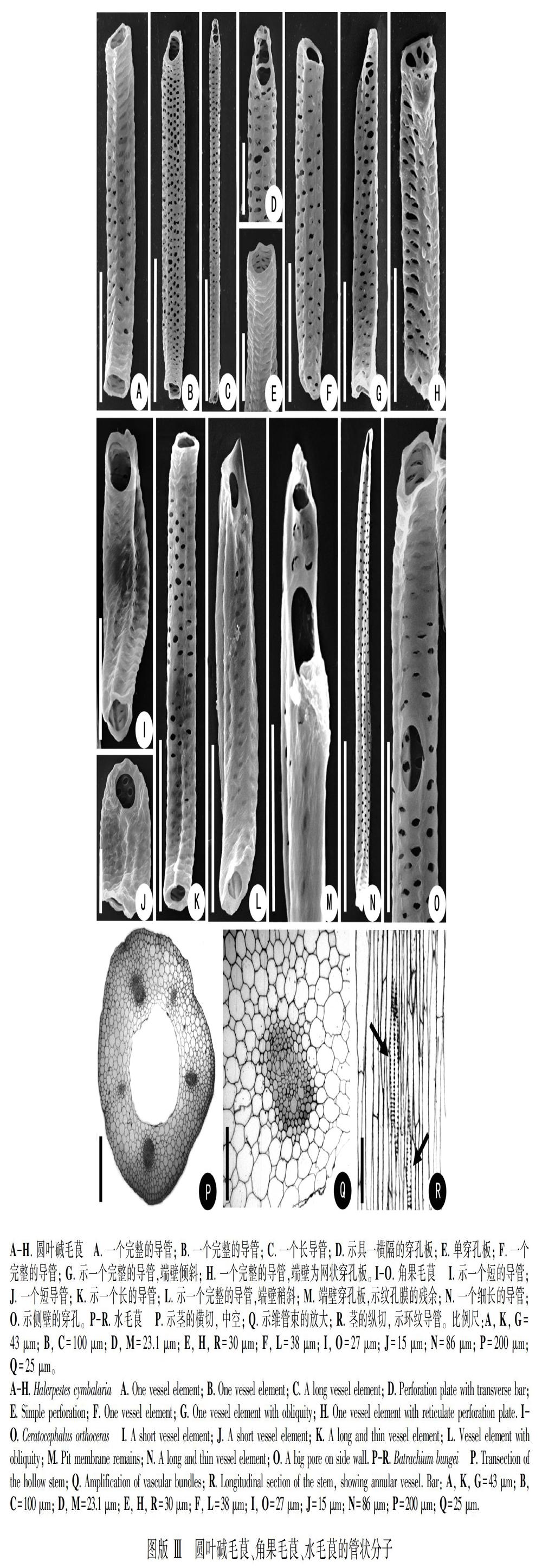
蜀侧金盏花导管端壁穿孔板类型较多:单穿孔板约占46% 、平均长度121.5 μm (图版Ⅰ:E, G),个别具喙 (图版Ⅰ:G);梯形穿孔板约占25%、平均长度154.82 μm (图版Ⅰ:I-L),其中有横隔为一条(图版Ⅰ:I),横隔为两条 (图版Ⅰ:K) 或横隔为三条 (图版Ⅰ:L);一条横隔具有分叉的 (图版Ⅰ:J);梯形-网状混合穿孔板约占8% (图版Ⅰ:M, N);买麻藤式穿孔板比例较低,约占3% (图版Ⅰ:F, H),边缘具一圈小孔。
2.3 石龙芮
石龙芮导管端壁穿孔板主要有以下类型:单穿孔板 (约占68%,平均长度98.86 μm,图版Ⅱ:C-E);梯形穿孔板 (约占32%,平均长度78.91 μm,图版Ⅱ:F-I),其中横隔为一条(图版Ⅱ:G, H),多条 (图版Ⅱ:F),或为横隔退化 (图版Ⅱ:I)。石龙芮侧壁纹孔差异较大:有对列纹孔 (图版Ⅱ:A) 或互列纹孔 (图版Ⅱ:B);纹孔膜完整 (图版Ⅱ:A)或有残余 (图版Ⅱ:B)。
2.4 茴茴蒜
茴茴蒜导管端壁全部为单穿孔板,平均长度181.53 μm,主要有以下类型:端壁倾斜的单穿孔板 (图版Ⅱ:M),端壁水平的单穿孔板 (图版Ⅱ:N);有时单穿孔板出现在侧壁 (图版Ⅱ:O)。
茴茴蒜侧壁纹孔差异较大:有对列纹孔,纹孔膜破损 (图版Ⅱ:K),或者对列纹孔具有较完整的纹孔膜 (图版Ⅱ:L),以及互列纹孔 (图版Ⅱ:J)。
2.5 鸦跖花
鸦跖花导管端壁穿孔板只有单穿孔板一种类型 (图版Ⅱ:P, Q),平均长度137.65 μm;端壁水平 (图版Ⅱ:Q) 或倾斜 (图版Ⅱ:P)。
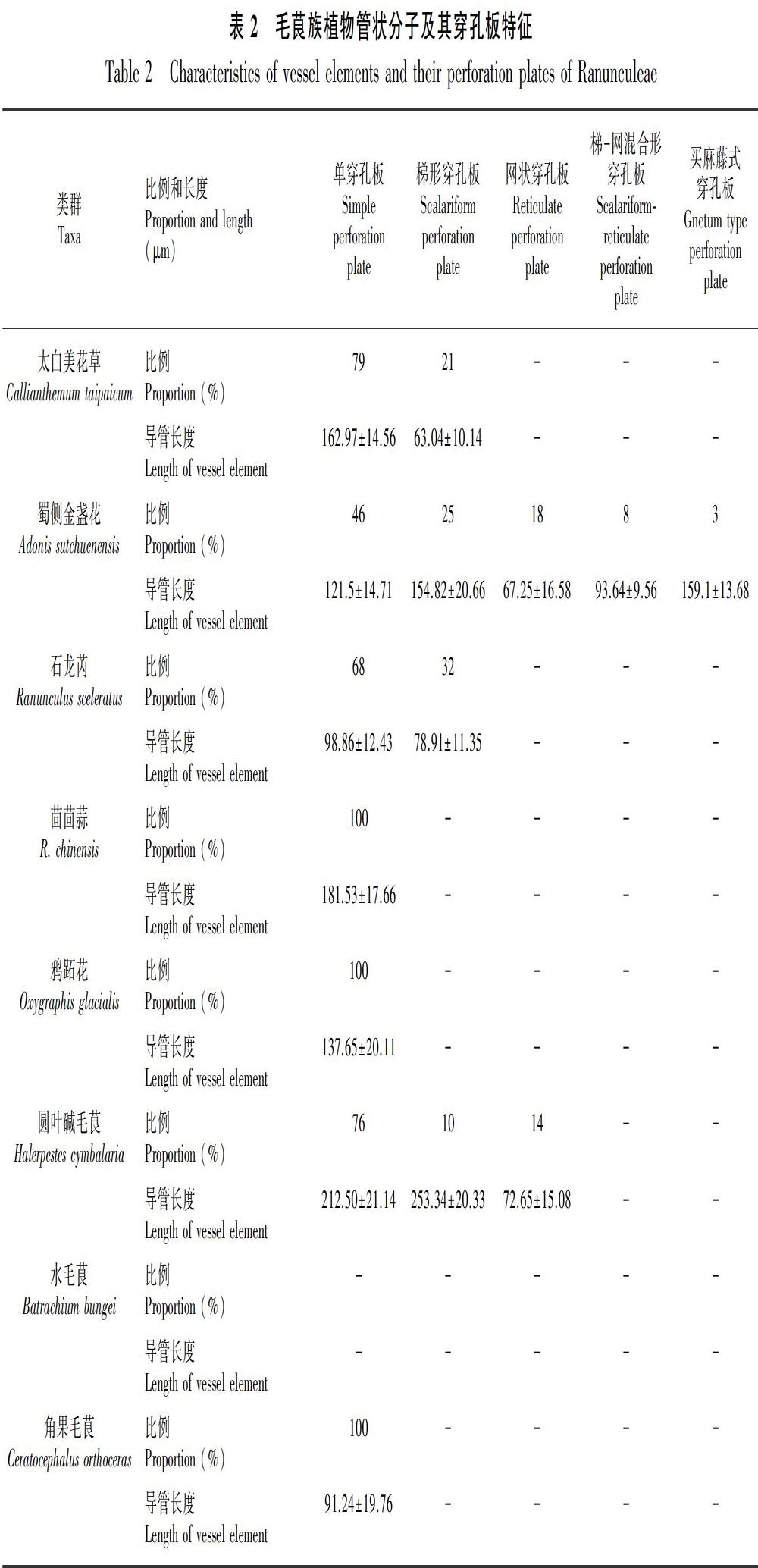
2.6 圆叶碱毛茛
圆叶碱毛茛导管端壁穿孔板主要有以下类型:单穿孔板 (约占76%,平均长度212.5 μm,图版Ⅲ:A, B, E);一端为单穿孔板,另一端为梯形穿孔板 (约占10%,平均长度253.34 μm,图版Ⅲ:C, D);一端为单穿孔板,另一端为网状穿孔板 (约占14%,平均长度72.65 μm,图版Ⅲ:F-H)。
2.7 角果毛茛
角果毛茛导管端壁穿孔板只有单穿孔板一种类型 (图版Ⅲ:I-O),粗短 (图版Ⅲ:I, J) 或细长 (图版Ⅲ:K, L, N),平均长度91.24 μm;侧壁有时出现单穿孔(图版Ⅲ:M, O)。
2.8 水毛茛
在水毛茛的根和茎均没有成功地离析出导管,通过切片发现其导管为较原始的环纹导管(图版Ⅲ:P-R)。
