入侵恶性杂草豚草和三裂叶豚草土壤种子库特征及其对地上种群的贡献
2021-03-15王瑞丽董合干刘彤赵文轩王寒月马倩倩刘延
王瑞丽,董合干,2,刘彤,赵文轩,王寒月,马倩倩,刘延
(1 石河子大学生命科学学院,新疆 石河子 832003;2 伊犁州农村能源环境工作站,新疆 伊宁 835000)
豚草(Ambrosiaartemisiifolia)和三裂叶豚草(Ambrosiatrifida)同为菊科豚草属一年生恶性入侵杂草,具有生命力强、适应性强、种子产量多、扩散速度快等特征。二者的入侵破坏生态环境,降低生物多样性,入侵农田会大幅降低作物产量[1]。自1935年起,在我国东北首先发现豚草和三裂叶豚草[2-4],目前广泛分布于东北、华北、华中和华东等地区[5-6]。2010年在新疆伊犁河谷发现豚草和三裂叶豚草的入侵,2014年呈爆发式蔓延,广泛分布于路边、农田、景区及牧场,对当地生态环境、农业和畜牧业生产造成了巨大影响[7]。
在自然状态下,豚草和三裂叶豚草种群呈单一的集群分布。其具有叶腋处结实的特性,单株种子产量高,可达 2 000~62 000粒[8-9]。在野外调查过程中我们发现,其高大植株在种子散布过程中不倒伏,绝大部分种子散布在母株丛附近,散布格局呈聚集型,容易形成庞大的土壤种子库,因此成为入侵防控的棘手问题。地上植被是土壤种子库的来源,而土壤种子库作为种源储备也通过萌发和成苗参与地上植被的更新[10-11],研究土壤种子库种子命运是深入研究物种的生态适应及其种群对策的基础。土壤种子库的研究涉及到的植被类型较多,包括对森林群落[12-13]、农田杂草植被[14-15]、草原群落[16-17]、沼泽湿地[18-19]和荒地植被及干旱区荒漠植被[20-22]等各种不同生境。研究的内容也非常丰富,如:土壤种子库的分布结构和组成[23-25]、在森林更新过程中的作用[26]、种子库与种子雨、地上植被及幼苗的关系[27]、休眠和萌发特征[28]、在植被恢复中的作用[29-31]等。对于豚草和三裂叶豚草等一年生植物,土壤种子库与地上植被具有较密切的相似性[32-33],所以研究其土壤种子库特征,有助于了解土壤种子库与其种群建立、增长和持续性的关系,进而对两个物种有效防控技术的形成起到一定指导作用。
