海南岛热带低地雨林2种攀缘竹的叶片功能性状差异*
2021-02-12徐瑞晶刘广路梁昌强孔祥河
徐瑞晶 胡 璇 刘广路 郭 雯 梁昌强 孔祥河
(1.国际竹藤中心 北京 100102; 2.海南省甘什岭自然保护区管理站 三亚 572000)
植物为适应不同生境,在叶片光合特性、解剖结构和形态性状上表现出不同的适应策略(张咏梅等, 2019; 寇萌等, 2019; 钟巧连等, 2018)。叶片是植物进行光合作用、呼吸作用等生理代谢活动的重要器官,其变化特征最能反映植物的环境适应性(张咏梅等, 2019; Gaoetal., 2017; 王勋陵等, 1989)。热带季雨林植物通过调节叶片净光合速率,积累更多干物质适应雾凉季低温环境(栗忠飞等, 2012),常绿植物通过调节气孔状态,提高光合水分利用效率维持叶片四季常绿(白坤栋等, 2010)。在逆境胁迫中的植物通过调节叶片水势、蒸腾速率和水分利用效率等光合生理因子,以适应季节更替和胁迫(邓彭艳等, 2010; Inoueetal., 2017)。单子叶植物通过增大叶的表皮和上角质层厚度等保护组织厚度来保护叶肉组织,同时依靠泡状细胞和较大的维管束等特殊解剖结构适应水分条件差、强光照的环境(寇萌等, 2019)。气孔密度体现植物对环境(强光、高温、大风等)的响应,在高光照强度或低水分利用率时叶片的气孔密度较高(Rossattoetal., 2012); 发达的维管组织不仅利于提高水分利用效率,同时也是对叶片干旱和强风环境的适应(史刚荣等, 2007)。如内蒙古高原草地植物依靠小而密的气孔,可快速有效地利用水分; 而青藏高原草地植物气孔大而疏,将更多能量用于抵抗低温胁迫(王媛媛等, 2018)。植物在长期适应环境过程中,通过内部不同功能之间的协同进化,形成能适应环境变化的形态和功能,叶功能性状是反映这种适应策略的有效指标,如普定喀斯特地区植物有较低的叶面积和比根长度,较高的叶干物质含量和叶组织密度,这是其适应干旱贫瘠的喀斯特环境的重要生态策略(钟巧连等, 2018)。表型可塑性是植物适应异质生境的重要生态对策,表型可塑性高的物种能适应更宽的环境变化范围,能在不利条件下优化利用资源(Narayan, 2012; 施建敏等, 2014; 梁宽等, 2017)。
竹子叶片功能性状及适应性相关研究已有报道,但只涉及直立竹,未涉及藤本状的攀缘竹。直立竹主要集中在勃氏甜龙竹(Dendrocalamusbrandisii)、黄条金刚竹(Pleioblastuskongosanensisf.aureostriatus)、黄竹(D.membranaceus)等引种后的生长指标和抗性指标评价(邱尔发等, 2006; 夏海涛等, 2016; 唐红等, 2019),以及毛竹(Phyllostachysedulis)的可塑性生长(施建敏等, 2014; 曹永慧等, 2016; 刘广路等, 2017),还有慈竹(Bambusaemeiensis)、八月竹(Chimonobambusaszechuanensis)和短锥玉山竹(Yushaniabrevipaniculata)对海拔梯度的适应生长(蒋琴等, 2018)及淡竹(Phyllostachysglauca)在石灰岩生境的适应生长等方面(梁宽等, 2017)。
攀缘竹是竹亚科(Bambusoideae)的一个特殊类群,主要分布于热带、亚热带高温多雨地区,以云南和海南岛最为丰富。攀缘竹植株奇特,竿呈攀援状或斜倚呈悬垂状,类似藤本植物,可作特色园林和立体绿化用竹,具有较高的观赏价值。攀缘竹是海南岛低地雨林的重要组成部分,具有较高的科学研究价值(胡璇等, 2018; 徐瑞晶等, 2020)。无耳藤竹(Dinochloaorenuda)和响子竹(Bonialevigata)是海南岛热带雨林的攀缘竹代表种,是热带低地雨林的重要组分。无耳藤竹喜阳、喜攀抵雨林冠层争夺光资源,能较好适应高光环境,多分布于雨林林缘或林窗阳光充足处; 响子竹较为耐荫,喜在林下攀爬,多分布在雨林林下弱光环境中。目前,有关2个竹种的研究较少,仅涉及分类学、地径与节长分布和光合特性研究(Xia, 1996; 胡璇等, 2018; 徐瑞晶等, 2020)。本文系统研究2种攀缘竹的叶片解剖结构、气体交换参数、单叶面积和叶片干物质含量等,可以揭示攀缘竹在热带雨林的生存与适应策略,为其资源保护和利用提供科学依据。
1 研究地概况
