植物群落和土壤对人工草地恢复演替的协同响应
2021-01-28潘攀王长庭胡雷刘斯莉李洁
潘攀,王长庭*,胡雷,刘斯莉,李洁
1. 西南民族大学青藏高原研究院,四川 成都 610225;2. 西南民族大学生命科学与技术学院,四川 成都 610225
三江源区作为青藏高原的腹地,是高寒草甸的集中分布区(王琰,2015),近年来由于受到气候变化(全球变暖)、鼠虫危害及人类活动(过度放牧)的影响,90%以上的草地出现了不同程度的退化(李穗英等,2007),部分严重退化为“黑土滩”,对当地和下游地区生态安全、社会和经济的可持续发展造成了恶劣的影响(Dong et al.,2010)。为了遏制高寒草甸的退化,目前已有许多的修复技术应用于该地区(例如围封、禁牧、施肥等),但由于高寒严酷自然条件的制约,仅靠单一的禁牧封育在短期内难以恢复“黑土滩”退化草地,只有通过建植人工草地等人为措施的干扰才能实现草地的快速恢复或重建(史惠兰等,2005)。而建植人工草地需经历物种入侵、定居繁殖到群落稳定的复杂生态过程,且由于草种相对单一,生态位集中,容易造成竞争,波动性大(Britton-Simmons et al.,2008)。研究指出,在人工草地演替过程中,辅助以人工干预手段仍然需要 12年其地上植物群落生产力、多样性才能与原始植物群落相似,土壤环境12年后也仍然难以达到原始状态(Baer et al.,2002)。其中在恢复7—9年,采用多年生禾本科植物建植的人工草地植物丰富度、盖度、生物量下降,草地易发生逆向演替,出现二次退化现象(Dong et al.,2012;吴晓慧等,2019)。但也有研究表明,恢复演替8年后,人工草地植被开始达到第一个相对稳定的阶段,有较好的恢复效果,随着恢复年限的增加,植物群落和不同功能群物种丰富度和多样性开始阶段性波动,并在建植16 a左右,人工草地植物群落高度和盖度显著下降(武胜男等,2019)。实质上,人工草地的恢复演替过程是物种多样性不断变化、优势物种不断更替、植物物种生活型不断演变的过程。在该过程中,不仅地上植物群落发生显著变化,土壤养分状况(王学霞等,2012)、土壤酶活性(王启兰等,2010),以及连接植物和土壤的根系(字洪标等,2016;樊博等,2020),包括生物量、分布格局和根土比也随之发生变化。植物根系、土壤理化性质、土壤酶活性等的变化又反作用于地上植物群落,协同影响地上植物群落的变化,从而决定了人工草地的恢复演替过程(王启兰等,2009)。