研究显示,豚草在多种入侵物种中的频度高达92%,这是因为其堆积土壤种子库中的种子特别丰富,且发芽率高,因此其传播速度和扩散速度非常之快[34],由此可见种子库的相关生物学研究十分必要。有学者提出通过物理清除豚草的地上部分植被,其种子数量就能明显减少[35]。另外一项关于种子库种子量减少的报道显示,在越冬期间昆虫等动物会将三裂叶豚草的种子吃掉近50%[36]。目前,种子库中其它相关研究仍未深入,而两个物种土壤种子库命运的研究还未见报道。
本研究以2010年开始至今大面积入侵新疆伊犁河谷新源县的豚草和三裂叶豚草为对象,通过对比和分析不同生育期豚草和三裂叶豚草地上植被密度变化及土壤种子库中的种子命运变化,以期深入了解二者在入侵过程中的种群适应和种群调节,从而为研究豚草和三裂叶豚草的入侵机制以及科学防控提供理论依据。
1 材料与方法
1.1 研究区概况
采样地点位于豚草和三裂叶豚草的入侵地——新疆维吾尔自治区伊犁州新源县,地理位置为82°28′~84°56′E,43°03′~43°40′N。新源县属温带大陆性半干旱气候,年均气温6.0 ℃~9.3 ℃,年平均降水476 mm。该地水、生物以及矿产等自然资源丰富,地形地貌主要由平原、丘陵、山地构成。
在我们前期研究基础上[7],选择豚草和三裂叶豚草入侵2、4、6年的不同生境。豚草入侵生境主要位于农田,三个生境植被相似,主要伴生物种为禾本科(Poaceae)的剪股颖(Agrostismatsumurae)、稗(Echinochloacrusgalli)、狗尾草(Setariaviridis);菊科(Compositae)的苍耳(Xanthiumsibiricum)、蒲公英(Taraxacummongolicum)、菊苣(Cichoriumintybus)和藜科(Chenopodiaceae)的藜(Chenopodiumalbum)。
三裂叶豚草入侵生境主要位于草场,3个生境植被相似,主要伴生物种除了上述提到的禾本科和菊科植物外,还有豆科(Leguminosae)的紫苜蓿(Medicagosativa)、红车轴草(Trifoliumpretense)、苦豆子(Sophoraalopecuroide);桑科(Moraceae)的大麻(Cannabissativa)和伞形科(Umbelliferae)的野胡萝卜(Daucuscarota)等。
1.2 地下土壤种子库取样方法
