不同作物轮作对谷田土壤酶活性和土壤细菌群落的影响
2021-01-28孙倩吴宏亮陈阜康建宏
孙倩,吴宏亮*,陈阜,康建宏
1. 宁夏大学农学院,宁夏 银川 750021;2. 中国农业大学农学院/农业农村部农作制度重点实验室,北京 100193
在生产上已经发现,作物轮作能减轻连作障碍,平衡土壤养分,有利于作物生长和提高产量,并对土壤微生物和酶活性具有重要影响(吴凤芝等,2007;Anna et al.,1999)。土壤微生物是土壤生态系统的重要组成部分,对维持土壤功能、生态系统稳定等具有重要作用(郑良永等,2005)。土壤酶活性是评价土壤肥力、土壤微生物活性的重要参数,其参与土壤中各种生物化学过程,可为植物的生长提供氮源、磷素等(董艳等,2009)。Alvey et al.(2003)对谷物/作物轮作下土壤细菌群落结构进行研究,认为种植制度对细菌群落结构的影响较大,而植物种类和取样时间的影响不大;轮作可以明显改变细菌群落的组成。吴宏亮等(2013)研究发现,轮作能够提高砂田土壤微生物多样性指数,改善微生物区系结构。邓玉清等(2018)研究了轮作对番茄连作土壤微生物群落结构、酶活性及养分的影响,发现番茄与秋葵或花生轮作均可提高土壤微生物多样性,提高土壤酶活性,有利于维持土壤生态环境。牛倩云等(2018)研究发现轮作改变了土壤理化性质、酶活性和根际土壤细菌多样性指数,并且提高了部分有益功能菌的丰度。
目前,关于作物轮作能够缓解连作障碍的研究报道较多(Tan et al.,2016;李锐等,2015),但针对谷子等杂粮作物的相关研究却很少。因此,本研究设置谷子→黑豆、谷子→籽粒苋、谷子→藜麦和谷子连作4种处理,分别分析各作物成熟期根际土壤酶活性、细菌群落结构和多样性的变化,旨在全面深入地了解谷子连作及轮作后土壤酶活性和微生物群落的变化,以期为揭示谷子连作障碍及采取合适的轮作措施提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于宁夏吴忠市同心县王团镇,该区域属宁夏中部干旱带,典型的中温带干旱、半干旱大陆性气候,干旱少雨,海拔 1370 m,无霜期 120—218 d,年降水量150—300 mm,多集中在7—8月,年蒸发量在2325 mm左右,年平均气温8.6 ℃,≥10 ℃的积温约3000 ℃。该区土壤类型为砂壤土。试验地土壤基本理化性质为:pH:8.76,有机质:12.66 g·kg-1,全氮:0.66 g·kg-1,速效钾:440.39 mg·kg-1,速效磷:1.00 mg·kg-1。