研究地点位于海南甘什岭省级自然保护区(109°40′4″E,18°23′2″N,海拔202 m)。保护区地处三亚市与保亭县南部交界处,属低山丘陵地貌,坡度30°~50°,海拔50~681 m,土壤母质为花岗岩,岩层裸露率10%,土壤含砂量为20%左右。该区属热带海洋季风气候,年降水量1 200~1 800 mm,干湿季分明,雨季为6—11月,其降水占全年90%以上,旱季为12月—翌年5月。年均气温23.4 ℃,最低月平均气温19.2 ℃,最高月平均气温27.1 ℃(Wheat, 2020)。地带性植被为热带低地雨林,乔木树种有铁凌(Hopeareticulata)、琼南柿(Diospyroshowii)、华润楠(Machiluschinensis)和青梅(Vaticamangachapoi)等,灌木有刺轴榈(Licualaspinosa)、海南龙船花(Ixorahainanensis)、山血丹(Ardisialindleyana)等,草本有高秆珍珠茅(Scleriaterrestris)、乌毛蕨(Blechnumorientale)和华山姜(Alpiniaoblongifolia)等,层间植物有百足藤(Pothosrepens)、无耳藤竹和响子竹等(漆良华等, 2014)。
2 研究方法
2.1 试验取样 所调查的无耳藤竹平均地径(2.3±0.2)cm,单节平均长(35.4±0.4)cm,分布密度为(180±51)丛·hm-2,攀爬高度可至18 m; 响子竹平均地径(0.6±0.1)cm,单节平均长(25±0.6)cm,分布密度为(60±12)丛·hm-2,攀爬高度可至6 m。
在试验前,每个竹种选5株长势相近的植株进行挂牌,作为不同月份取样测定的固定植株。分别于2019年6、9、12月和2020年3月选择晴朗天气的8:30—11:30和14:30—17:30期间,测定2种攀缘竹叶的光合气体交换参数,每个竹种每次测量5株长势相近的植株,每株选取枝条顶端第3~5片完全展开的3片健康成熟叶作为待测叶片。在叶片光合气体交换参数测定结束后,在3片测定叶中采集1片叶,剪成1 cm×1 cm大小,分别置于70% FAA固定液中保存,带回实验室进行叶片石蜡切片和叶片表皮制片,用于观察解剖结构。每个竹种采集植株上5个枝条的全部叶片,带回实验室测定叶片功能性状,每次采样5个植株。
2.2 测定方法 1) 叶片光合气体交换参数测定 用红蓝光源模拟自然光,在无耳藤竹(1 100 μmol·m-2s-1)和响子竹(900 μmol·m-2s-1)的光饱和点附近设定光强,每片叶在饱和点光强下诱导15 min后读数5次,测定其叶片净光合速率(Pn)、气孔导度(Cond)、胞间二氧化碳浓度(Ci)和蒸腾速率(Tr),并计算水分利用效率(WUE=净光合速率/蒸腾速率)(张向峰等, 2012)。
2) 叶片解剖结构观测 叶片石蜡切片制作参照李正理(1987)的方法,材料经软化、脱水浸蜡、包埋、切片、番红-固绿染色和中性树胶封片等步骤; 而后在奥斯微PH50-3 M100光学显微镜下观测和拍照,用ImageView图像分析软件测量各组织叶厚(LT)、上角质层厚度(USCT)、乳突厚度(MPT)、上表皮厚度(UET)、下表皮厚度(LET)、泡状细胞数量(NBC)、泡状细胞截面积(SABC)、单个梭形细胞截面积(SAFC)、主脉维管束截面积(SAFVB)、二级维管束直径(DSVB)、二级维管束横截面积(SASVB)、二级维管束间距(DBAVB)共12个指标。每个竹种在每次观测均取30个视野统计指标值。
叶表皮切片制作用过氧化氢-冰醋酸离析法(唐光大等, 2010)。将叶片放入装有1∶1的冰醋酸与30%过氧化氢混合液的消煮瓶,置于60 ℃水浴温箱中27 h,待叶肉组织与上、下表皮开始分离时,把离析后的材料取出移入盛有蒸馏水的容器中,用毛笔分离上、下表皮,将洗净后的叶表皮放入染色皿中,在1%番红50%酒精溶液内染色5 min后用中性树胶封片。切片在奥斯微PH50-3 M100光学显微镜下观测和拍照,用ImageView图像分析软件测量各组织的气孔长度(SL)、气孔宽度(SW)、单个气孔器面积(AS),并计算气孔密度(SD=视野内气孔个数/视野面积)和气孔器面积百分比(AP=单个气孔器面积×气孔密度×100%)(李冬林等, 2019)。每个竹种在每次观测均取30个视野统计指标值。
3) 单叶面积、叶片干物质含量和比叶面积测定 采用Epson perfection V19通过对叶片扫描测定单叶面积。将成熟叶剪去叶柄,浸泡在清水中12 h,用吸水纸吸去叶表水分,称饱和鲜质量,然后105 ℃下杀青、80 ℃烘干至恒质量,称取干质量。干物质含量(LDMC)=干质量(g)/饱和鲜质量(kg); 比叶面积(SLA)=叶面积(m2)/干质量(kg)(郭雯等, 2019)。
4) 叶片指标可塑性指数计算 叶片指标可塑性指数=(某指标的最大值-最小值)/最大值。可塑性指数范围在0~1之间,数值越大表明可塑性越大,说明该指标对环境适应性越强(Valladaresetal.,2000)。
2.3 数据处理 采用SPSS 20.0对叶片光合气体交换参数、解剖结构指标和形态性状进行多重比较(SNK法),竹种间的指标差异显著性分析使用独立样本t检验,使用Excel 2010和Sigma plot 12.5数据处理软件对所测数据进行统计分析和绘图。
