双叉犀金龟表皮蛋白TdCPR12611与TdCPR7854的表达纯化及特性分析
2021-03-09叶长青
叶长青,包 涵,刘 田,杨 青
(大连理工大学生物工程学院,辽宁大连 116024)
仿生材料是材料学的重要研究方向,生物体各具特色的表面结构对于仿生材料的设计具有重要的借鉴意义。其中昆虫表皮凭借高强度、轻量型的特性,一直是仿生材料研究的热点。陈斌等(2000)通过研究蝉的表皮结构,设计出具有高强度高韧性的复合材料;Vincent和Wegst (2004)对昆虫表皮的硬度强度进行研究,提出了仿昆虫表皮的纤维复合材料设计方案。仿生表皮的研发应用为人造材料的性能优化提供重要的启示和指导。
昆虫表皮(外骨骼)对于其维持运动能力、对抗外部环境压力、抵御机械损伤以及免疫病原微生物侵害等方面具有至关重要的作用(Vincent and Wegst,2004;Websteretal.,2017;Deshouxetal.,2018)。昆虫表皮是由上皮细胞分泌沉积而来,它覆盖虫体表面的组织器官,维持虫体的完整性(梁欣等,2014),主要由几丁质以及多种类型的表皮蛋白组成。几丁质是N-乙酰氨基葡糖聚合物,可以分为晶态几丁质、非晶态几丁质和脱乙酰几丁质(壳聚糖)等多种形式(Fukamizo and Kramer,1985)。晶态几丁质根据其糖链排列方式的差异又可分为α-几丁质(反平行)和β-几丁质(平行)等,非晶态几丁质是破坏晶体结构的胶体几丁质;晶态和非晶态几丁质都不溶于水。昆虫表皮蛋白种类繁多,目前主要被分为12个家族(Jasrapuriaetal.,2012;Liuetal.,2015;Vanninietal.,2015;Songetal.,2016;Cinegeetal.,2017;Liuetal.,2018),其中CPR(cuticular proteins that contain the R&R consensus)蛋白是所有表皮蛋白中含量最为丰富的家族(Liuetal.,2018)。CPR家族成员都含有较为保守的R&R基序,被认为与几丁质结合相关。根据R&R基序的差异,CPR家族又被分为3个亚族,分别为RR-1(主要分布于柔软表皮中)、RR-2(主要分布于坚硬表皮中)以及RR-3(含量较少)(Andersen,1998)。已有研究表明,CPR家族表皮蛋白对于维持表皮完整性以及刚性韧性等方面具有关键作用(Arakaneetal.,2012;Zhaoetal.,2019)。另外,种类丰富的CPR家族成员与不同类型的几丁质具有相互作用(Rebers and Willis,2001;Togawaetal.,2004;Tangetal.,2010)。
双叉犀金龟Trypoxylusdichotomu的头部角突是由其表皮特化而来,它是一种天然的防御攻击武器,不仅能够产生强力的机械攻击,也能够将自身以及大脑承受的伤害降至最低,具有显著的抗断裂韧性和抗形变刚性。Lee等(2017)利用聚焦离子束原位操作和测试的方法对双叉犀金龟头部角突的机械性能进行研究,结果表明其头部角突具有很高的弹性模量及抗断裂拉伸强度。Zhang等(2019)利用扫描电镜观察双叉犀金龟的头部角突发现其角质层是由高度有序的几丁质纳米纤维束排布构成,纤维束能够排列成环向层,层与层之间通过纤维涂层相互连接,相邻层之间旋转堆叠形成典型的胶合板结构,这种构造为头突角突的刚性和韧性提供支撑;X射线能量色散光谱(energy dispersive X-ray spectrum,EDS)表明角中的蛋白由内向外逐渐富集并呈梯度变化,且几丁质纳米纤维的取向也逐渐由环向往径向改变,蛋白阶梯式变化与几丁质纳米纤维的重新定向进一步增强了角的刚韧程度;此外,拉曼光谱表明角的主要成分是几丁质和蛋白质。
因此,本研究选择两个存在于双叉犀金龟头部角突中的表皮蛋白作为研究对象,首先克隆了编码两个表皮蛋白TdCPR12611和TdCPR7854的基因;然后通过原核表达及金属离子亲和层析纯化的方法获得了两个目的蛋白,并对其序列特征进行生物信息学分析;对两个目的蛋白与不同类型几丁质结合能力的进行了检测;观察比较两个目的蛋白形成液液相分离性质的差异;结合生物信息学分析结果对两个目的蛋白性质差异的原因进行了探讨,旨在通过研究加深对昆虫表皮形成机制的了解,为开发优质的仿生表皮材料提供思路。