1.2 试验材料
供试谷子(Setaria italica)品种为“豫谷18号”,黑豆(Glycine max)品种为“宁黑1号”,籽粒苋(Amaranthus hybridusL.)品种为“旱雷神”,藜麦(Chenopodium quinoa)品种为“蒙藜1号”。
1.3 试验设计
试验采用单因素随机区组试验设计,共设4个处理,分别为:谷子→黑豆(MRG)、谷子→籽粒苋(MRA)和谷子→藜麦(MRQ)和谷子连作(CK),每个处理5次重复,小区面积25.6 m2,小区间用田埂隔开,不同小区的管理方式相同。
1.4 土壤样品的采集
于2018年10月作物成熟收获时进行采集。每小区选取3个采样点,同时选取长势一致的作物3株,然后挖取作物的整个根部,先去掉大块土和附着土,用刷子刷取根毛上的土,即为根际土,将每样点3株作物的根际土混匀作为该样点的根际土。将取得的根际土装入无菌自封袋,于冰盒保存带回实验室,迅速过2 mm网筛,一部分保存于-80 ℃冰箱中,用于后续土壤微生物测定,另一部分自然风干,用于土壤酶活性的测定。
1.5 测定指标及方法
1.5.1 土壤酶活性的测定方法
土壤过氧化氢酶活性的测定采用高锰酸钾滴定法;土壤脲酶活性的测定采用苯酚钠-次氯酸钠比色法;土壤蔗糖酶活性的测定采用3, 5-二硝基水杨酸比色法;土壤碱性磷酸酶活性测定采用磷酸苯二钠比色法(关松荫,1986)。
1.5.2 土壤总DNA的提取及16S rRNA扩增测序
使用快速 DNA旋转提取试剂盒(MP Biomedicals,Santa Ana,CA,USA)提取样本基因组总DNA,并用紫外分光光度计和琼脂糖凝胶电泳分别测定提取的 DNA的量和质量。采用 ABI 2720型PCR扩增仪,选取细菌16S rRNA基因V3—V4区进行PCR扩增,扩增选用前引物(5′-GTGCC AGCMGCCGCGG-3′)和后引物(5′-CCGTCAATTC MTTTRAGTTT-3′)合成的引物,进行PCR扩增。反应体系:q5反应缓冲液(5×5)5 μL,q5高保真GC缓冲液(5倍)5 μL,q5高保真DNA聚合酶(5 U·μL-1)0.25 μL,dntps(脱氧核糖核酸)(2.5 mm)2 μL,正向和反向引物(10 μm)各 1 μL、DNA模板2 μL和ddH2O 8.75 μL。扩增参数:98 ℃下的初始变性15 s,然后是25个循环,包括98 ℃15 s,55 ℃退火30 s,72 ℃延伸30 s,最终延伸5 min 72 ℃,使产物延伸完整,4 ℃保存。扩增结果用2%琼脂糖凝胶电泳检测,切取目的片段后用Axygen凝胶回收试剂盒回收目的片段。将回收目的片段送至上海派森诺科技股份有限公司进行测序。