豚草研究地位于新源县则克台镇的农田,三裂叶豚草研究地位于新源县土尔根乡的草场。由于二者各自入侵地的微生境相似,采用随机取样法进行土壤种子库(持久种子库)的取样。土壤种子库于2019年5月初统计幼苗期种群密度后采集,不同生境样地选取20 cm×20 cm的样方各6个,将土层深度为0~10 cm、10~20 cm、20~30 cm的土壤全部取样带回,用筛分法(2 mm筛网)统计样方内种子数,做种子库密度分析。其中,将饱满、未被啃食的种子统计为完好种子,未发育完全的种子统计为干瘪种子,开裂且完整的种皮统计为萌发后蜕掉的种壳,有孔洞、被啃食的种子统计为虫食种子,有腐烂或霉菌痕迹的种子统计为霉变种子,统计各类种子的数量及在其总种子数中的占比。
1.3 地上部分种群密度调查方法
2019年5月初,即豚草和三裂叶豚草幼苗生长期,对不同生境(分别入侵2、4、6年的生境)单位面积内的种群密度进行调查统计。每个生境随机取6个1 m×1 m的样方,统计样方内的全部植株数量,以6个样方种群密度的平均值作为其种群密度估值。同年,在营养期(7月)、繁育期(9月)、成熟期(10月)按照同样方法再测量一次,测定种群密度的变化情况。
1.4 数据分析
将土壤种子库取样面积的种子数目换算成1 m2面积的数量作为种子库的种子密度,计算各类型种子在总种子数中的占比。将种子库密度作为变量进行单因素方差分析(One-way ANOVA),不同土层深度和不同生境间的差异显著性用Duncan多重比较检验(Duncan’s multiple range test),置信区间95%。将种群密度作为变量进行单因素方差分析,不同生境和土层深度间的差异显著性用Duncan多重比较检验,置信区间95%。利用SPSS19.0软件统计分析本试验所得数据,利用origin9.0软件进行作图。
2 结果
2.1 土壤种子库不同类型种子的分布格局
在筛选土壤种子库种子过程中,我们发现豚草入侵生境的种子量较少,且仅在入侵六年生境的0~10 cm土层深度土壤中统计到种子数(表1),这可能与豚草主要入侵的是干扰较为严重的田间地头以及豚草种子质量较小有关。其中,完整种子和萌发种壳分别占该土层深度土壤种子库总种子数的150粒·m-2和250粒·m-2,虫食种子和干瘪种子共125粒·m-2,占种子库总种子的23.81%,未发现霉变种子。
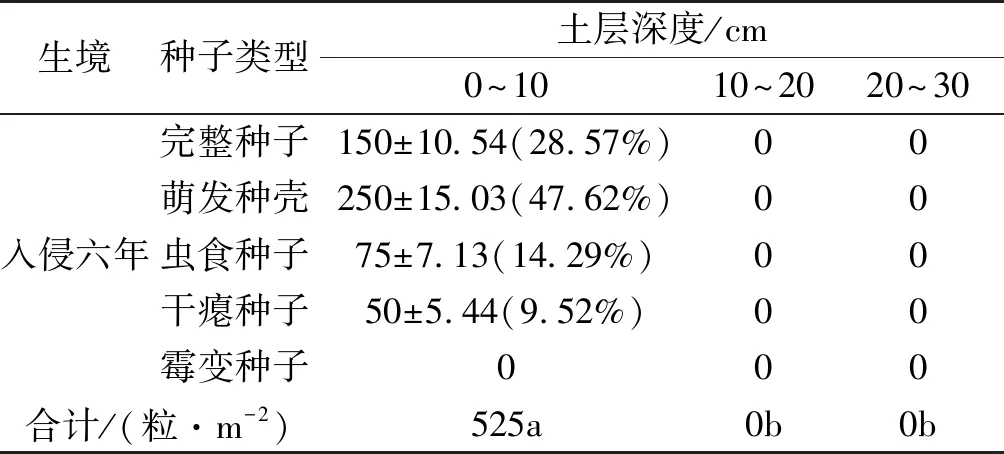