1 材料与方法
1.1 材料与试剂
大肠杆菌Escherichiacoli基因克隆菌株DH5α和蛋白表达菌株BL21(DE3)、PrimeSTAR® HS (Premix)高保真酶、Premix TaqTM(Ex TaqTMVersion 2.0)、Mighty TA-cloning Kit、RNAiso Plus RNA提取试剂盒、PrimeScriptTMRT Reagent Kit反转录试剂盒、限制性核酸内切酶NdeⅠ和XhoⅠ以及In-Fusion连接酶购自TaKaRa公司;pET28a质粒载体为实验室保存;其他未标明试剂均为国产分析纯。
1.2 表皮蛋白基因克隆和测序验证
基于实验室早期通过对双叉犀金龟蛹末期的头部角突组织蛋白组与转录组测序数据,将获得的氨基酸及核酸序列信息通过Blast及CutProtFam-Pred进行序列比对并分类,筛选出两个属于CPR_RR-2家族的表皮蛋白TdCPR12611和TdCPR7854。利用RNAiso Plus试剂盒提取双叉犀金龟预蛹期头壳组织总RNA,利用PrimeScriptTMRT Reagent Kit反转录试剂盒合成cDNA,引物序列见表1,PCR反应体系(50 μL):cDNA模板1 μL,Premix TaqTM(2×) 25 μL,T-12611/7854-F和T-12611/7854-R(10 μmol/L)各1 μL,ddH2O 22 μL。反应条件:98℃ 10 s,55℃ 30 s,72℃ 1 min,30个循环;4℃保存。将PCR扩增产物连接到克隆载体T-vector上,通过测序验证扩增片段的正确性。通过信号肽预测确定信号肽编码序列的分割点位置,以T-vector为模板,根据信号肽之后的片段设计连接引物(表1),同时在序列两端引入20 bp左右的与表达载体序列一致的同源区域,并在两端引入限制性内切酶的酶切位点,在预测蛋白序列N末端加入6×His Tag,PCR反应体系(50 μL):T-vector模板1 μL,PrimeSTAR HS (2×) 25 μL,pET28a-12611/7854-F和pET28a-12611/7854-R (10 μmol/L) 1 μL。反应条件:98℃ 10 s,55℃ 5 s,72℃ 5 s,35个循环;4℃保存。利用限制性内切酶NdeⅠ和XhoⅠ对表达载体质粒pET28a进行双酶切,通过In-Fusion同源重组的方法将携带目的基因的片段与切割好的载体连接成完整的表达质粒pET28a-TdCPR12611及pET28a-TdCPR7854,并进行测序验证。

表1 引物信息Table 1 Primer information
1.3 TdCPR12611及TdCPR7854蛋白的结构特征及系统发育分析
利用表皮蛋白序列在线分析软件CutProtFam-Pred(http:∥aias.biol.uoa.gr/CutProtFam-Pred/search.php)对双叉犀金龟表皮蛋白进行分类表征;利用 SignalP-5.0 Server(http:∥www.cbs.dtu.dk/services/SignalP/)进行信号肽预测;利用 ExPASy 数据库(https:∥web.expasy.org/protscale/)进行亲疏水性预测分析;利用PLAAC (http:∥plaac.wi.mit.edu/)进行蛋白的内部无序性(intrinsic disorder)预测;利用CIDER (http:∥pappulab.wustl.edu/CIDER/)进行蛋白带电荷数分析,并对蛋白的每个残基的净电荷数(net charge per residue,NCFR)和带电残基分数(fraction of charged residues,FCR)进行计算。利用BLAST(https:∥blast.ncbi.nlm.nih.gov/Blast.cgi)在线搜索工具将TdCPR12611和TdCPR7854氨基酸序列与GenBank数据库中的蛋白的序列进行比对,筛选出与之序列一致性最高的蛋白;利用MEGA7软件采用邻接(neighbor-joining)法对47种昆虫表皮蛋白进行系统进化树分析(500次重复)。