1.5.3 序列分析
对 Illumina的测序结果进行分析,首先运用QIIME(Caporaso et al.,2010)软件对测序的疑问序列进行过滤、检查并剔除嵌合体序列。通过以下标准过滤质量序列:长度<150 bp的序列,含有模糊碱基的序列以及含有大于 8 bp的单核苷酸重复序列。使用 QIIME软件,在 97%序列相似度下对操作分类单元进行聚类,并制作 Venn图;利用QIIME中的OTU数据计算Chao1指数(Chao,1984;Dawid,1993)、Shannon 指数(Shannon,1948)和Simpson指数(Simpson,1949)等OTU水平的α多样性指数,并生成OTU水平稀释曲线。利用R软件对细菌门和属进行聚类,并制作柱状图和热图。使用PICRUSt菌群代谢功能预测分析对各处理的细菌功能进行预测,并绘制成柱状图。
1.6 数据处理
使用Microsoft Excel 2010对土壤酶数据进行处理和作图,采用SPSS 20.0统计软件进行方差分析和多重比较(Duncan新复极差法),显著性水平设为P<0.05。使用Canoco 4.5软件对土壤酶活性和细菌群落进行多元分析。用Person相关分析检验土壤细菌多样性指数与土壤酶活性之间的相关性。
2 结果分析
2.1 作物轮作对土壤酶活性的影响
由图1可知,与连作相比,轮作下过氧化氢酶、脲酶和碱性磷酸酶活性均升高,除MRG处理的蔗糖酶活性低于连作外,其余两种轮作模式下的蔗糖酶活性也升高。过氧化氢酶和碱性磷酸酶活性在MRQ处理最高,MRG和MRA次之,CK最低。CK处理的过氧化氢酶活性分别比 MRQ、MRG、MRA降低了8.02%、5.2%、4.38%,且与MRQ呈显著性差异(P<0.05);CK 处理的碱性磷酸酶活性分别比MRQ、MRG、MRA降低了10.34%、1.65%、1.08%,但CK与MRG和MRA间的差异不显著,但与MRQ间差异显著(P<0.05)。脲酶活性的表现与过氧化氢酶活性相同,CK处理分别比MRQ、MRG、MRA降低了7.03%、6.12%、6.12%,但差异不显著。蔗糖酶活性也表现为MRQ处理最高,MRG和MRA次之,CK最低;CK处理分别比MRA、MRQ降低了 16.78%、10.27%,比 MRG提高了8.61%;其中,MRA与MRG和CK间的差异达显著水平。