目前关于人工草地生态系统长时间序列演替的规律仍存在较大争议,尤其是人工草地恢复演替过程与地下生态过程的互作机制尚不清楚。而将地上植物群落与土壤环境,特别是地下根系相结合,进行地上—地下耦合演替恢复机制的研究,才有可能准确评估和预测人工草地演替方向、动态过程和恢复的时间阈值。因为地下土壤指标驱动着地上植物群落的变化,是人工草地演替的重要驱动因子,在一定程度上决定了人工草地的演替方向(张锦华,2006)。如土壤含水量和养分供应能增加植物地下生物量并提高植物生产力(Chapin et al.,1987;Yuan et al.,2010;Chen et al.,2018);其中氮素是限制植物生长最主要的因子,氮素的增加会促进少数植物的生长,从而减少植物多样性(Stevens et al.,2004);土壤酶作为土壤的重要组成部分,对土地利用方式和管理措施的改变反应迅速,土壤酶活性在一定程度上表征了土壤微生物分解和利用土壤养分的能力(马荣荣,2015),间接影响了地上植物群落的养分供给,且酶活性也是退化草地土壤恢复的重要指标之一(杨殿林等,2020);同时,根系是土壤和植物的动态界面,是土壤资源的直接利用者和产量的重要贡献者(周本智等,2007),地下根系的吸收和转运决定了地上植物群落的生长状况,影响生态系统的演替进程(王长庭等,2012)。其中根土比(R/S)作为衡量根系在土壤中的生物量大小和空间分布规律的重要指标,其大小反映出不同退化演替阶段高寒草地生态系统地下根系和土壤“载体”量的动态变化趋势(字洪标等,2016)。因此,本研究选择了三江源区不同建植期的人工恢复草地,测定了人工草地土壤理化性质、酶活性、根土比和地上植物群落特征等参数,利用冗余分析(RDA)和Pearson相关等进行综合分析,旨在研究:(1)不同建植年限人工草地植物群落的动态演替特征及其驱动因子;(2)不同建植年限人工草地植物群落和土壤环境的协同响应。
1 研究区概况
研究区位于青海省果洛藏族自治州玛沁县,地理坐标 33°34′—34°30′N,99°54′—100°29′E,平均海拔4150 m。该区属于典型的高原大陆性气候,春季干旱多风,冬季寒冷漫长。年平均气温为-1.7 ℃,最冷月1月平均气温-14.8 ℃,最热月7月平均气温9.8 ℃。年平均降雨量600 mm,主要降水集中在5—9月,约占年降水量的 80%。土壤类型为高山草甸土,高寒嵩草草甸为该地区主要的草地类型。由于多年来的气候变化、鼠害肆虐,加之草地管理制度不健全、过度放牧等问题,导致其原生植被遭到不同程度的破坏,草地呈现出大面积退化的趋势。
2 研究方法
2.1 试验样地设计
选择青海省果洛藏族自治州玛沁县大武镇军牧场的 4个面积为 20 hm2,建植年限分别为 5 a(34°28′10′′N,100°21′01′′E,海拔 3740 m)、6 a(34°28′09′′N,100°21′03′′E,海拔 3738 m)、9 a(34°27′41′′N,100°12′35′′E,海拔 3757 m)、13 a(34°21′45′′N,100°29′58′′E,海拔 3949 m)的人工草地为试验样地,4块人工草地均为垂穗披碱草(Elymus