1.4 TdCPR12611及TdCPR7854的原核表达及分离纯化
将测序验证无误后的表达质粒pET28a-TdCPR12611及pET28a-TdCPR7854转化入大肠杆菌表达菌株BL21(DE3),菌株活化后扩大培养到对数生长期OD600=0.5~0.6,加入终浓度为0.5 mmol/L的IPTG,37℃诱导5 h后离心收集细菌。加缓冲液(20 mmol/L Tris,500 mmol/L NaCl,pH 7.4)重悬后,利用高压匀浆破碎仪破碎细菌,离心去除细胞碎片沉淀,上清过镍离子亲和层析柱;用AKTA蛋白纯化仪对表达蛋白TdCPR12611进行洗涤(20 mmol/L Tris,500 mmol/L NaCl,100 mmol/L咪唑,pH 7.4)和洗脱(20 mmol/L Tris,500 mmol/L NaCl,200 mmol/L咪唑,pH 7.4),对表达蛋白TdCPR7854蛋白进行洗涤(20 mmol/L Tris,500 mmol/L NaCl,100 mmol/L咪唑,pH 7.4)和洗脱(20 mmol/L Tris,500 mmol/L NaCl,250 mmol/L咪唑,pH 7.4);最后用SDS-PAGE检测蛋白纯化的结果,并用Western blot进行验证。
1.5 TdCPR12611及TdCPR7854蛋白与几丁质结合实验
采用将蛋白与不同类型的几丁质混合结合并洗脱的方法(张薇薇等,2017),选择α-几丁质(α-chitin)、β-几丁质(β-chitin)、壳聚糖(chitosan)和胶体几丁质(colloidal chitin)4种类型的几丁质与重组表达的表皮蛋白进行体外结合实验。具体实验操作如下:将纯化后的重组表达蛋白透析到结合缓冲液(20 mmol/L Tris,500 mmol/L NaCl,100 mmol/L咪唑,pH 7.4)中。构建200 μL反应体系,使蛋白终浓度为0.5 mg/mL,几丁质终浓度为2 mg/mL。反应在室温(25℃)进行,两种蛋白与不同类型几丁质在2 mL离心管中持续颠倒混匀2 h。反应结束后,12 000 r/min离心10 min,将上清取出记为未结合几丁质的蛋白。在沉淀中加入相当于反应体系5倍体积的漂洗缓冲液(20 mmol/L Tris,500 mmol/L NaCl,100 mmol/L咪唑,pH 7.4),将沉淀重悬并颠倒混匀5 min进行漂洗后12 000 r/min离心5 min弃上清,重复漂洗3次。最后将沉淀放入烘箱中彻底干燥,加入200 μL的结合缓冲液后对沉淀进行重悬,取同样体积的上清和沉淀重悬液进行SDS-PAGE,检测蛋白与不同类型几丁质的结合情况。利用Image J灰度分析软件将SDS-PAGE结果进行灰度扫描量化,将用于结合实验的蛋白条带灰度值作为背景,进行反应体系上清液及沉淀重悬液蛋白条带相对灰度计算,用相对灰度值代表结合几丁质的蛋白占总蛋白的百分比,以此表征蛋白与几丁质的结合能力。利用SPSS 22.0对结合几丁质蛋白条带的灰度数据进行单因素方差分析及差异显著性分析(LSD法)。
1.6 TdCPR12611及TdCPR7854蛋白液液的相分离性质观察
在对两蛋白分离纯化的过程中,发现TdCPR12611蛋白在浓度约为15 mg/mL,缓冲液为20 mmol/L Tris,500 mmol/L NaCl,pH 7.4条件下,存在温度响应的液液相分离现象,而在相同条件下TdCPR7854不存在类似现象。将TdCPR12611蛋白溶液置于比色皿中,用肉眼观察在4℃和25℃下蛋白溶液产生液液相分离的宏观情况。用光学显微镜观察400倍(目镜10×,物镜40×)放大倍数条件下TdCPR12611蛋白溶液形成相分离的微观情况。
2 结果
2.1 TdCPR12611及TdCPR7854蛋白的结构特征