3 结果与分析
3.1 叶片光合气体交换参数月份变化 2种攀缘竹在不同月份的光合气体交换参数结果(图1)表明,无耳藤竹的净光合速率、气孔导度、胞间二氧化碳浓度和蒸腾速率在6月最高,12月最低; 水分利用效率则相反; 反映出无耳藤竹的光合作用能力在雨季(6、9月)高于旱季(12、3月)。响子竹净光合速率、气孔导度、胞间二氧化碳浓度和蒸腾速率在9月最高,3月和6月较低; 水分利用效率在3月最高,9月最低。对2个竹种,除12月的胞间二氧化碳浓度外,无耳藤竹的净光合速率、气孔导度、胞间二氧化碳浓度和蒸腾速率在各月均高于响子竹,2竹种各指标参数在6月和3月之间的差异显著(P<0.05)。无耳藤竹的水分利用效率在6月和3月均显著低于响子竹(P<0.05),在9月和12月高于响子竹,但差异不显著(P>0.05)。以上结果反映了无耳藤竹的叶片光合能力强于响子竹,它们的水分利用效率在各月表现不同。
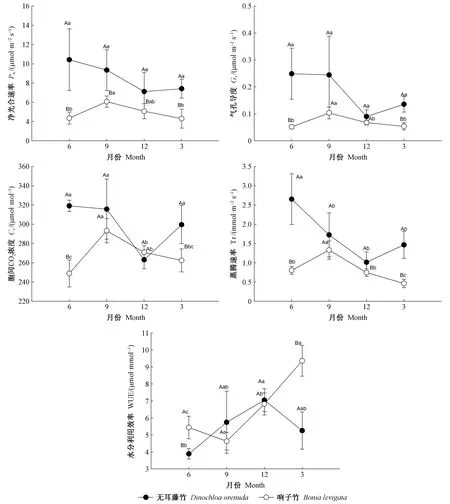
图1 2种攀缘竹叶片的光合气体交换参数月份动态Fig. 1 Monthly dynamics of photosynthetic gas exchange parameters in the leaves of two climbing bamboo species 同月份不同大写字母表示2物种在0.05水平存在显著差异,而同物种不同小写字母表示月份间在0.05水平存在显著差异(P<0.05)。 下同。 Different capital letters in the same month indicate the significant difference between species at 0.05,while different lowercase letters in the same species indicate the significant difference among months at 0.05(P<0.05). The same below.
3.2 叶片解剖结构月份变化 2种攀缘竹的叶片由表皮、叶肉和叶脉3部分组成(图2),它们的解剖结构组成基本相似。竹叶表皮分上表皮和下表皮,均含有1层细胞,上表皮细胞较厚,下表皮细胞较薄; 表皮细胞由长、短2种细胞组成,上表皮有少量气孔分布,下表皮气孔分布较多,气孔的每个保卫细胞外侧各有一个副卫细胞,气孔器分布在脉间区域并与叶脉平行; 在上表皮的2个维管束之间分布有2~6个排列在一起、横切面呈扇形的泡状细胞,表皮上层有少量角质层; 下表皮分布有乳突、气孔器和粗毛,无耳藤竹着生粗毛较多,响子竹较少有或没有。2种攀缘竹的叶肉细胞没有栅栏和海绵组织分化,为等面叶,叶肉分布有大量自上而下的单向指状内褶叠的臂细胞,在维管束两侧有2个透明的梭形细胞,梭形细胞随维管束增大而变小。2种攀缘竹的主脉由多个维管束组成,二级脉只有1个维管束,维管束鞘细胞1~3层,均没有形成花环结构,为C3植物。

图2 2种攀缘竹叶片解剖特征Fig. 2 Anatomical characteristics in the leaves of two climbing bamboo species a.无耳藤竹叶片横切面; b.响子竹叶片横切面; c.无耳藤竹上表皮; d.响子竹上表皮; e.无耳藤竹下表皮; f.响子竹下表皮。UC: 上角质层; UE: 上表皮细胞; LE: 下表皮细胞; EH: 表皮毛; MP: 乳突; BC: 泡状细胞; BS: 维管束鞘; V: 导管; Ph: 韧皮部; Xy: 木质部; St: 气孔; LC: 长细胞; SC: 短细胞; FC:梭形细胞。a. Leaf transverse section of D. orenuda; b. Leaf transverse section of B. levigata; c. Upper epidermal thickness of D. orenuda; d.Upper epidermal thickness of B. levigata; e.Lower epidermal thickness of D. orenuda; f. Lower epidermal thickness of B.levigata. UC: Upper stratum corneum; UE: Upper epiderma; LE: Lower epiderma; EH: Epidermal hair; MP: Mastoid process; BC: Bulliform cell; BS: Vascular bundle sheath; V: Vessels; Ph: Phloem; Xy: Xylem; St:Stomata; LC: Long cell; SC: Short cell; FC: Fusoid cell.