nutans)、冷地早熟禾(Poa crymophila)、中华羊茅(Festuca sinensis)和星星草(Puccinellia tenuiflora)按照 4∶1∶0.5∶0.5(每公顷播种重量比)的比例混播,播种量为72 kg·hm-2,千粒重分别为:垂穗披碱草:2.46 g、冷地早熟禾:0.22 g、中华羊茅:0.67 g、星星草:0.61 g。播种前先清除试验地杂草,翻耕土地,精细平整,播种后覆土1—2 cm,并以150 kg·hm-2混合肥作基肥。
2.2 植物样品采集和测定
于2015年植物生长旺季(8月中旬),在4块样地内分别随机选取3个面积为50 m×50 m的平行样地,在每个平行样地内随机选择6个50 cm×50 cm的小样方进行植被群落调查。在各个小样方内记录样方总盖度及所有植物物种数。植被调查结束后,将地上植被按物种齐平地面刈割,分装进信封,带回实验室60 ℃、48 h烘干至恒质量,并称质量(Li et al.,2014)。于2015年10月进行凋落物采集,在4块样地内分别随机选取3个面积为50 m×50 m的平行样地,在每个平行样地内随机选择 6个 50 cm×50 cm的小样方进行凋落物的收集,带回实验室60 ℃、48 h烘干至恒质量,并称质量。本试验选择Shannon- Wiener多样性指数和Simpson多样性指数对植物多样性进行评价。各指标计算公式(张金屯,2004)如下:
Shannon-Wiener多样性指数:

Simpson多样性指数:

式中,Pi为种i的个体数与群落中全部物种个体数的比值;S为群落中物种数目。
2.3 土壤样品采集和测定
植被群落调查结束后,在选取的每个平行样地内用内径5 cm的土钻分层(0—10、10—20 cm)按“S”型钻取5钻土壤,均匀混合为1个土壤样品,每个样地3个重复,做好标记,放入有冰袋的采集箱中运回实验室。其中一部分样品过1 mm筛后保存于4 ℃冰箱用于酶活性及速效养分的测定,另一部分样品经风干后过0.15 mm筛用于测定土壤养分(鲍士旦,2000)14-21。
根土质量比采用土钻法(任继周,1998),在每个观测样方,用内径5 cm土钻在0—10 cm和10—20 cm土层取样,每层取5钻,重复3次,样品过筛得到土壤样品并称质量,清水冲洗得到根系样品,分别在60 ℃和105 ℃烘至恒质量,称干质量,根土质量比=根质量/土壤质量。用环刀法测定 0—10 cm和10—20 cm土壤容重(SBD)(孔凡伟,2010),每个样地重复3次。土壤含水量(SWC)测定采用烘干法;土壤pH测定采用电位法,水土比为2.5∶1;采用重铬酸钾氧化滴定法测定土壤有机质(SOM),凯氏定氮法测定全氮(TN),钼锑抗比色法测定全磷(TP),碳酸氢钠浸提-钼锑抗比色法测定速效磷(AP),原子吸收分光光度法测定全钾(TK),四苯硼钠法测定速效钾(AK)(鲍士旦,2000)22-108。