克隆获得双叉犀金龟表皮蛋白基因TdCPR12611(GenBank登录号:MT813021)和TdCPR7854(GenBank登录号:MT813022)序列。通过对TdCPR12611和TdCPR7854蛋白一级结构分析发现这两个蛋白具有许多相似点,它们均属于表皮蛋白CPR家族中RR-2亚族成员,其N端存在一段信号肽,为外泌型蛋白,且都存在一段与几丁质结合相关的结构域R&R基序(图1)。但两者也存在许多差异,TdCPR12611存在两段富含谷氨酰胺的区域(Q-rich region),蛋白中Q的含量很高,为16.8%(52/309);而TdCPR7854则存在两段富含甘氨酸的区域(G-rich region),蛋白中G的含量很高,为28.7%(52/181)。分析蛋白的疏水性发现,TdCPR12611除了信号肽部分比较疏水外,该蛋白整体为亲水性蛋白(图1:A);TdCPR7854的信号肽及第一段G-rich region较为疏水,其余部分也为亲水性结构(图1:B)。
分析蛋白的内部无序性发现,TdCPR12611蛋白的信号肽部分以及R&R motif部分比较有序,而两段Q-rich region比较无序,相对无序的部分可能对形成液液相分离做出贡献,暗示该蛋白具有形成功能性结构的潜力。分析其每个氨基酸残基的带电荷数和带电残基分数可得,两段Q-rich region几乎不带电荷,R&R motif为正负电荷交替出现(图2:A);而TdCPR7854蛋白整体都是较为有序的状态,且该蛋白整体为正负电荷交替出现状态(图2:B)。推测静电相互作用可能是导致不同片段结构无序性差异的原因之一。
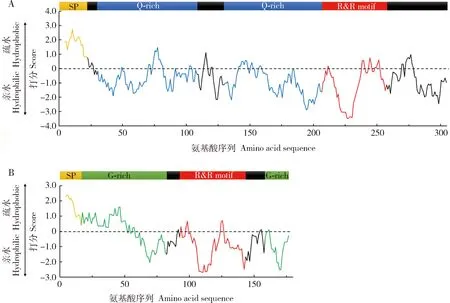
图1 双叉犀金龟表皮蛋白TdCPR12611 (A)及TdCPR7854 (B)的结构组成及疏水性分析Fig.1 Structural composition and hydrophobicity analysis of the cuticular proteins TdCPR12611 (A) and TdCPR7854 (B) of Trypoxylus dichotomusSP:信号肽Signal peptide;Q-rich:富含谷氨酰胺的区域Glutamine-rich region;R&R motif:与几丁质结合相关的R&R基序R&R domain associated with chitin-binding;G-rich:富含甘氨酸的区域Glycine-rich region.图2同The same for Fig.2.

图2 双叉犀金龟表皮蛋白TdCPR12611(A)及TdCPR7854 (B)内部无序性分析Fig.2 Intrinsic disorder analysis of the cuticular proteins TdCPR12611 (A) and TdCPR7854 (B) of Trypoxylus dichotomusPLD:内部无序性预测值在0.5以上符合朊蛋白样结构,为无序的结构,红线为预测线,黑线为背景线 Prediction of internal disorder,the predicted value above 0.5 conforms to prion protein-like structure,which is the disordered structure.The red line is the prediction line,and the black line is the background line.NCPR:每个氨基酸残基的带电荷数分析,其中蓝色为正电荷,红色为负电荷 Net charge per residue,in which blue is positive charge and red is negative charge.FCR:带电残基分数,带电的氨基酸残基对于蛋白带电性的贡献比例Charged residue fraction,the percentage of charged amino acid residues contributing to total protein charge.