2种攀缘竹的叶厚、泡状细胞数量、泡状细胞截面积、二级维管束直径、二级维管束截面积、二级维管束间距和气孔器面积占比均为雨季月份高于旱季月份(表1); 此外,单个梭形细胞截面积、气孔长度、单个气孔器面积在6月或9月达到最高,响子竹的上角质层厚度旱季高于雨季。2竹种的乳突厚度、下表皮厚度、气孔宽度和气孔密度最大值在各月表现不同。在各月的17个解剖结构指标中,2竹种的二级维管束直径、二级维管束截面积、气孔宽度和单个气孔器面积4个指标和无耳藤竹的泡状细胞数量、气孔长度以及响子竹的上角质层厚度、气孔器面积占比的最大值与最小值的差异均达显著水平(P<0.05),说明这些指标对月份变化较敏感。以上结果表明攀缘竹大部分解剖结构指标值在雨季较大,旱季较小; 二级维管束直径、二级维管束截面积、气孔宽度和单个气孔器面积4个指标较为敏感,其余12个指标的月份变化较小,较为稳定。
从2个竹种对比来看(表1),无耳藤竹的叶厚、气孔宽度、单个气孔器面积、气孔器面积占比在各月均高于响子竹,乳突厚度、上表皮厚度、二级维管束间距、气孔密度均低于响子竹。此外,无耳藤竹的上角质层厚度、泡状细胞数量、泡状细胞截面积、气孔长度在雨季各月高于响子竹,而下表皮厚度在雨季各月低于响子竹。2种攀缘竹的叶厚、乳突厚度、气孔宽度、单个气孔器面积、气孔密度和气孔器面积占比差异显著(P<0.05),说明叶厚、乳突和气孔是2个竹种叶片解剖结构差异的关键指标。
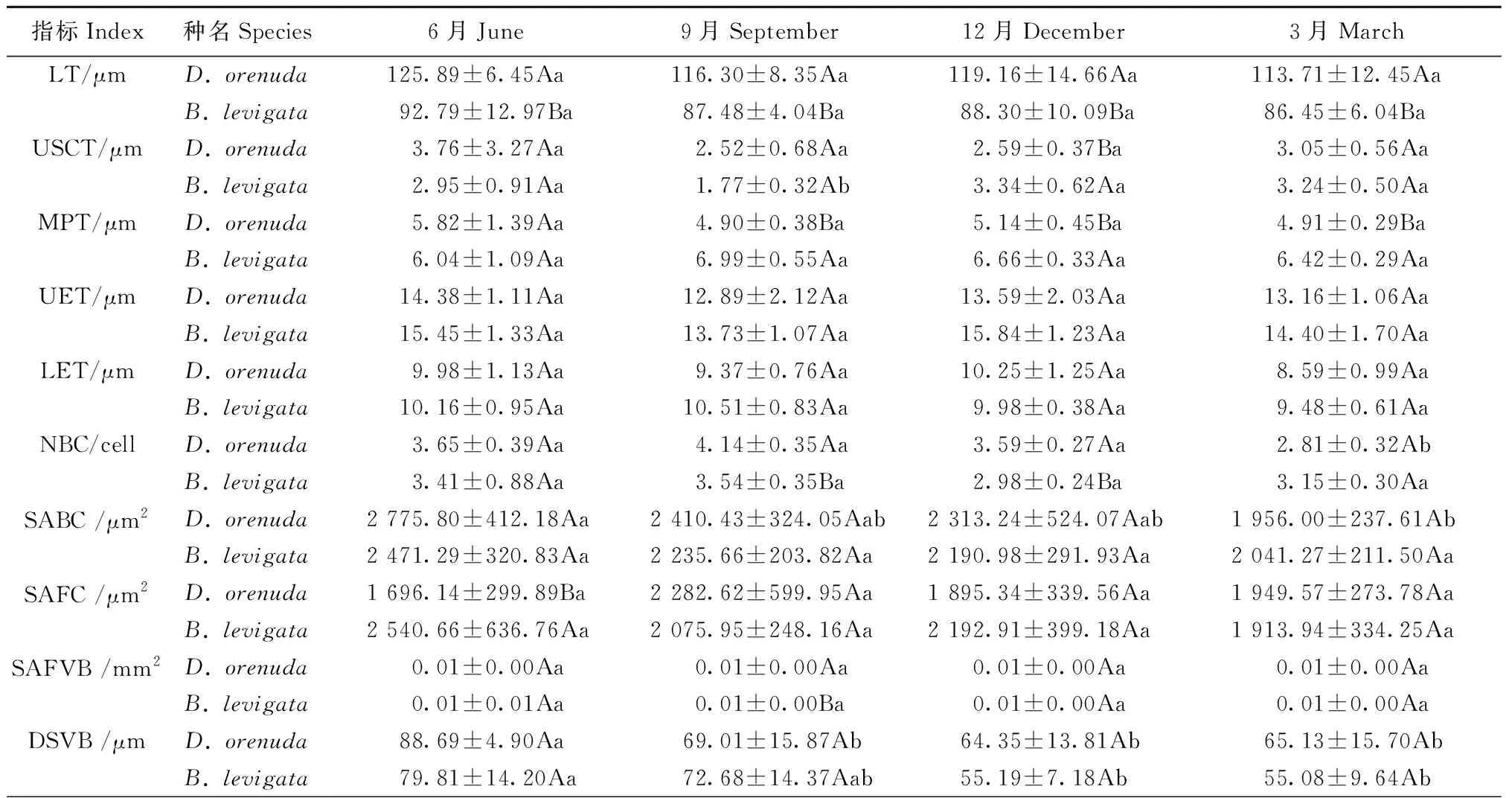
表1 2种攀缘竹叶片解剖结构月份动态①Tab.1 Monthly dynamics of anatomical characteristics in the leaves of two climbing bamboo species
3.3 叶片形态性状的月份变化 各月的叶片形态性状测定结果存在差异(图3)。无耳藤竹的单叶面积在9月最高,6月最低; 响子竹则在6月最高,3月最低,最高和最低值之间的差异显著(P<0.05)。叶片干物质含量在6月最低,最高值在2个竹种表现不同,2个竹种的最高值和最低值之间差异显著(P<0.05)。2个竹种的比叶面积在6月和3月较高,9月和12月较低,最高和最低值之间差异显著(P<0.05)。以上结果表明攀缘竹的叶片形态性状存在明显的季节差异。