采用比色法测定土壤蔗糖酶、纤维素酶、酸性磷酸酶、脲酶、硝酸还原酶和过氧化氢酶的活性(关松萌,1986;王榛等,2018),酶活性测定均进行3次重复。
2.4 数据处理与分析
原始数据采用Excel 2007进行整理,采用IBM SPSS Statistics 21.0进行统计学分析,对土壤理化性质和植物群落指标(多样性、丰富度、地上生物量、盖度和凋落物)进行单因素方差分析,不满足方差齐性的指标先进行log函数转换,满足条件后用最小显著极差法(LSD)进行不同处理间的差异显著性检验(P<0.05),不同土层间的差异则采用独立样本t检验;用线性回归分析对土壤酶活性与建植年限进行一次、二次拟合,最佳模型以最小赤池信息值(AIC)为准;用Canoco 4.5对植物群落特征与土壤因子的相互作用关系进行排序分析,在降趋对应分析(DCA)得到第1轴梯度最大为0.144(小于3)的前提下选择冗余分析(RDA);用Pearson相关性分析土壤理化性质与植物群落特征和酶活性之间的相关性;采用Origin 8.5进行绘图。
3 结果与分析
3.1 不同建植年限人工草地植物群落特征的变化
根据研究区不同建植年限人工草地植物群落特征的变化(图 1)所示,随建植年限的增加,Simpson指数、Shannon-Wiener指数、物种丰富度先降低后增加,地上生物量、盖度和凋落物呈“N”字型变化。Simpson指数和Shannon-Wiener指数在建植6年显著低于其他年限,分别为0.85和1.32;物种丰富度、地上生物量和凋落物在建植 13年显著增加,分别为 8.67、374.63、44.09 g·m-2。
3.2 不同建植年限人工草地土壤理化性质的变化
根据研究区不同建植年限人工草地土壤理化性质的变化可知(表1),随着建植年限的增加,0—10 cm和10—20 cm土层的R/S均不断增加,在建植13年分别达到该土层最高的3.58和1.17。SWC在0—10 cm土层呈“N”字型变化,在10—20 cm土层呈“V”字型变化,其中两个土层在建植9年的SWC均为各处理中最低,分别为7.81%和7.08%。土壤pH随着建植年限的增加呈现先增加后降低的趋势,且各处理之间差异显著(P<0.05),在 0—10 cm和10—20 cm土层中均在建植13年达到最低的6.13和6.95。就土层间而言,随着土层增加,R/S显著降低(P<0.05),土壤pH显著升高(P<0.05),SWC呈降低趋势,但土层间差异不显著(P>0.05)。SBD在不同建植年限及不同土层中均无显著差异(P>0.05),变化范围在 1.35—1.49 g·cm-3。