2.2 TdCPR12611及TdCPR7854的系统发育分析
通过将TdCPR12611(GenBank登录号:MT813021)和TdCPR7854(GenBank登录号:MT813022)编码的氨基酸进行BLAST序列比对,结果表明:TdCPR12611蛋白与食粪金龟Onthophagustauruscuticular protein 1(GenBank登录号:XP_022920963.1)、光肩星天牛Anoplophoraglabripenniscuticular protein 1(GenBank登录号:XP_018577010.1)、北美萤火虫Photinuspyraliscuticular protein ts15(GenBank登录号:XP_031347513.1)及边褐端黑萤Absconditaterminaliscuticular protein hp65(>KAF5297863.1)的序列一致性较高,分别为83%,78%,75%和75%;TdCPR7854与食粪金龟adult-specific cuticular protein ACP-20-like(GenBank登录号:XP_022914362.1)和中欧山松大小蠹Dendroctonusponderosaeadult-specific cuticular protein ACP-20-like(GenBank登录号:XP_019761647.1)的序列一致性相对较高,分别为46%和40%。将47种昆虫表皮蛋白进行系统进化树分析发现:TdCPR12611与食粪金龟RR-2家族蛋白OtCP-1(GenBank登录号:XP_022920963.1)的亲缘关系最近,在同一分支上;TdCPR7854与食粪金龟3种不同类型的adult-specific cuticular protein ACP-20-like蛋白OtCP-acp20-1(GenBank登录号:XP_022914362.1)、OtCP-acp20-2(GenBank登录号:XP_022920192.1)和OtCP-acp20-3(GenBank登录号:XP_022903303.1)的亲缘关系最为接近,它们均属于CPR_RR-2家族(图3)。
2.3 双叉犀金龟表皮蛋白TdCPR12611及TdCPR7854的表达及纯化
在带有TdCPR12611和TdCPR7854基因的克隆载体和表达载体测序鉴定无误后,对两个蛋白进行了原核细胞的诱导表达,经过镍离子亲和层析纯化后获得的目的蛋白,SDS-PAGE和Western blot检测结果如图4所示。TdCPR12611蛋白分子量约35 kD(理论分子量32.52 kD),TdCPR7854蛋白分子量约18 kD(理论分子量17.62 kD),从SDS-PAGE结果可以看出蛋白的纯度较高,可以作为后续实验的材料。Western blot结果均为单一信号条带,说明蛋白确实是重组表达的蛋白,并且蛋白较为稳定没有降解的现象。
2.4 TdCPR12611和TdCPR7854蛋白与几丁质结合能力
利用SDS-PAGE检测TdCPR12611及TdCPR7854与不同类型几丁质的结合能力情况,灰度量化的结果表明2种蛋白与4种不同类型几丁质的结合能力均存在显著差异。TdCPR12611仅有蛋白能够与α-几丁质结合,蛋白主要存在于上清(未结合的蛋白)中;与β-几丁质和胶体几丁质的结合能力相近,约有28.0%的TdCPR12611蛋白结合,在沉淀(结合的蛋白)中存在部分蛋白;与壳聚糖的结合能力相对较强,41.4%的TdCPR12611蛋白结合(图5:A,B)。TdCPR7854蛋白几乎不与α-几丁质结合,约有30.0%的蛋白能够与壳聚糖和胶体几丁质结合,与β-几丁质存在较强结合能力,约有62.3%的蛋白结合(图5:C,D)。
2.5 TdCPR12611及TdCPR7854蛋白的液液相分离性质
肉眼观察两个蛋白溶液发现TdCPR12611蛋白在4℃条件下是清澈透明的状态,随着温度升高到室温(25℃),蛋白溶液逐渐由澄清变浑浊;而TdCPR7854蛋白未出现类似现象。在室温下用光学显微镜观察两者的蛋白溶液,发现TdCPR12611蛋白出现团聚体(coacervate)现象,团聚体具有流动形变性,并能够相互融合形成更大的团聚体;而且团聚体形成与消失是可逆的,温度升高至室温团聚体形成,当温度降低到4℃条件时团聚体会消失,蛋白溶液恢复澄清状态(图6);而TdCPR7854蛋白则不具有这种性质。
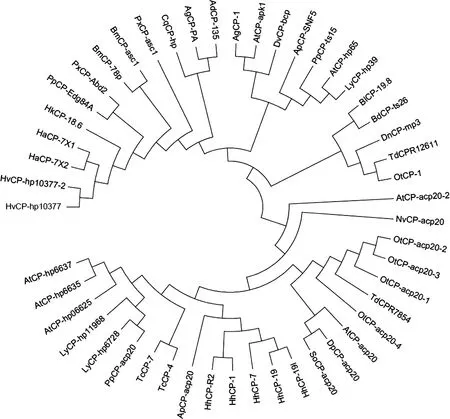