在2个竹种之间,响子竹的单叶面积和干物质含量(6月除外)高于无耳藤竹(图3)。无耳藤竹的叶子干物质含量在雨季高于响子竹,在旱季则相反。除6月的单叶面积、12月和3月的叶片干物质含量外,2个竹种的各形态性状指标差异不显著(P>0.05)。以上结果表明2个竹种的叶片形态性状指标存在差异,但差异不明显。
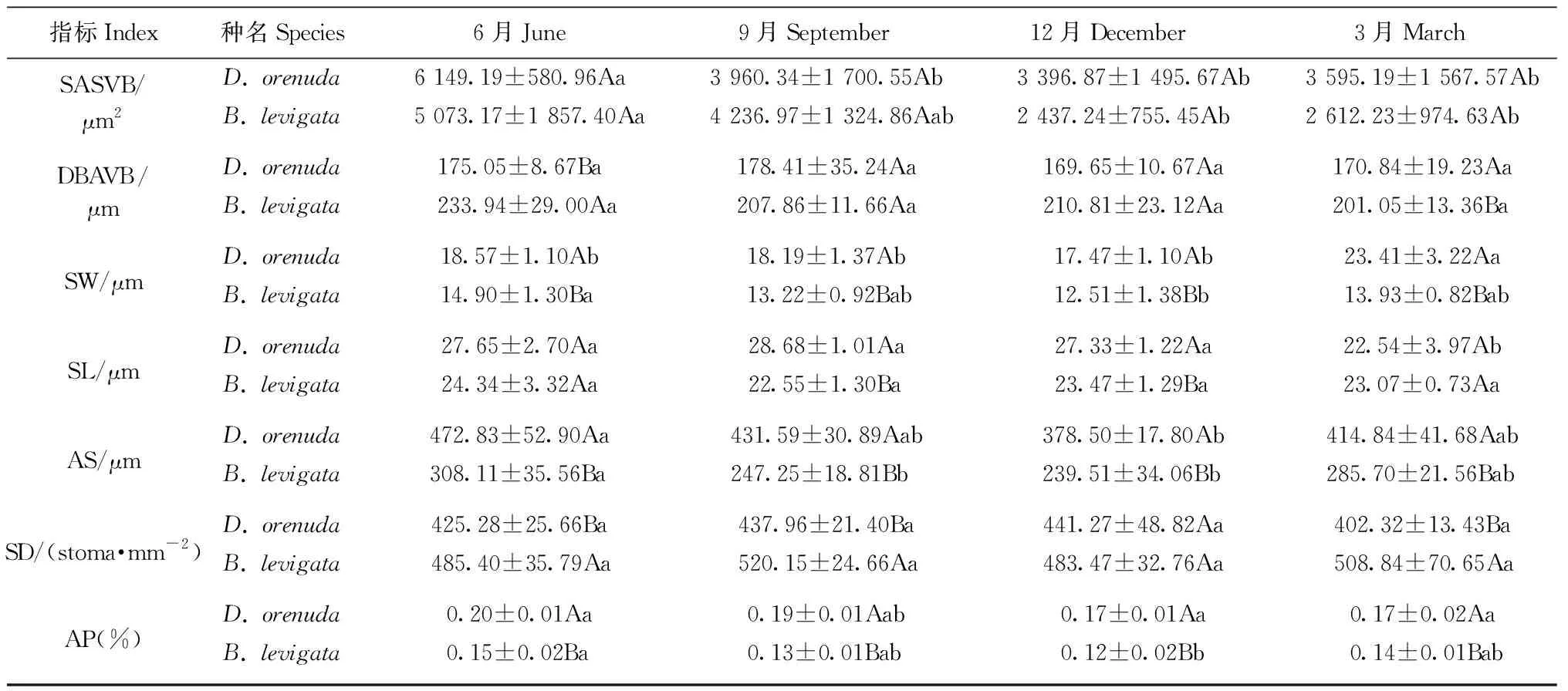
续表1 Continued
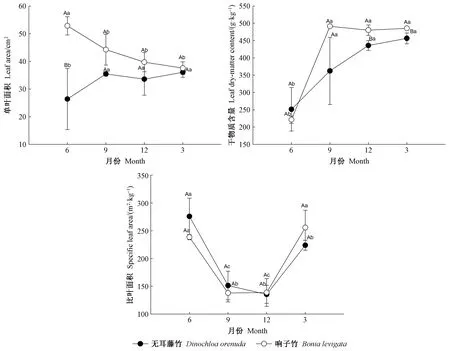
图3 2种攀缘竹叶片形态性状月份变化Fig. 3 Monthly dynamics of morphological traits in the leaves of two climbing bamboo species
3.4 叶片指标的可塑性 攀缘竹各月的叶片指标可塑性对比表明(表2),无耳藤竹的各指标平均可塑性在9月最高(0.34),12月最低(0.27),响子竹则是6月最高(0.28),9月最低(0.22)。无耳藤竹的各月平均可塑性指标均高于响子竹,全年的各指标平均可塑性(0.53)也高于响子竹(0.49),反映了无耳藤竹的环境适应能力高于响子竹。2竹种的气孔导度、蒸腾速率、上角质层厚度、主脉截面积和二级维管束截面积的可塑性较大,表明这些指标具有较大环境适应性。胞间二氧化碳浓度、叶厚、上表皮厚度、下表皮厚度、单个气孔器面积、气孔密度的可塑性均较低,表明这些指标具有较高的稳定性。
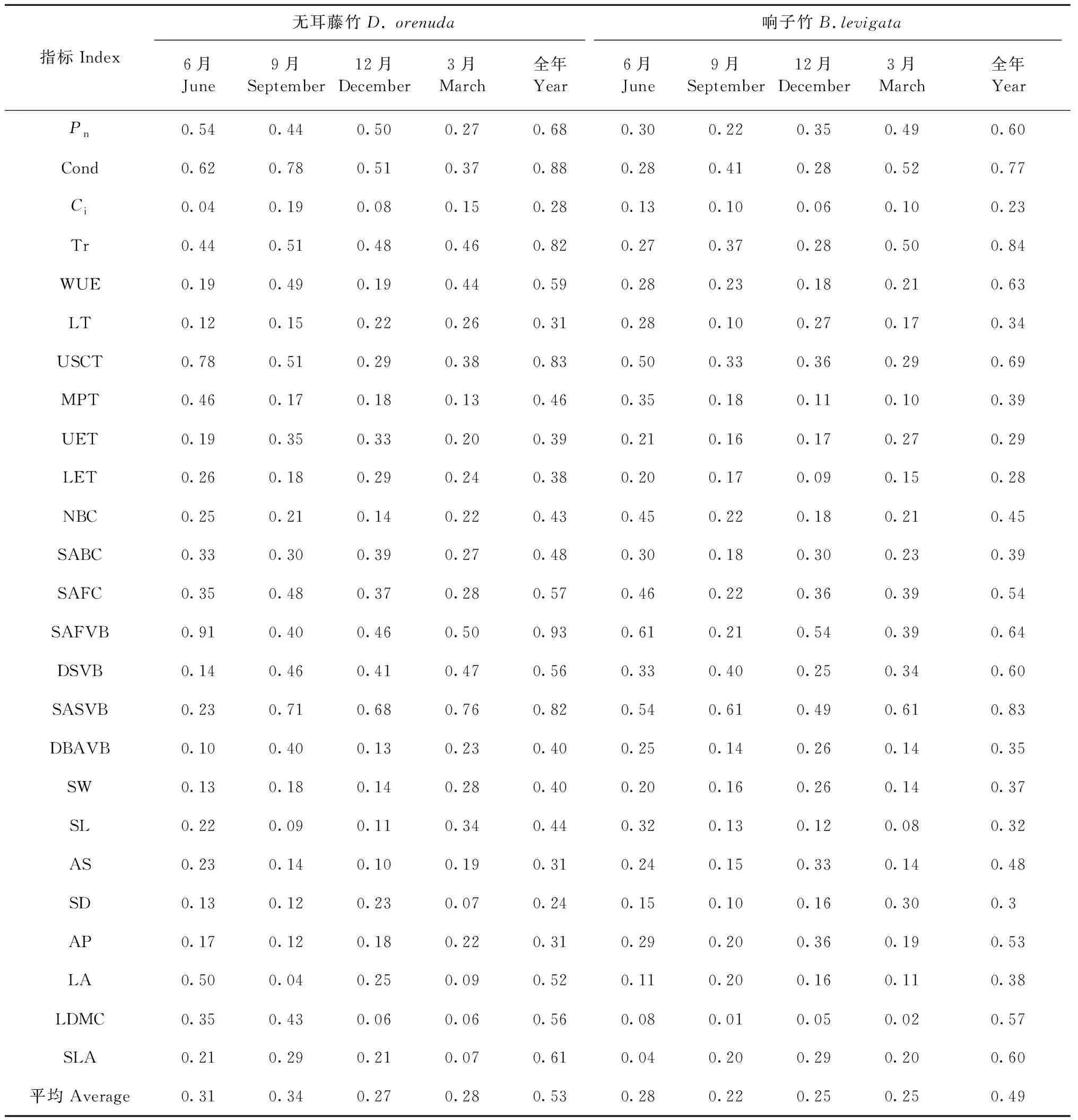
表2 2种攀缘竹叶片可塑性指数①Tab.2 Plasticity index in the leaves of two climbing bamboo species
3 讨论