表1 不同建植年限人工草地土壤理化性质的变化Table 1 Change of soil physicochemical properties in artificial grassland of different cultivation periods
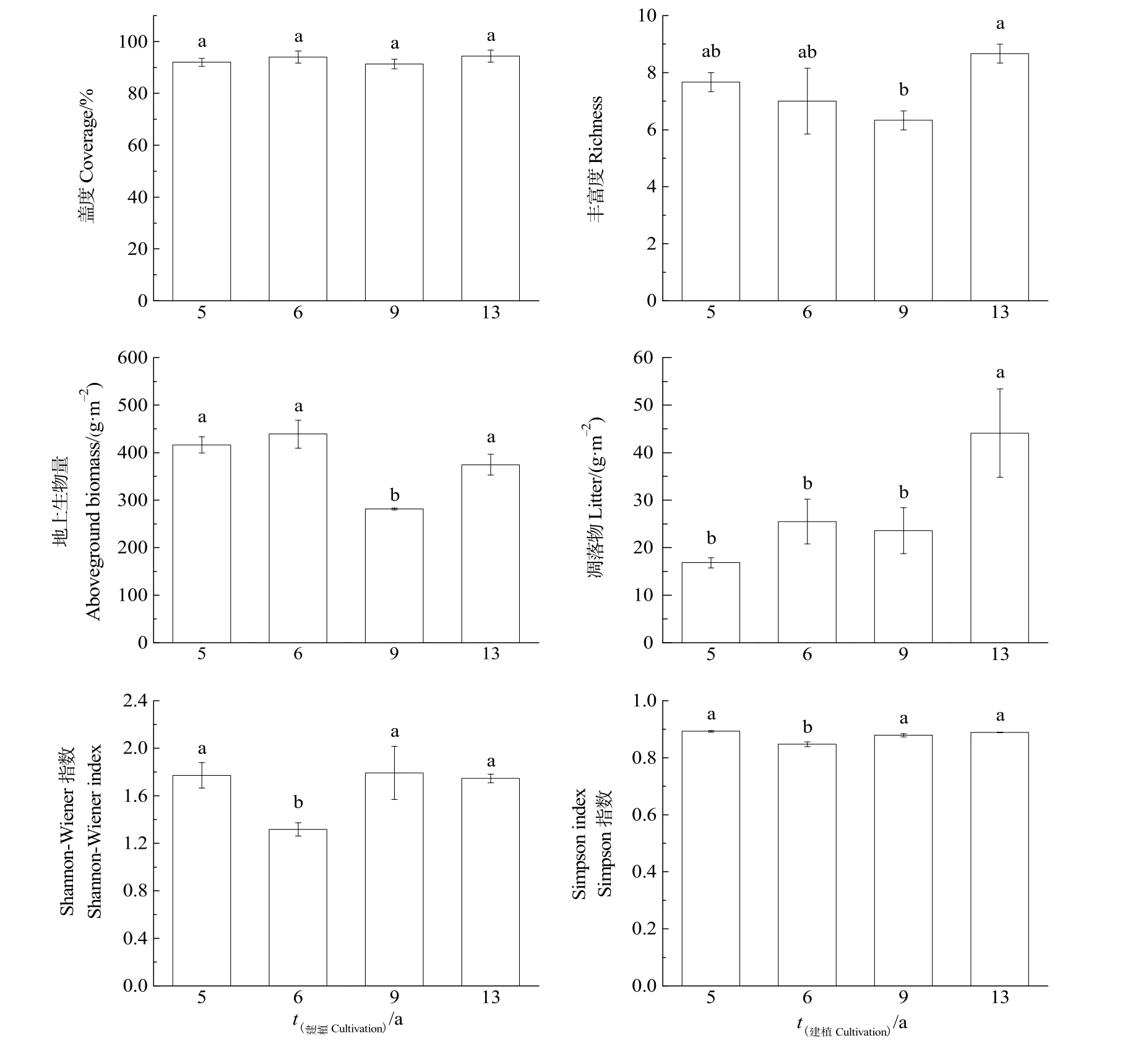
图1 不同建植年限人工草地植物群落特征的变化Fig. 1 Change of plant community characteristics in artificial grassland of different cultivation periods
在0—10 cm土层,随建植年限的增加,SOM呈“V”字型变化;土壤 TN、TP、TK、AP、AK呈“N”字型变化,且SOM、TN、TP、TK及AP均在建植9年达到最小值,分别为75.67、5.45、0.54、18.84 g·kg-1及 3.73 mg·kg-1。在 10—20 cm 土层,随建植年限的增长,SOM、TN、TK呈“V”字型变化;TP、AP呈“N”字型变化;AK不断增加。且SOM、TN、TP及AP均在建植9年达到最小值,分别为 62.47、5.09、0.63 g·kg-1及 3.08 mg·kg-1。就土层间比较而言,SOM、TN及AK随土层加深显著降低(表1)。
3.3 不同建植年限人工草地土壤酶活性的变化
对人工草地建植年限与土壤酶活性进行拟合,最优拟合函数均为二次函数(图 2,表 2)。在 0—10 cm土层,随建植年限的增加,土壤蔗糖酶、纤维素酶、酸性磷酸酶、脲酶、硝酸还原酶和过氧化氢酶活性先降低后增加,其中脲酶、硝酸还原酶和过氧化氢酶活性与建植年限有较强的相关性(R2>0.7);蔗糖酶、酸性磷酸酶、脲酶、硝酸还原酶和过氧化氢酶活性均以建植 9年最低,分别为1583.40 mg·g-1·h-1、2.33 mg·g-1·h-1、0.34 mg·g-1·h-1、0.027 μg·g-1·h-1、24.07 μg·g-1·h-1(平均值)。在 10—20 cm土层,随建植年限的增加,蔗糖酶、脲酶、硝酸还原酶和过氧化氢酶活性先降低后增加,而纤维素酶、酸性磷酸酶活性先增加后降低,其中脲酶、硝酸还原酶和过氧化氢酶活性与建植年限的相关性最强(R2>0.7);蔗糖酶、脲酶和硝酸还原酶活性以建植 9 年最低,分别为 1388.52 mg·g-1·h-1、0.36 mg·g-1·h-1、0.050 μg·g-1·h-1。

图2 土壤酶活性随建植年限增加的变化规律Fig. 2 Change of soil enzyme activities with the increase of planting years
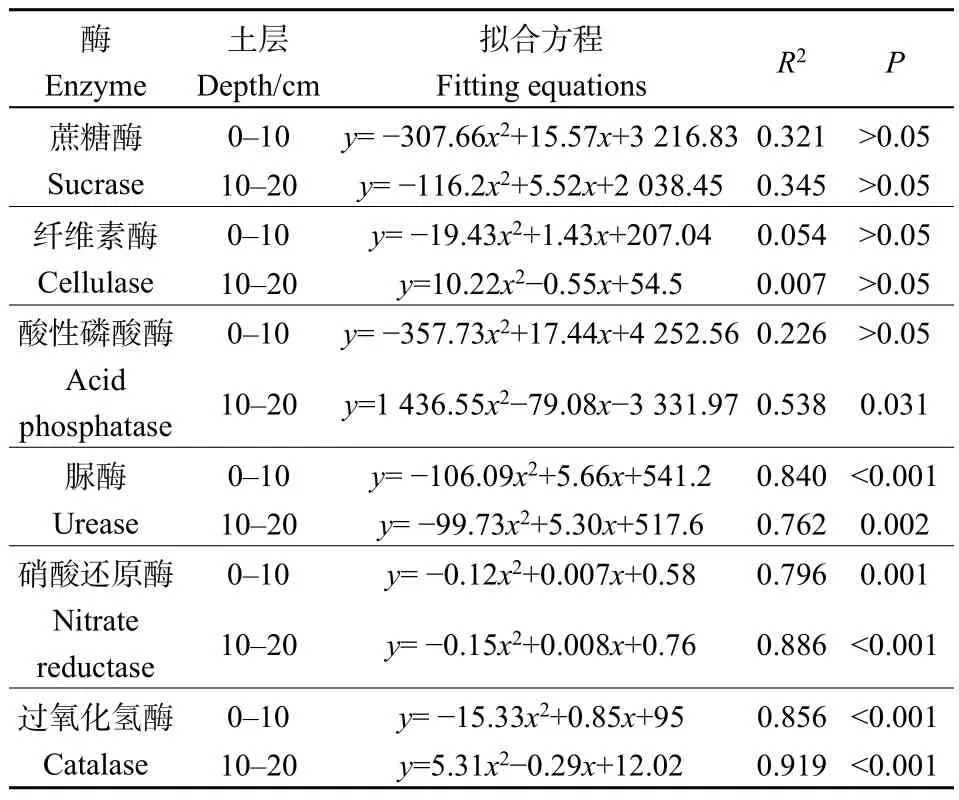