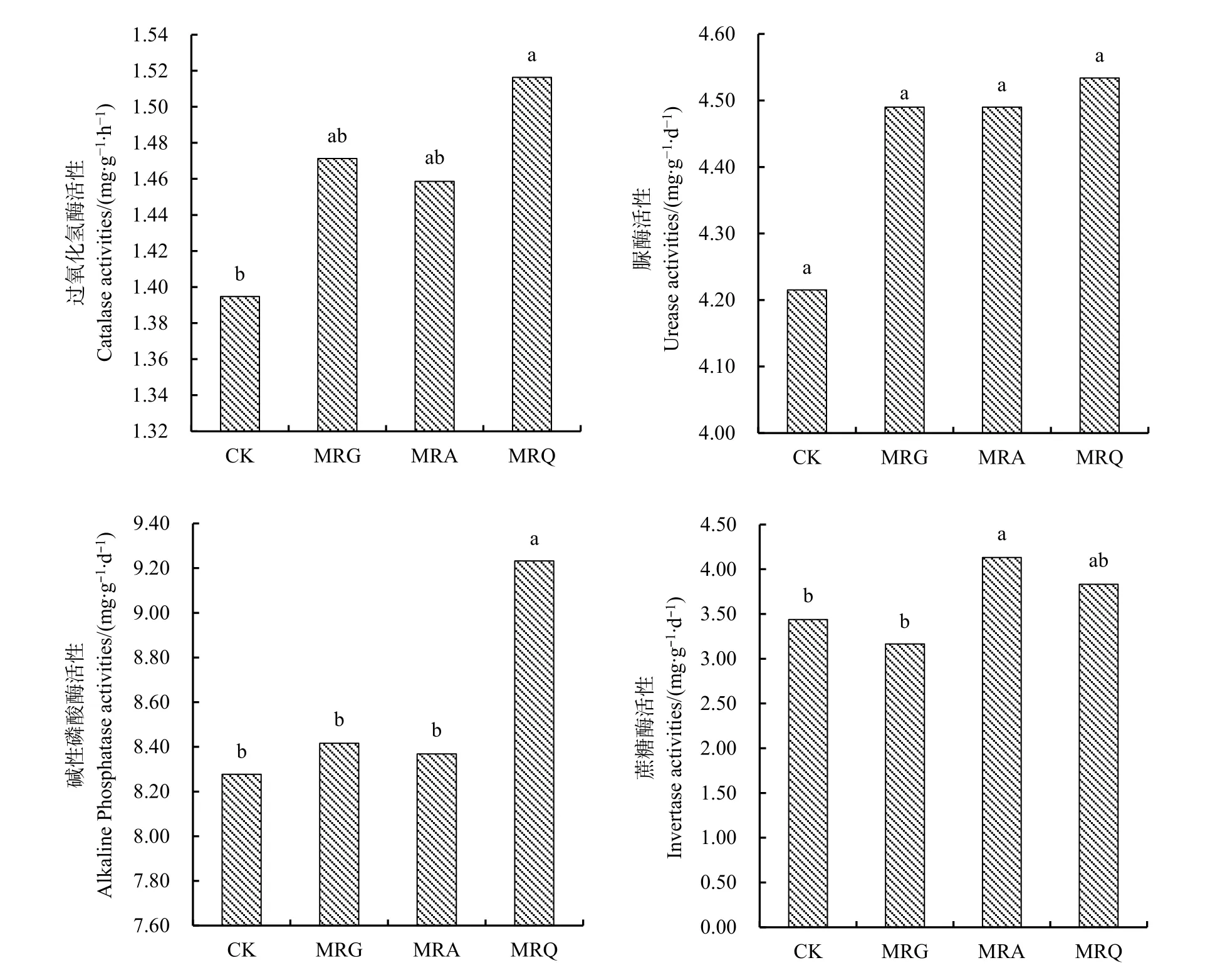
图1 不同轮作式下的土壤酶活性Fig. 1 Soil enzyme activity under different rotation patterns
2.2 作物轮作对土壤细菌群落的影响
2.2.1 作物轮作下根际土壤细菌16S rRNA 的V3—V4区原始数据分析
利用Illumina MiSeq高通量测序平台对细菌基因V3—V4区测序,共得到有效序列720015条,谷子→黑豆(MRG)、谷子→籽粒苋(MRA)、谷子→藜麦(MRQ)、谷子连作(CK)各处理所获得的细菌有效序列分别有33675、34108、32722、43498条,平均长度在405—435 bp之间。稀释曲线用来反映该测序深度是否合理、能否真实的反映样品中细菌物种的丰富度。由图2可知,各曲线均趋于平坦,表明该测序深度合理,测序结果可以用来分析细菌群落结构。
2.2.2 作物轮作下根际土壤细菌OTUs数
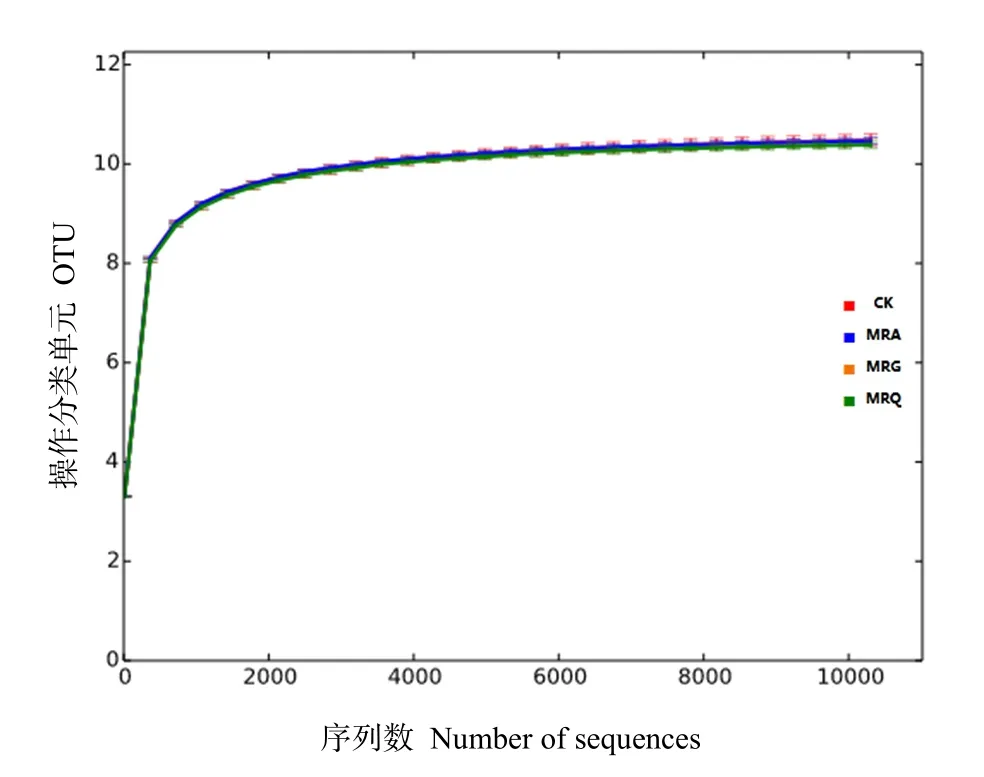
图2 稀释曲线Fig. 2 Dilution curve

图3 根际土壤细菌OTUs数Venn图Fig. 3 Venn diagram of bacterial OTUs in rhizosphere soil
在 97%序列相似度下对各处理土壤细菌的OTU进行归并和划分,并利用R软件绘制Venn图,结果可以很清楚的展示各处理共有和特有的 OTU数,并可直观的展示各处理 OTU的重叠情况。由图 3可知,谷子→黑豆(MRG)、谷子→籽粒苋(MRA)、谷子→藜麦(MRQ)、谷子连作(CK)共获得了10185个OTU,各处理分别获得了5616、5713、5561、6492个OTU,各处理共有2614个OTU,谷子→黑豆(MRG)、谷子→籽粒苋(MRA)、谷子→藜麦(MRQ)、谷子连作(CK)特有的OTU数为 864、787、846、1254,其余均为样本两两共有的。
2.2.3 作物轮作下土壤细菌多样性和丰富度分析