表1 不同土层豚草土壤种子库的种子密度和比例
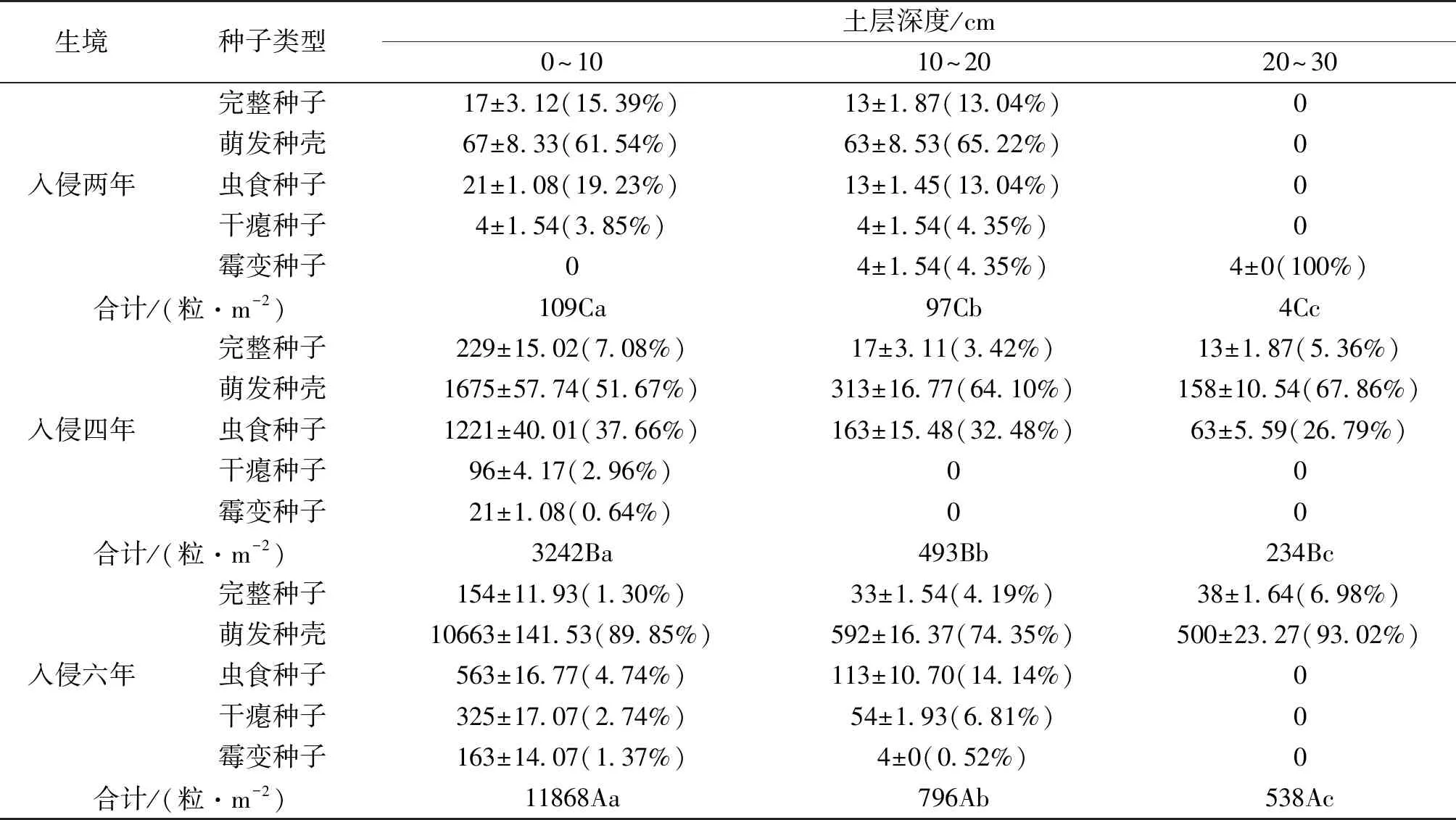
表2 不同土层三裂叶豚草土壤种子库的种子密度和比例 粒·m-2
不同土层深度的三裂叶豚草种子库大小依次为:0~10 cm>10~20 cm>20~30 cm,随土层深度增加,其种子数量显著变少。不同生境种子库大小依次为:入侵六年>入侵四年>入侵两年,入侵时间越久,该生境土壤种子库种子密度越大,且差异显著。由表2可知,入侵两年生境的三裂叶豚草种子库0~10 cm土层同样未发现霉变种子,其余类型种子共109粒·m-2,10~20 cm土层完整种子、萌发种壳、虫食种子、干瘪种子和霉变种子数量分别为13、63、13、4、4粒·m-2,20~30 cm土层仅发现4粒霉变种子;入侵四年生境0~10 cm土层中萌发种壳和虫食种子较多,分别为1 675粒·m-2和1 221粒·m-2,其次是完整种子,为229粒·m-2,干瘪种子和霉变种子数量较少,共占该土层总种子数的3.60%。10~20 cm和20~30 cm土层中均未发现干瘪种子和霉变种子,其余类型种子中,萌发种壳数最多(分别为313、158粒·m-2),虫食种子数次之(分别为163、63粒·m-2),完整种子最少(分别为17、13粒·m-2);入侵六年生境0~10 cm土层中的种子数是所有生境及各土层中最多的,萌发种壳高达上万粒(10 663粒·m-2),完整种子、虫食种子、干瘪种子、霉变种子分别为154、563、325、163粒·m-2,其中后三者共占该土层种子库总种子的8.85%。10~20 cm土层中的总种子数为0~10 cm土层的6.70%,其中,萌发种壳数>虫食种子数>干瘪种子数>完整种子数>霉变种子数,虫食、干瘪、霉变种子数共占该土层总种子数的21.47%。20~30 cm土层仅发现完整种子和萌发种壳,其中完整种子仅38粒·m-2,占该土层总种子数的6.98%。
2.2 土壤种子库种子命运
从图1可以看到,豚草入侵六年生境土壤种子库中不同类型种子所占比例:萌发种壳>完整种子>干瘪种子>虫食种子,分别占总种子数的47.62%、28.57%、14.29%和9.52%。
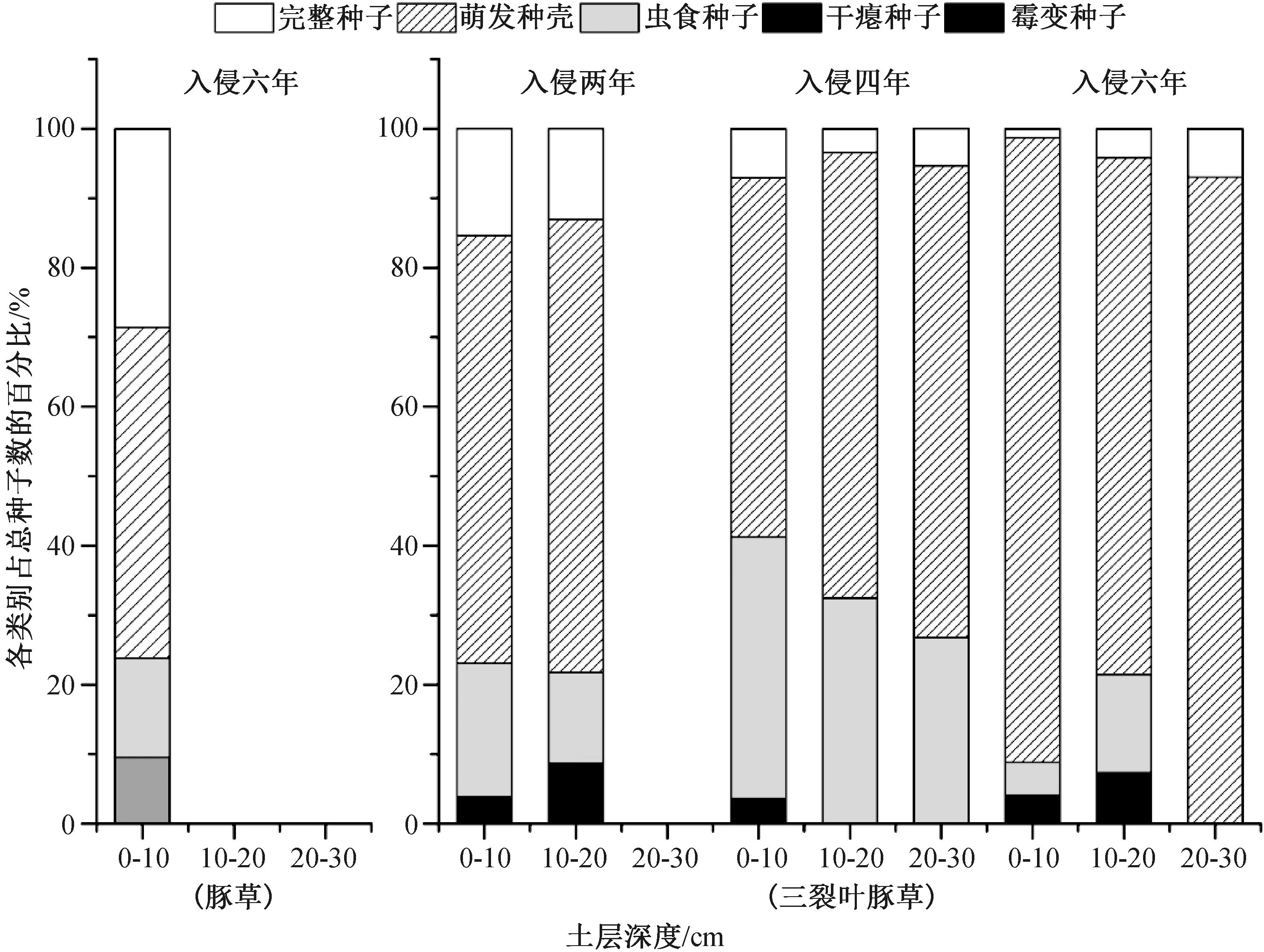
图1 不同生境的豚草和三裂叶豚草土壤种子库中各类型种子比例情况