3.1 2种攀援竹叶片功能性状的月份动态 2种攀援竹叶片的净光合速率、气孔导度、胞间二氧化碳浓度和蒸腾速率在雨季高于旱季,与云南季雨林的网脉核果木(Drypetesperreticulata)和福建红树林的秋茄(Kandeliaobovata)等树种的叶片光合特性研究结果(栗忠飞等, 2012; 黄丽, 2013)一致; 水分利用效率和叶片光合能力表现与前期开展的攀缘竹光合特性研究结果(徐瑞晶等, 2020)一致,说明2竹种的光合作用年际间较稳定,这是2种攀缘竹长期适应热带低地雨林生境的结果。
攀缘竹的叶厚、乳突厚度、气孔宽度、气孔密度等解剖结构指标值在雨季大于旱季,反映了攀缘竹叶片在雨季通过增大以上组织结构以适应雨季快速生长。同时,二级维管束直径、二级维管束截面积和单个气孔器面积在雨季显著高于旱季(P<0.05),可有效增强竹叶的物质运输和光合气体交换能力,促进攀缘竹在雨季快速生长。
叶片性状的季节变化可反映植物的环境适应能力,不同指标间的表现是植物生长策略权衡的结果(薛立等, 2012)。2竹种的比叶面积在6月和3月较高,9月和12月较低,主要原因是竹叶的雨季光合作用增强,叶片积累了大量的有机物,其干质量增加,表现出具有较低的比叶面积; 而进入旱季后,叶片光合能力下降,降低了积累有机物的能力,因此比叶面积处于较高水平。2竹种叶片的干物质含量的月份变化趋势与其光合作用有机物积累的季节变化相关。2竹种的单叶面积变化特征与它们的叶片生长发育规律有关,其新生叶片的单叶面积在前期较小,在发育到一定阶段后基本保持不变。
3.2 2种攀缘竹的叶片适应策略 植物叶片可塑性越大,其种群分布越广泛,生态适应性越强(蒋琴等, 2018)。作者野外调查中发现,无耳藤竹在热带雨林分布更广泛,响子竹种群分布范围较小,这与藤竹叶片可塑性有关。海南岛甘什岭热带低地雨林区,雨季降雨量占全年的90%以上,雨季和旱季更替是该区重要的环境变化。本研究中,2竹种的蒸腾速率、角质层厚度、主脉截面积和二级维管束截面积的可塑性均较高,它们是水分利用的重要结构指标,通过这些指标的调节利于适应雨季旱季更替引起的环境变化。