3 讨论与结论
3.1 关于毛茛族植物管状分子的形态与进化
具有穿孔板是导管分子的主要特征之一,分为单穿孔板和复穿孔板两大类型。毛茛族植物的导管分子特征见表2,该族植物的管状分子以单穿A-D. 太白美花草 A. 示网状穿孔板; B. 单穿孔板; C. 示一个完整的导管; D. 示一个完整的导管。 E-N. 蜀侧金盏花 E. 示一个完整的导管; F. 一个导管两端均为买麻藤式穿孔板; G. 单穿孔板; H. 买麻藤式穿孔板; I. 示具有一条横隔的穿孔板; J. 示横隔有分叉的穿孔板; K. 具两条横隔的穿孔板; L. 梯形穿孔板; M. 梯网混合形穿孔板; N. 梯网混合形穿孔板。比例尺:A=13.6 μm; B, I, J=20 μm; C, E=50 μm; D=60 μm; F=43 μm; G=27 μm; H=25 μm; K, L, N=30 μm; M=23.1 μm。
管类型最丰富的类群。在毛茛族中,仅在蜀侧金盏花中发现了买麻藤式穿孔板,而在矮金莲花、天葵、鸡爪草、拟扁果草、耳状人字果等物种中也报道有买麻藤式穿孔板 (陈永喆和李正理, 1990)。在本研究中,蜀侧金盏花和圆叶碱毛茛中发现有网状穿孔板,该类型在毛茛科的黄三七和类叶升麻中也有报道(张阿娟等, 2005;宋春慧等, 2006)。
本研究结果显示,毛茛族植物导管长度的多样性较高:不同物种导管的平均长度差异显著,同一物种内不同类型导管的长度差异也较大。圆叶堿毛茛具梯形穿孔板的导管最长,其具单穿孔板的导管也较长,但其具网状穿孔板的导管长度却较短。在太白美花草中,具单穿孔板的导管长度较长,但其具梯形穿孔板的导管在所有研究的导管类型中长度最短。蜀侧金盏花的导管类型多样,但导管长度变化幅度较小。石龙芮具单穿孔板和梯形穿孔板的导管长度相近。尽管茴茴蒜、鸦跖花、角果毛茛均具有单穿孔板的导管,但其长度多样性也很高。
Frost (1930a, b) 认为,如果一种植物的导管类型比较原始,那么这种植物的系统位置也是原始的。毛茛族植物次生木质部中导管分子的穿孔板以单穿孔板为主,同时存在着梯状、网状穿孔板,并且在网状、梯状穿孔板上的穿孔中有纹孔膜残留这一具有过渡性的指标。在升麻属、乌头属、耧斗菜属、Hydrastis和Xanthorhiza中均发现具有梯形穿孔板的导管分子,但在较原始的星果草属中却发现单穿孔板的导管分子(陈永喆和李正理, 1990)。因此,就导管分子穿孔板这一性状而言,毛茛族代表了毛茛科在系统发育位置上较为高级的类群,这与该科最新的系统发育基因组学的研究结果一致(Zhai et al., 2019)。