三裂叶豚草不同类型种子命运分析结果表明,随入侵年限的增加,萌发种壳和虫食种子的比例逐渐增大,完整种子、干瘪种子和霉变种子的比例逐渐变小。土层越深种子类型越少,其中,入侵两年生境10~20 cm土层土壤种子库种子类型最多,包括萌发种壳、完整种子、干瘪种子、虫食种子、霉变种子,分别占总种子数的13.04%、65.22%、13.04%、4.35%和4.35%;0~10 cm土层中无霉变种子,完整种子占总种子数的15.39%,萌发种壳占总种子数的61.54%,虫食和干瘪种子分别占19.23%和3.85%;20~30 cm土层内未发现三裂叶豚草种子。
三裂叶豚草入侵四年和入侵六年生境下的种子库对比发现,随土层深度的增加种子类型减少。在入侵四年生境的0~10 cm土层中萌发种壳、完整种子、干瘪种子、虫食种子、霉变种子分别占总种子数的7.08%、51.67%、37.66%、2.96%和0.64%。
入侵六年生境0~10 cm土层中萌发种壳的比例有所上升,为该土层总种子数的89.85%,其余各类型种子占比相应降低;在入侵四年生境的10~20 cm和20~30 cm土层中均未发现干瘪种子和霉变种子,完整种子分别为各土层中总种子数的3.42%和5.36%,萌发种壳分别为64.10%和67.86%,虫食种子分别为32.48%和26.79%;入侵六年生境的20~30 cm土层中仅有完整种子和萌发种壳,分别为该土层总种子数的6.98%和93.02%;各土层中种子数仍为萌发种壳最多,虫食种子次之,其余类型种子总计占比仅为总种子数的10%左右。
2.3 不同生育期种群密度的变化
由图2可知,不同生境(入侵二、四、六年生境)的豚草和三裂叶豚草种群密度随生育期的延长而下降,依次是:幼苗期>营养期>繁育期>成熟期。入侵时间越久的生境种群密度越高,依次分别是:入侵六年>入侵四年>入侵两年。
从幼苗期到成熟期,两物种的种群密度有剧烈变化,与幼苗期相比,进入成熟期的豚草种群密度分别减少了99、119、406株·m-2,三裂叶豚草分别减少了93、480、698株·m-2。幼苗期至营养期阶段是豚草种群植株密度大幅下降阶段,不同生境豚草幼苗期种群密度分别为115、145、443株·m-2,到了营养期依次减少了50、59、343株·m-2,到了繁育期种群密度已相对稳定,和成熟期无显著差异,均低于50株·m-2。