表2 土壤酶活性随建植年限变化规律的拟合结果Table 2 Fitting results of soil enzyme activities with the change of planting years
3.4 人工草地植物群落与土壤环境因子间相关性分析
3.4.1 人工草地植物群落与土壤环境因子的 RDA排序
对人工草地植物群落特征与土壤理化指标进行了RDA排序(图3),轴1和轴2分别解释总变异的63.2%和15.1%,土壤AP、AK、TP和酶活性(Enzyme activities)对人工草地植物群落的影响最显著(P<0.05)(表3)。植物生物量(Biomass)和盖度(Coverage)方向相同,表明在群落演替过程中植物生物量和盖度显著相关,R/S则与凋落物(Litter)和物种丰富度正相关。不同土壤理化性质线段长度及其与植物群落指标的夹角表明,影响植物生物量的主要因素为TN、SOM、酶活性(Enzyme activities)和SWC,影响植物盖度的主要因素为酶活性、SWC和TK,植物多样性与土壤TN含量呈负相关。

图3 不同建植年限土壤理化性质和植物群落特征的RDA分析Fig. 3 Redundancy analyses for soil physicochemical properties and plant community characteristics
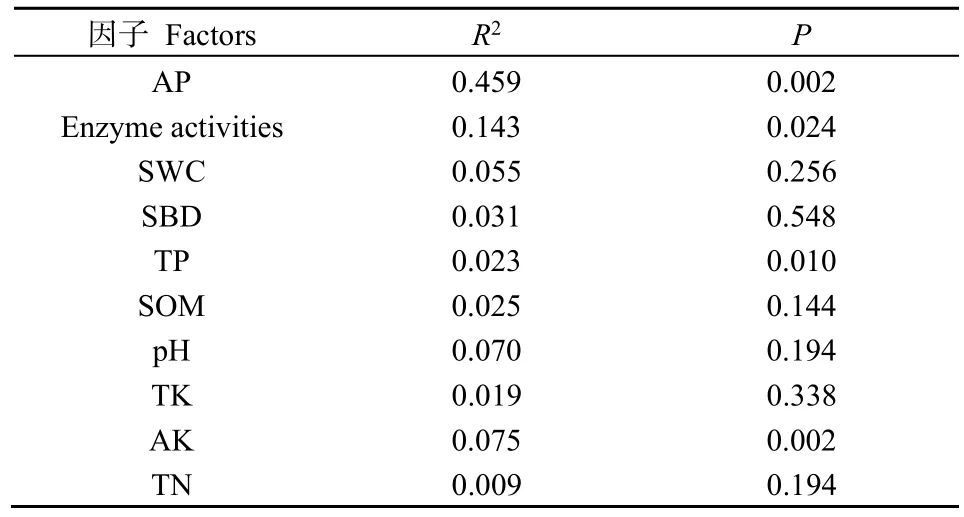
表3 冗余分析蒙特卡洛置换检验结果Table 3 Monte Carlo permutation test results of redundancy analysis
3.4.2 人工草地植物群落与土壤环境因子的相关性分析
植物群落特征与土壤理化性质的Pearson相关性表明(表4),植物生物量与土壤TN和SOM呈极显著正相关(P<0.01),与 SWC呈显著正相关(P<0.05);凋落物与土壤AP、TP和AK呈极显著正相关(P<0.01),与R/S呈显著正相关(P<0.05);植物多样性与土壤pH呈显著负相关(P<0.05)。土壤酶活性与土壤理化性质的相关性结果表明,土壤蔗糖酶活性与SOM、TN、SWC和pH呈极显著正相关(P<0.01);土壤酸性磷酸酶活性与 SOM和 pH呈显著正相关(P<0.05);土壤脲酶、硝酸还原酶和过氧化氢酶活性与土壤TN和SWC呈极显著正相关(P<0.01),土壤脲酶活性与 SOM 呈极显著正相关(P<0.01),硝酸还原酶、过氧化氢酶活性与SOM呈显著正相关(P<0.05)。
4 讨论
4.1 高寒人工草地植物群落的演替过程