Simpson指数和Shannon指数是反映微生物群落的多样性指数,Chao1指数和ACE指数是反映微生物群落的丰富度指数。表1为各轮作处理下土壤细菌群落的多样性指数和丰富度指数。从表中可知,除MRQ轮作处理的细菌多样性指数低于连作外,其余两种轮作处理的多样性指数均高于连作;3种轮作处理的细菌丰富度指数均低与连作。与连作相比,MRG、MRA处理的多样性指数(Simpson指数、Shannon指数)分别提高了0.04%、0.34%和0.04%、0.21%;MRQ处理的多样性指数(Simpson指数、Shannon指数)降低了0.02%、0.57%;MRG、MRA、MRQ处理的丰富度指数(Chao1指数、ACE指数)分别降低了 12.98%、18.40%和 15.90%、18.96% 、35.84%和40.66%。由于轮作后群落中优势种数量(比例)的下降,导致多样性指数增加,由此可以推断,轮作可以显著降低细菌群落丰富度指数,而增加了多样性指数。
2.2.4 作物轮作下土壤细菌群落结构分析
门水平下各处理的细菌群落结构显示(图4),相对丰度大于1%的门类有7门,分别为放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)、芽单胞菌门(Gemmatimonadetes)、拟杆菌门(Bacteroidetes)和 Saccharibacteria。其中,放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)为优势菌门,在各处理中的占比达74.86%—81.68%。与连作相比,变形菌门、绿弯菌门、酸杆菌门、芽单胞菌门的相对丰度增加,分别增加了 4.78%—18.71%、0.91%—27.40%、25.09%—85.71%和 11.28%—37.74%,而放线菌门相对丰度降低,分别降低了8.45%—30.99%。
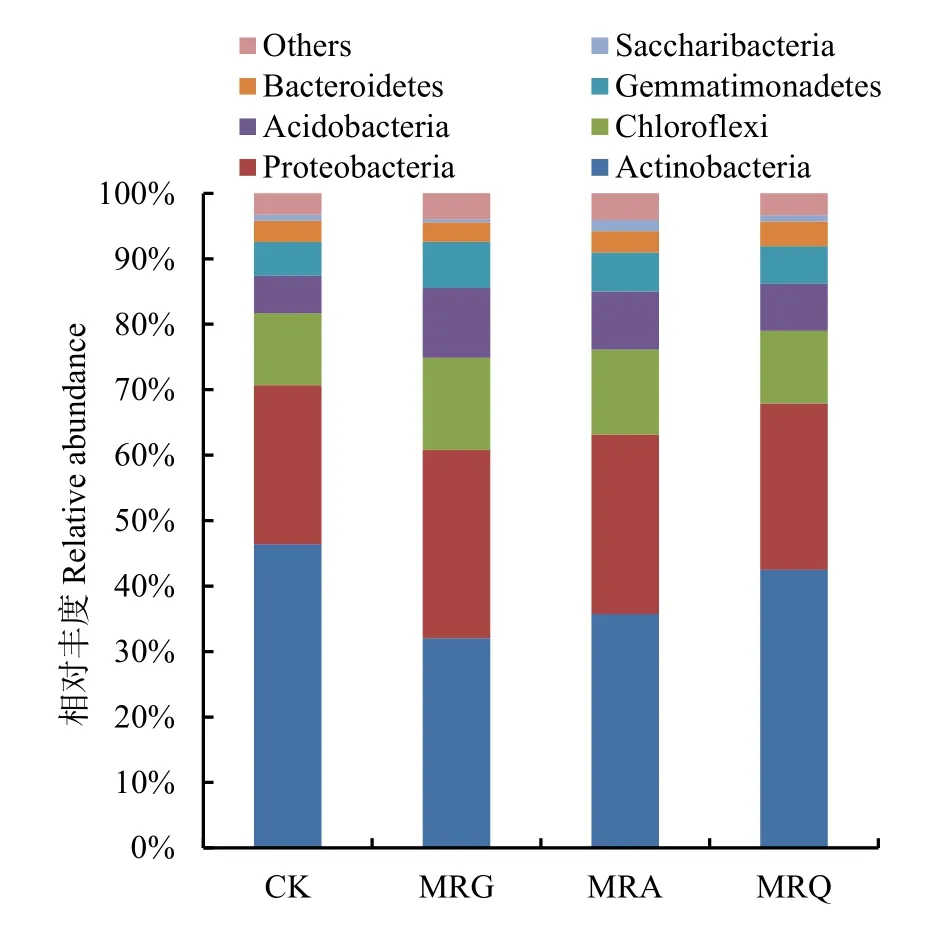
图4 门水平的细菌群落组成及丰度Fig. 4 Bacterial community composition and abundance at phylum level

表1 各轮作处理下土壤细菌多样性指数Table 1 Bacterial diversity index of soil under different rotation treatments
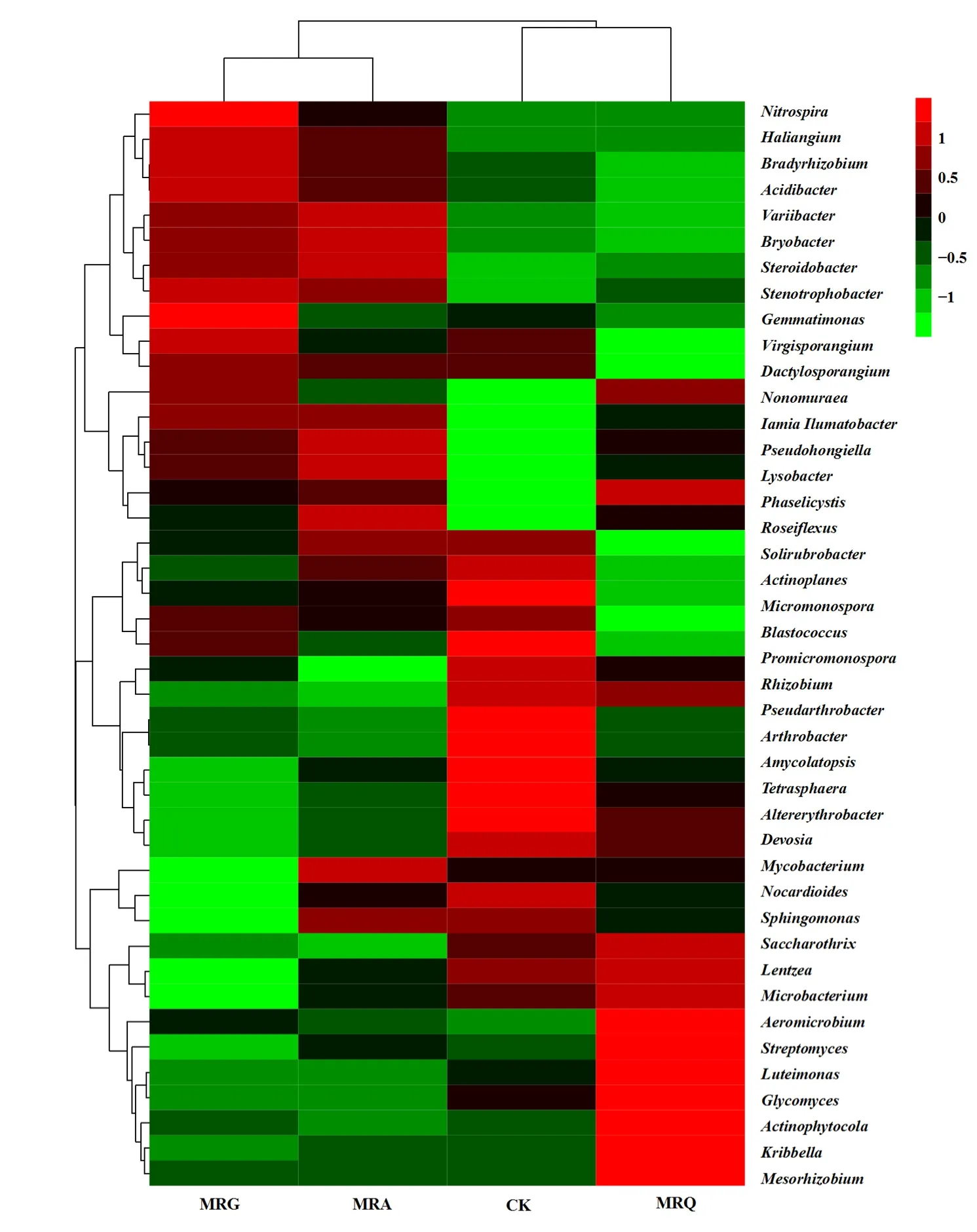
图5 属水平的细菌群落组成Fig. 5 Bacterial community composition at genus level