三裂叶豚草入侵四年、六年的生境种群在幼苗期至繁育期阶段植株密度持续呈大幅下降趋势,在幼苗期高达516、737株·m-2,繁育期减少至100株·m-2左右,成熟期减少至40株·m-2左右。而入侵两年生境的种群密度在整个生育期的变化趋势与其他两个生境密度变化相对较为平缓,从幼苗期的107株·m-2递减到成熟期的16株·m-2。
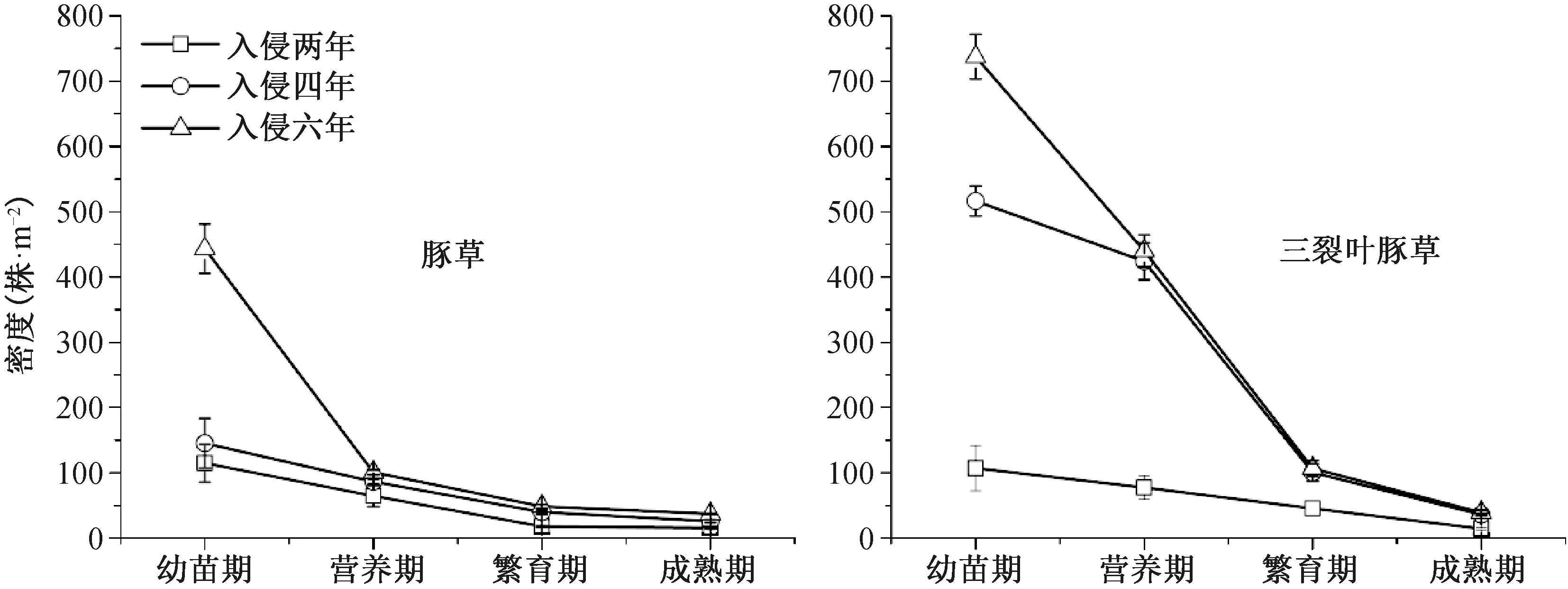
图2 豚草和三裂叶豚草不同生育期种群密度的变化
3 讨论
3.1 土壤种子库特征
研究发现不同生境的土壤种子库大小差异大,三个土层深度(0~10、10~20、20~30 cm)种子密度分别在109~11 868、97~796、4~538粒·m-2,入侵时间越久的生境种子库密度越大,且存在明显的垂直分布。研究表明,豚草产生的种子有15%能在下一年萌发,30%~40%没有发芽的种子能在土壤中存活1年,然后需要40年以上才能完全耗尽[37-38]。随种群定居时间的增长,能够在地下长久的保存这一特性使得二者土壤种子库储量越来越大。
二者种子在土壤种子库中的垂直分布格局基本相似,均是随土层深度的加深而逐渐减小,三个生境中0~10 cm土层中的种子密度最大,是20~30 cm土层种子密度的14~27倍,随土层深度增加,种子数量呈减少趋势且种子类型也在减少。由于二者成熟植株产生的种子进入土壤仅受到自身重力作用,较难下沉到土壤深层,且土壤表层往往堆积枯枝落叶,具有捕获“种子”的作用[39-41],只有受到外力作用时种子才能到达一定土层深度,而二者种子在春季的萌发数量较大,因此只有较少部分的种子能够到达土壤深层。但这部分种子能长期保持活力,对于保存自身种群的基因库、维持种群延续和重建具有重要作用。
3.2 土壤种子库种子类型对种群密度的影响
土壤种子库代表着植被的过去记忆和种群未来的结构[42]。对于种群的更新来说,种子库储量和质量尤为重要[43-44],也能在不稳定环境中稳定种群[45]。
本文研究结果表明,入侵六年的豚草和三裂叶豚草种群密度最高,相应的种子库密度也高,说明种子库密度与种群植被覆盖密度有关,张建利等[33](2008)研究发现扭黄茅(Heteropogoncontortus)、升马唐(Digitariaciliaris)、丝叶球柱草(Bulbostylisdensa)等物种种群的土壤种子库与植被密度呈极显著正相关。不同生境的豚草和三裂叶豚草种群密度在幼苗期最高,幼苗期种子的大量萌发消耗了种子库的存储,随着生育期的变化种群密度下降,成熟期新产生的种子又将补充土壤种子库。