图3 邻接法构建的基于氨基酸序列的昆虫表皮蛋白系统进化树(500次重复)Fig.3 Phylogenetic tree of insect cuticular proteins by neighbor-joining method based on amino acid sequences (500 replicates)蛋白来源物种及其GenBank登录号Origin species of proteins and their GenBank accession numbers:TdCPR12611:双叉犀金龟Trypoxylus dichotomus,MT813021;TdCPR7854:双叉犀金龟Trypoxylus dichotomus,MT813022;OtCP-1:食粪金龟Onthophagus taurus,XP_022920963.1;AgCP-1:光肩星天牛Anoplophora glabripennis,XP_018577010.1;PpCP-ts15:北美萤火虫Photinus pyralis,XP_031347513.1;AtCP-hp65:边褐端黑萤Abscondita terminalis,KAF5297863.1;OtCP-acp20-1:食粪金龟Onthophagus taurus,XP_022914362.1;OtCP-acp20-2:食粪金龟Onthophagus taurus,XP_022920192.1;OtCP-acp20-3:食粪金龟Onthophagus taurus,XP_022903303.1;LyCP-hp39:扁萤Lamprigera yunnana,KAF5299004.1;HkCP-18.6:海波斯莫科马属蛾Hyposmocoma kahamanoa,XP_026328431.1;AgCP-PA:冈比亚按蚊Anopheles gambiae,XP_556966.3;AdCP-135:达氏按蚊 Anopheles darling,ETN63120.1;PxCP-asc1:小菜蛾 Plutella xylostella,XP_011550962.1;ApCP-SNF5:白蜡窄吉丁Agrilus planipennis,XP_018318748.1;AtCP-apk1:小蜂窝甲虫 Aethina tumida,XP_019877778.1;CqCP-hp:致倦库蚊Culex quinquefasciatus,XP_001865100.1;DnCP-mp3:麦双尾蚜Diuraphis noxia,XP_015375949.1;DvCP-bcp:玉米根虫Diabrotica virgifera,XP_028141552.1;BlCP-19.8:马来西亚实蝇Bactrocera latifrons,XP_018801521.1;HvCP-hp10377:烟芽夜蛾Heliothis virescens,PCG64628.1;HaCP-7X2:棉铃虫Helicoverpa armigera,XP_021196834.1;HvCP-hp10377-2:烟芽夜蛾Heliothis virescens,PCG64627.1;HaCP-7X1:棉铃虫Helicoverpa armigera,XP_021196833.1;BdCP-ts26:橘小实蝇 Bactrocera dorsalis,XP_011200150.1;BmCP-78p:家蚕Bombyx mori,NP_001166682.1;BmCP-asc1:野桑蚕Bombyx mandarina,XP_028026464.1;PpCP-Edg84A:玉带凤蝶Papilio polytes,XP_013134319.1;PxCP-Abd2:柑橘凤蝶Papilio xuthus,NP_001298788.1;DpCP-acp20:中欧山松大小蠹Dendroctonus ponderosae,XP_019761647.1;LyCP-hp11968:扁萤Lamprigera yunnana,KAF5297637.1;TcCP-4:赤拟谷盗 Tribolium castaneum,EFA04643.2;TcCP-7:赤拟谷盗Tribolium castaneum,XP_008193006.2;AtCP-hp06625:边褐端黑萤 Abscondita terminalis,KAF5307754.1;HhCP-1:茶翅蝽 Halyomorpha halys,KAE8573268.1;PpCP-acp20:北美萤火虫Photinus pyralis,XP_031344218.1;LyCP-hp6728:扁萤 Lamprigera yunnana,KAF5275617.1;SoCP-acp20:米象Sitophilus oryzae,XP_030749553.1;AtCP-acp20:小蜂窝甲虫Aethina tumida,XP_019875905.1;NvCP-acp20:大红葬甲Nicrophorus vespilloides,XP_017784612.1;AtCP-acp20-2:小蜂窝甲虫Aethina tumida,XP_019875906.1;HhCP-19l:茶翅蝽Halyomorpha halys,XP_014279712.1;OtCP-acp20-4:食粪金龟Onthophagus taurus,XP_022903303.1;HhCP-19:茶翅蝽 Halyomorpha halys,XP_014279718.1;ApCP-acp20:白蜡窄吉丁Agrilus planipennis,XP_025830566.1;AtCP-hp6635:边褐端黑萤Abscondita terminalis,KAF5307764.1;HhCP-7:茶翅蝽Halyomorpha halys,XP_024217407.1;HhCP-R2:茶翅蝽 Halyomorpha halys,KAE8573269.1;AtCP-hp6637:边褐端黑萤Abscondita terminalis,KAF5307766.1.