将相对丰度大于 1%的细菌属进行聚类,发现各处理中优势细菌类群发生改变。由图 5可知,MRA和MRG处理聚为一支,CK和MRQ聚为一支,说明MRA与MRG、CK与MRQ相似的优势细菌属较多。CK与MRQ处理共有的优势属有拟诺卡氏菌属Nocardioides、鞘氨醇单胞菌属Sphingomonas、伦茨氏菌属Lentzea、Pseudarthrobacter、链霉菌属Streptomyces、Saccharothrix、Kribbella、Iamia和Aeromicrobium;MRA与MRG处理共有的优势均属有鞘氨醇单胞菌属Sphingomonas、拟诺卡氏菌属Nocardioides、Haliangium和Aeromicrobium。各处理共有的优势均属有鞘氨醇单胞菌属Sphingomonas(2.25%—3.11%)、拟诺卡氏菌属Nocardioides(1.65%—4.12%)和Aeromicrobium(1.05%—1.37%)。
2.2.5 PICRUSt菌群代谢功能预测分析
根据 PICRUSt预测的各处理根际土壤菌群代谢功能(图6),共发现12种主要的二级基因功能代谢通路(相对丰度>1%)。从图可知,碳水化合物和氨基酸代谢显著高于其他代谢通路(P<0.05)。其次,发现轮作处理提高了聚糖生物合成代谢和酶家族代谢通路。
2.3 土壤酶活性与土壤细菌群落和多样性的相关性分析
2.3.1 土壤酶活性与细菌属的冗余分析
为进一步研究不同处理下土壤酶活性和细菌群落之间的相关关系,将土壤酶与土壤细菌属类群作冗余分析(RDA)。冗余分析(RDA)结果显示(图7),第一排序轴贡献率为65.4%,第二排序轴贡献率为 26.9%,排序轴能在超过 92.3%%的累计贡献率上解释土壤细菌群落与土壤酶活性之间的关系。从图中可知,相同处理的点分布较为集中,而不同处理的点分布较为分散。此外,还可知,4种酶活性与多数细菌属呈正相关,说明它们受酶活性的影响较大。
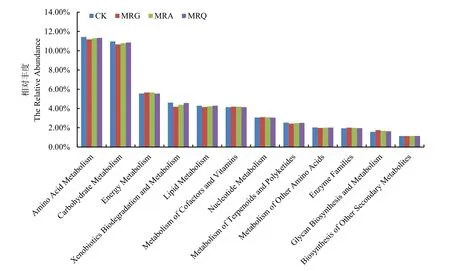
图6 轮作模式下作物根际土壤菌群代谢功能预测Fig. 6 Prediction of microbial metabolism in rhizosphere soil under crop rotation