3.2 导管与植物生境的联系
在植物木质部中,结构的差异是功能适应于生长环境而不断进化的结果,与水分的利用、蒸腾作用以及机械强度的需要等因素有关 (Carlquist, 1975)。导管官腔的长度、宽度与生境密切相关的。Carlquist & Schneider (1998) 认为干旱加速导管的进化,在温暖环境下植物的导管长而粗 (Pool & Kfrancis, 2000),而窄和短的导管在寒冷的地方更具优势 (Sperry, 2003)。Zimmermann (1978, 1982) 认为导管结构与水分运输的有效性和安全性有关:窄的导管输导效率低,但抗负压能力较强,而宽的导管输导效率较高,但易被挤压变形。对毛茛科高山植物黄三七的导管研究表明,该类群木质部中导管以长度较短的单穿孔板为主,网状、网状-梯形、网状-梯形-单穿孔混合型穿孔板同时存在,并有纹孔膜残余,表明黄三七是毛茛科系统发育上相对高级,但具过渡性的类群,同时展示出适应高海拔环境的特征 (宋春慧等, 2006)。
太白美花草、蜀侧金盏花、鸦跖花也属于典型的高山植物,其木质部长度较短的导管可以较为有效地运输水分,表现出更好地适应高寒环境的特征,这与黄三七属植物适应高海拔环境的导管特征是相似的 (宋春慧等, 2006)。角果毛茛为典型的旱生植物,其木质部由口径略大而短和口径略细而长的导管共同构成,其管状分子类型表现出与旱生环境的高度适应:大口径的导管可以在雨季有效地运输水分,但由于其口径宽、抗负压的能力较弱,在干旱缺水时容易被形成的气泡堵塞;尽管窄口径的导管输送效率较低,但其抗负压力强,细小的官腔不易形成气泡,保证了运输的安全性 (Bass et al., 1983; Zimmermann, 1993)。关于水毛茛的管状分子,由于其长期生长于水环境中 (Tamura, 1995),不再需要复杂而高级的导管来供应植物的水吸收,导管为较原始的环纹导管。同时,茎中薄壁细胞口径较大、中央具有空腔,这些特征均是在水生环境中所表现出来的适应性性状。因此,毛茛族植物不同的导管结构差异是其功能长期适应不同环境的结果。
参考文献:
APG IV, 2016. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV [J]. Bot J Linn Soc, 181(1): 1-20.
BAILEY IW, 1944. The development of vessels in angiosperms and its significance in morphological research [J]. Am J Bot, 31(7): 421-428.