图4 纯化的双叉犀金龟重组表皮蛋白TdCPR12611 (A)和TdCPR7854 (B)的SDS-PAGE及Western blot检测Fig.4 SDS-PAGE and Western blot detection of the purified recombinant cuticular proteins TdCPR12611 (A) and TdCPR7854 (B) of Trypoxylus dichotomusM:蛋白低分子量标准Low molecular weight protein marker;WB:Western blot.

图5 双叉犀金龟表皮蛋白TdCPR12611(A,B)和TdCPR7854 (C,D)与不同类型几丁质的结合能力Fig.5 Binding abilities of the cuticular proteins TdCPR12611 (A,B) and TdCPR7854 (C,D) of Trypoxylus dichotomus with different types of chitinA,C:SDS-PAGE;B,D:灰度扫描量化结果 Gray scan quantization result.T:用于结合实验的表皮蛋白样品 Cuticular protein sample for binding experiment;F:反应体系上清液,即未结合几丁质的蛋白Supernatant of the reaction system,unbound protein with chitin;E:反应体系沉淀重悬液,即结合几丁质的蛋白Sediment resuspension of the reaction system,bound protein with chitin;M:蛋白低分子量标准Protein low molecular weight marker.图中灰色柱表示结合几丁质的蛋白占总蛋白的百分比,每个处理3个技术重复。柱上不同小写字母表示与不同类型几丁质结合的蛋白的百分比(平均值±标准误)差异显著(P<0.05,LSD法)。The gray column in the figure indicates the percentage of protein binding with chitin,and each treatment has three technical replicates.Different lowercase letters above bars represent significant differences in the percentage (mean±SE) of protein binding with different types of chitin (P<0.05,LSD method).

图6 双叉犀金龟表皮蛋白TdCPR12611液液相分离现象观察Fig.6 Observation of liquid-liquid phase separation of cuticular protein TdCPR12611 of Trypoxylus dichotomusA:TdCPR12611蛋白溶液在不同温度下的肉眼观察结果,蛋白出现液液相分离现象随着温度改变是可逆的The macroscopic observation of TdCPR12611 protein solution at different temperatures showed that the protein separation in liquid phase was reversible with the change of temperature;B:TdCPR12611蛋白在室温条件下光学显微镜观察结果,蛋白溶液中出现的圆球即为团聚体TdCPR12611 protein was observed under light microscope at room temperature,and the globules in the protein solution were coacervates.