无耳藤竹的乳突厚度、上表皮厚度、二级维管束间距、气孔密度均低于响子竹。此外,无耳藤竹的上角质层厚度、泡状细胞数量、泡状细胞截面积、气孔长度在雨季高于响子竹,而下表皮厚度在雨季低于响子竹。2种攀缘竹的叶厚、乳突厚度、气孔宽度、单个气孔器面积、气孔密度和气孔器面积占比差异显著(P<0.05),叶厚、乳突和气孔是反映2个竹种叶片解剖结构差异的关键指标。
植物叶性状与植物的生长对策和植物的资源利用能力密切相关(Reichetal., 2010)。本研究表明,无耳藤竹的叶片净光合速率、叶厚、气孔宽度、单个气孔器面积和气孔器面积占比在各月均显著高于响子竹,但二级维管束间距低于响子竹。无耳藤竹有极强的攀爬能力,其枝条可攀抵热带雨林的林冠层,叶片更适应强光环境,这与马光天等(2018)研究认为强光环境下,叶片通过增大叶厚减小单叶面积以减少光辐射量,避免强光对叶片的伤害; 弱光环境下,植物倾向于减小叶厚增大单叶面积,以增强叶片细胞对弱光的捕获能力的研究结果一致。林冠层叶片偏向旱生结构,在结构上偏向于减少水分散失,在功能上提高水分利用效率,从而适应水分亏缺(何春霞等, 2013)。叶厚是评价植物干旱适应性的重要指标,叶片越厚则适应干旱能力越强(马建静等, 2012),较厚叶片可帮助无耳藤竹缓解林冠层水分亏缺和旱季雨量减少造成的干旱胁迫。气孔宽度、单个气孔器面积和气孔器面积占比是控制叶片光合作用气体交换的关键结构指标,它们的较高数值利于提高叶片同化二氧化碳的效率。无耳藤竹叶片的二级维管束间距较小,说明其叶片单位面积内维管束密度较高,输送养分水分的功能更强; 以上叶片特征有助于无耳藤竹提高其光合和攀爬能力、适应冠层水分亏缺和旱季胁迫。与无耳藤竹相反,响子竹的净光合速率、气孔宽度、单个气孔器面积、气孔器面积占比较低,很难抵达林冠层,决定了其较为耐荫的习性; 乳突可减少气孔水分蒸发,利于响子竹适应逆境胁迫(蒋琴等, 2018),响子竹凭借较厚的乳突和上表皮、数量多而面积小的气孔来减少水分蒸发,可更高效地利用水分(王媛媛等, 2018),以适应旱季胁迫。