BASS P, WERKER E, FAHN A, 1983. Some ecological trends in vessel characters [J]. Iawa J, 4(2): 141-159.
CARLQUIST S, 1975. Ecological strategies of xylem evolution [M]. NewYork: University of California Press.
CARLQUIST S, 1980. Further concepts in ecological wood anatomy, with comments on recent work in wood anatomy and evolution [J]. Aliso, 9(4): 499-553.
CARLQUIST S, SCHNEIDER EL, 1998. Origin and nature of vessels in monocotyledons. Araceae subfamily Colocasioideae [J]. Bot J Linn Soc, 128(1): 71-86.
CHEN YZ, LI ZL, 1990. Comparitive studies of perforation plate structures of vessels in Ranunculaceae [J]. Acta Bot Sin, 32(4): 245-251. [陈永喆, 李正理, 1990. 毛茛科植物导管穿孔板结构的比较观察 [J]. 植物学报, 32(4): 245-251.]
FAHN A, 1982. Plant anatomy [M]. 3rd ed. Oxford: Pergamon Press.
FIELD TS, ZWEINIECHI MA, BRODRIBB T, et al., 2000. Structure and function of tracheary elements in Amborella trichopoda [J]. Int J Plant Sci, 161(5): 705-712.
FROST FH, 1930a. Specialization in secondary xylem of dicotyledon: I. Origin of vessel [J]. Bot Gaz, 89(1): 67-94.