3 讨论
本研究选择了双叉犀金龟头部角突组织中表达含量较高的两种表皮蛋白TdCPR12611及TdCPR7854作为研究对象。生物信息学分析发现它们均属于昆虫表皮蛋白CPR_RR-2家族成员,携带有保守的R&R结构域,这说明两者可能与几丁质具有相互作用。系统进化树分析表明两者与来源于食粪金龟中不同的CPR_RR-2家族表皮蛋白亲缘关系较近。与几丁质体外结合实验表明,两种蛋白与不同类型的几丁质结合具有显著的差异,其中TdCPR12611与壳聚糖具有较强的结合能力,TdCPR7854与β-几丁质的结合能力较强,这说明两蛋白在双叉犀金龟头部角突的形成过程中可能发挥重要作用,能够帮助双叉犀金龟头部角突形成坚硬的组织结构。这进一步说明RR-2家族表皮蛋白在昆虫表皮形成装配过程中发挥重要作用。同时,由于RR-2家族蛋白多存在于昆虫坚硬组织中(刘晓健等,2019),因此推测两蛋白具有作为优良蛋白质仿生材料的潜力。已有研究证明蛋白质的正确折叠对几丁质材料的刚度有很大影响,几丁质结合域(chitin-binding domain,CBD)的存在极大地影响了材料的最大应力和杨氏模量。例如天然材料鱿鱼的软骨中去除蛋白而保留几丁质骨架会造成软骨部分的最大应力损失>80%,杨氏模量损失>90%(Montronietal.,2021)。含有两个纤维素结合模块(cellulose-binding modules,CBM)的工程蛋白与纳米纤维素复合制膜,也能够显著提升纳米纤维素膜的极限拉伸强度、刚度及韧性(Malhoetal.,2015)。TdCPR12611蛋白具有能够与几丁质结合的结构模块,实验表明该蛋白与壳聚糖具有较强的相互作用。因此,推测TdCPR12611蛋白可能能够与几丁质复合,形成具备优良性能的仿生材料。
实验过程中发现TdCPR12611蛋白能够在室温条件下自发团聚形成LLPS,与Mohammadi等(2018)利用蛛丝蛋白ADF3形成液体状的团聚体(liquid-like coacervate,LLC)的性质类似,而TdCPR7854则不具备类似的性质。在研究弹性蛋白(elastin)和节肢弹性蛋白(resilin)温度响应相变过程中发现,它们普遍存在一种P-Xn-G基序(P:脯氨酸;G:甘氨酸;X:任意一种氨基酸;n为0~4整数),作者认为这种基序是组成蛋白自身无序结构的框架(Quiroz and Chilkoti,2015)。而在TdCPR12611蛋白的序列中不存在类似的基序,推测是不带电的极性氨基酸Q含量高,分子内缺乏静电相互作用稳固蛋白结构,使占主导地位的两段Q-rich region为低复杂度序列,从而使整个蛋白呈现内部无序状态。而蛋白内部的无序性,可以对形成液液相分离的现象做贡献(Albertietal.,2019),这可能是造成TdCPR12611蛋白能够形成LLPS的原因。团聚是指将均质大分子溶液分离成两种不同的液相。在水溶液中团聚体被认为有利于组装生物大分子网络结构,原因是团聚体具有很高的局部蛋白质浓度,但它仍呈液体状且不溶于水,这有利于在细胞外环境中高浓度地运输蛋白质。团聚体在低浓度下具有较低的粘度以及极低的界面自由能,并且具有剪切稀释特征,使其能够浸渍到纳米多孔骨架(如几丁质-几丁质结合蛋白骨架结构)之中。已有研究表明,许多蛋白在形成具有功能性的结构之前都需要经历团聚体的过渡形式(Mohammadietal.,2018),如在研究贻贝类等海洋生物的粘附剂领域,足丝蛋白首先经历液液相分离然后迅速固化达到粘附于固体表面的效果(Stewartetal.,2011;Waite,2017);鱿鱼喙在形成过程中,一种富含组氨酸的recDgHBP-1蛋白也需要经历形成团聚体这一中间环节(Tanetal.,2015);在弹性蛋白组装成具有功能性结构的过程中也涉及蛋白团聚步骤(Muiznieksetal.,2014)。TdCPR12611蛋白在纯化后也存在形成团聚体的现象,由此进一步推测该蛋白具有组装形成优良性能结构材料的潜力。
综上所述,本研究比较了两种来源于双叉犀金龟头部角突中的表皮蛋白,两者均属于CPR家族中的RR-2亚族成员,它们均具有良好的亲水性,但它们在内部无序性、形成液液相分离的性质及几丁质结合能力上均具有明显的差异,其中TdCPR12611蛋白具备优良的性能及应用于仿生材料制备的潜力。本研究进一步表明了表皮蛋白CPR家族在昆虫表皮结构中承担着重要角色,对研究昆虫表皮的形成过程有所启发,并且为制备新型蛋白质仿生材料的原料选择方面提供了新依据。