图7 土壤细菌属和土壤酶活性的多元分析Fig. 7 Multivariate analysis of soil bacteria and soil enzyme activities
2.3.2 土壤酶活性与细菌多样性指数和丰富度指数的相关性分析
为进一步了解不同处理下土壤酶活性和细菌多样性和丰富度之间的相关关系,将土壤酶与土壤细菌属类群作相关分析(表 2)。由表可知,过氧化氢酶和碱性磷酸酶活性与细菌多样性指数(Simpson指数、Shannon指数)和丰富度指数(Chao1指数、ACE指数)均呈负相关关系;其中,与ACE指数的相关性达显著水平。脲酶活性与多样性指数(Simpson指数、Shannon指数)均呈正相关关系,但不显著。蔗糖酶仅与Shannon指数呈正相关,相关性也不显著。

表2 土壤细菌多样性指数与土壤酶活性的相关性分析Table 2 Correlation analysis between soil bacterial diversity index and soil enzyme activity
3 讨论
土壤酶活性反映了土壤中各种生物化学过程的强度和方向,与土壤微生物数量、全氮含量、有机质含量等密切相关。研究显示,轮作有利于土壤微生物的繁殖,增加微生物数量,增强代谢能力,使土壤维持较高的微生物活性与多样性,增加土壤肥力水平,提高土壤酶活性(张成君等,2020)。本研究结果显示,轮作提高了土壤过氧化氢酶、土壤磷酸酶和脲酶的活性,只是影响效果不显著,这与曾玲玲等(2008)的研究结果相似。
轮作改变了土壤微生物群落结构,提高了土壤微生物多样性。本研究结果显示,与谷子连作相比,谷子→黑豆和谷子→籽粒苋轮作下细菌多样性指数升高,各轮作处理的细菌丰富度指数均降低,分析其原因可能是优势菌门(属)比例的降低而导致的。此外,本研究发现放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)为各处理的优势菌门,这与Edward et al.(2015)在水稻、Niu et al.(2017)在玉米及曹鹏熙等(2020)在冰川棘豆上的研究结果相类似。在属水平上,各处理的优势菌属存在差异,其中以 MRA与 MRG较为相似,CK与MRQ处理较为相似;鞘氨醇单胞菌属Sphingomonas、拟诺卡氏菌属Nocardioides和Aeromicrobium是各处理共有的优势属。土壤细菌群落的这些变化归因于不同作物根系分泌物的差异,因为根系分泌物是植物与根际微生物相互沟通的媒介,不同作物的根际分泌物的种类不同,而微生物的生长又会受到这些分泌物的影响,从而导致微生物的生长和多样性发生变化,进而影响微生物群落结构的变化(Hu et al.,2018)。