3.3 2种攀缘竹资源的保护和利用 无耳藤竹和响子竹均自然分布于热带雨林中,其攀援特性决定了必须依靠其他植物才能更好生长。尤其是无耳藤竹,需依靠其他树木的支撑才能攀至林冠层,满足其喜阳特性的生境要求; 响子竹较耐荫,不需抵达林冠层,但其竿茎纤细柔软,必须依附于其他树木才能更好生存。因此,为保护攀缘竹,在加强攀缘竹种群和个体保护的同时,还要重视对其群落和支撑树木的保护。在攀缘竹利用上,无耳藤竹和响子竹植株均呈攀援或悬垂状,明显区别于直立竹,具有较高观赏价值。响子竹竹竿纤细,叶片纤长柔美,竹态曼妙,可作为廊道、花架和墙体等垂直绿化特色竹种; 无耳藤竹的竹茎可达3 cm,柔韧性较好,可作为编织材料,其竹笋可食用,颇受当地农民喜爱,亦可作特色笋用竹。在攀缘竹资源培育上,2种攀缘竹属特殊的丛生竹,可采用分蔸移植。无耳藤竹较喜阳,苗期管理时要保证一定光照强度,而响子竹较耐荫,需营造遮荫环境。在野外状态下,2种攀缘竹的光饱和点存在一定的月份差异,光照条件营造需根据月份适当调整。无耳藤竹和响子竹的各月平均光饱和点分别为(1 032±217)和(877±183) μmol·m-2s-1(徐瑞晶等, 2020),可作为其育苗光照条件的参考,但苗期的最适光照强度还需进一步研究。无耳藤竹和响子竹的光合能力均在雨季月份(6月和9月)最高,培育竹苗时这段时间内要加强水肥供给,以保证竹苗更好生长; 在旱季月份(12月和3月),要保证足够水分供给,减少干旱对生长的胁迫。
4 结论
2种攀缘竹叶片的结构、光合特性和形态性状特征决定了它们不同的生态习性和生存策略。无耳藤竹和响子竹在雨季通过增大与物质运输和光合气体交换相关的结构,来实现快速生长,同时为即将到来的旱季胁迫做好物质储备。无耳藤竹具有较高的叶片指标可塑性,表明其有更强的旱季适应性,能凭借较大的光合能力、叶厚、气孔和较低的单叶面积、比叶面积、二级维管束间距等指标,来适应林冠层的强光环境、冠层水分亏缺和旱季胁迫。响子竹则通过其叶子的较高的单叶面积、比叶面积、上表皮厚度、乳突厚度和较薄的叶片及小而密的气孔,适应旱季胁迫和林下弱光环境。