FROST FH, 1930b. Specialization in secondary xylem in dicotyledons: Ⅱ. Evolution of end wall of vessel segments [J]. Bot Gaz, 90(2): 198-211.
LI HF, CHAW SM, DU CM, et al., 2011.Vessel elements present in the secondary xylem of Trochodendron and Tetracentron (Trochodendraceae) [J]. Flora, 206(6): 595-600.
LI HF, ZHANG XH, 2018. Comparative observation on vessel elements and perforation plates in four species of Annonaceae [J]. Guihaia, 38(5): 665-671. [李紅芳, 张小卉, 2018. 四种番荔枝科 (Annonaceae) 植物导管分子及其穿孔板的比较观察 [J]. 广西植物, 38(5): 665-671.]
LIU SH, GU LW, ZHANG DW, 2015. Morphological structure of vessel elements in 14 species of Maloideae (Rosaceae) from Heilongjiang, China [J]. Acta Bot Oreal-Occident Sin, 35(4): 723-729. [刘树焕, 谷利伟, 张大维, 2015. 黑龙江苹果亚科5属14种植物导管分子形态结构研究 [J]. 西北植物学报, 35(4): 723-729.]
MEDEIROS JS, LENS F, MAHERALI H, et al., 2019. Vestured pits and scalariform perforation plate morphology modify the relationships between angiosperm vessel diameter, climate and maximum plant height [J]. New Phytol, 221(4): 1802-1813.
METCALFE CR, CHALK L, 1950. Anatomy of the Dicotyledons [M]. Oxford : Clarendon Press.
POOL EI, KFRANCIS JE, 2000. The first record of fossilwood of Winteraceae from the upper cretaceous of Antarctica [J]. Ann Bot, 85(3): 307-315.
SONG CH, REN Y, TIAN XH, et al., 2006. An investigation of vessel perforation plates in the secondary xylem of stem of Souliea vaginata (Ranunculaceae) [J]. Acta Bot Yunnan, 28(2): 157-161. [宋春慧, 任毅, 田先华, 等, 2006. 黄三七(毛茛科) 茎次生木质部导管穿孔板的研究 [J]. 云南植物研究, 28(2): 157-161.]
SPERRY JS, 2003. Evolution of water transport and xylem structure [J]. Int J Plant Sci, 164(3): 115-127.
TAMURA M, 1995. Phylogeny and classification of the Ranunculaceae [J]. Plant Syst Evol, (Suppl.) 9(9): 201-206.
THOMPSON WP, 1923. The relationships of the different types of angiospermic vessels [J]. Ann Bot, 37(166): 183-191.
YOUNG DA, 1981. Are the angiosperms primitively vesslelless? [J]. Syst Bot, 6(4): 313-330.
ZHAI W, DUAN XS, ZHANG R, et al., 2019. Chloroplast genomic data provide new and robust insights into the phylogeny and evolution of the Ranunculaceae [J]. Mol Phylogenet Evol, 135(6): 12-21.
ZHANG AJ, REN Y, TIAN XH, et al., 2005. Tracheary elements in secondary xylem of Actaea asiatica (Ranunculaceae) [J]. Acta Bot Boreal-Occident Sin, 25(12): 2363-2370. [张阿娟, 任毅, 田先华, 等, 2005. 类叶升麻(毛茛科)次生木质部管状分子的研究 [J]. 西北植物学报, 25(12): 2363-2370.]
ZHENG BJ, LI GX, 2015. Morphological observation of vessel elements in secondary xylem of Ribes [J]. J Beijing For Univ, 37(11): 48-58. [郑宝江, 李国秀, 2015. 茶藨子属次生木质部导管分子形态学研究 [J]. 东北林业大学學报, 37(11): 48-58.]
ZIMMERMANN MH, 1978. Hydraulic architecture of some diffuse-porous trees [J]. Can J Bot, 56(18): 2286-2295.
ZIMMERMANN MH, 1982. Functional xylem anatomy of angiosperms trees [M]//BAAS P. New perspectives in wood anatomy. Dordrecht: NijhoffJunk Publishers.
ZIMMERMANN MH, 1993. Xylem structure and the ascent of sap [M]. Berlin: Springer-Derlag: 2-20.
(责任编辑 李 莉)