土壤微生物群落结构和多样性受多种因素的影响,包括土壤化学组成、土壤类型、植被类型、人类活动、气候条件等(杨瑞红等,2016;Zhao et al.,2010;Archer et al.,2019)。刘岳燕等(2006)和李华等(2014)认为土壤微生物多样性受土壤酶活性的影响很大。本研究对土壤细菌群落与土壤酶活性的冗余分析表明,排序轴能在超过92.3%的累计贡献率上解释细菌群落结构与酶活性的关系;其次,本研究中的4种土壤酶活性与多数细菌属呈正相关关系,仅与少数细菌属呈负相关关系,这说明细菌群落受土壤酶活性的影响较大。这与前人的研究结果一致(于高波等,2011)。通过对土壤细菌多样性和酶活性进行相关性分析,发现过氧化氢酶和碱性磷酸酶活性与细菌多样性指数和丰富度指数均呈负相关关系,其中与 ACE指数呈显著性负相关,这说明过氧化氢酶和碱性磷酸酶活性对某些细菌的生长繁殖有抑制作用。而脲酶和蔗糖酶与细菌多样性指数呈正相关,这说明脲酶和蔗糖酶活性的增强会促进某些细菌的繁殖,这是因为脲酶是参与氮素转化的关键酶,脲酶活性的增强会提高土壤氮素肥力,从而影响了微生物群落结构(彭有才等,2009);蔗糖酶能够促进有机物质的分解,为微生物的生长提供充足的养料,从而促进微生物的繁殖(杨阳等,2011)。
土壤酶活性和微生物群落结构受多种因素的共同影响,本试验只研究了土壤酶活性对细菌群落结构的影响,并且针对的是轮作一茬的作物,因此建议持续进行试验,观察轮作倒茬的效果。研究显示土壤养分是土壤微生物生长的主要能量来源,被认为是调控土壤微生物数量和群落结构的关键因素(罗达等,2017;楚海燕等,2019),因此建议后续结合土壤理化性状、作物根系分泌物等进行研究。
4 结论
作物轮作对土壤酶活性、根际土壤细菌群落结构和多样性等均产生影响。与连作相比,除谷子→黑豆轮作模式下的蔗糖酶活性略有降低外,其余轮作模式下的土壤酶活性均增大,其中,以谷子→藜麦处理增加最明显。与连作相比,轮作改变了作物根际土壤细菌群落结构,改变了细菌多样性指数和丰富度指数,过氧化氢酶和碱性磷酸酶活性对细菌丰富度指数的影响较大。细菌丰富度指数表现为连作处理高于轮作处理,由于其中优势种数量(比例)的降低使得细菌多样性指数以谷子→黑豆、谷子→籽粒苋处理高于连作,因此推荐其为较好的轮作作物。