豚草和三裂叶豚草结实量巨大,又以近距离传播为主,因此有着庞大的土壤种子库。本研究发现,豚草和三裂叶豚草土壤种子库中的虫食种子、干瘪种子和霉变种子占了相当一部分比例。其中,豚草入侵六年生境的土壤种子库中这三种类型种子约25%。三裂叶豚草入侵二、四、六年生境的土壤种子库这三种类型种子占比分别达到了23%、34%、15%。这些种子在后续的生育期中基本不可萌发,这大大降低了两物种种群数量过剩的问题。
豚草和三裂叶豚草的种子进入土壤种子库后,种子的干瘪、虫食、霉变等不同类型的损失构成了土壤种子库动态变化的各种要素,这些要素将影响到种子的萌发格局,成为决定种群密度的因素之一。
3.3 土壤种子库时空变化对防控的指示意义
从研究结果来看,豚草和三裂叶豚草土壤种子库中的上年种子对地上种群密度的贡献最大。采用化学药剂处理[46-47]或苗期至开花结实之前及时刈割[5、48]等方法,杜绝或降低其种子的产生以及向地下土壤种子库的输入,可能会有效地降低豚草和三裂叶豚草种群数量,从而大幅度降低其入侵危害。需要注意的是,植株割除后的残留部分往往会从根部或留下的叶腋处生成新的植株,所以割除时期的选择以及注意贴地割除、不留高茬非常重要,否则其新生株仍会开花结实,导致防控效果较差[49]。
通过种群密度动态变化可以发现,两个物种均有种群密度大幅降低阶段,豚草为幼苗期至营养期阶段,三裂叶豚草为幼苗期至繁育期阶段。此阶段其种群有着激烈的种内竞争,因此,可以采用诱萌等方式,提高萌发期种群整体的萌发率,使种群密度超过阈值,增强同胞竞争,从而控制种群增长。一方面消耗种子库的种子量,一方面减少结实量及对种子库的输入。
随着时间延长,两个物种土壤种子库的消耗逐渐增大。对于入侵时间较久的生境,可以通过物理手段将0~10 cm种子较多的土层进行深翻,使种子不能正常萌发,压制种子萌发数量,促进种子霉变、腐烂等,增加土壤种子库的自然消耗。化学手段方面则可以通过药剂或化感物质的浇灌抑制或延迟种子的萌发,也是防控豚草和三裂叶豚草的一个辅助手段。由于深层土壤种子库的存在,如果采用刈割等物理防控或化学防控后,需要后期连续多年的观测。