时间梯度上植物群落物种多样性的变化,在一定程度上可以反映出植被的演替变化(武胜男等,2019)。本研究中,植物多样性在建植6年最低,而地上生物量在建植6年最高,建植9年最低。这与Dong et al.(2010)的研究结果相似,人工草地的建植会显著降低物种丰富度和多样性,但是能改善植被地上生物量。草地生态系统中氮素是限制植物生长最主要的因子(Mooney et al.,1987),氮素的增加会促进少数植物的生长,减少植物多样性,同时增加地上生物量(Wedin et al.,1996;Gough et al.,2000;Stevens et al.,2004)。本研究中土壤TN在建植5—6年内显著增加,提高了植物生物量。而建植9年,土壤养分显著降低,养分资源的限制导致植株个体趋向于变小(Geng et al.,2007),根据最优分配模型,植物物种将按照一定比例向根系分配更多的资源,以提高其对有限土壤养分的吸收效率(Gedroc et al.,1996)。因此,建植9年人工草地植物地上生物量和土壤养分均显著降低,但地下根系量增加。这可能是植物由有性繁殖向无性繁殖转变的表现,建植9年后,因为无性繁殖逐渐占主导,植物群落自下而上逐渐恢复,植物生物量和丰富度增加。且增加的根系提高了土壤的保水能力和养分吸收效率(樊小林等,1995),进一步促进了地上植物的生长,在建植 13年植物丰富度、多样性和生物量显著升高。

表4 植物群落特征和酶活性与土壤理化性质间的相关性Table 4 Correlation between plant community characteristics, enzyme activities and soil physicochemical property
4.2 不同建植期土壤理化性质的变化
人工草地的建植对土壤质量具有显著的改善作用,能加速碳氮循环和土壤养分的恢复过程(Ren et al.,2007)。本研究发现,土壤养分、SWC在建植6年显著增加,和Wu et al.(2010)的研究结果一致。在人工草地恢复演替初期,地上植物生物量的增加和凋落物的积累向土壤输入大量养分(Dong et al.,2010),同时植被盖度和根系的增加,减少了土壤水分蒸发(欧延升等,2019),土壤保水能力提高,SWC增加,SBD降低。随着恢复演替的进行(6—9年),由于放牧压力的增加,畜蹄的踩踏会增加土壤紧实度,从而阻碍地表水分和养分下渗(Zhao et al.,2009),归还土壤的植物凋落物减少,土壤水分和养分含量显著降低。家畜的啃食会影响地上植被的分布特征及生物量,从而间接影响土壤养分的分布(常晶晶等,2018),使土壤养分在建植9年显著降低,植物地上部分生长受阻,根系分配养分随之增加,植物根土比逐渐升高。建植9年后,由于放牧干扰导致优质牧草比例下降,毒杂草增加,从而减少了家畜的啃食(王长庭等,2009),植物开始自我调节和恢复,土壤结构和土壤肥力得以改善。与地上植物群落动态变化特征相似,土壤养分和含水量随人工草地恢复演替同样呈现“N”字型变化规律,但由于土壤容重不变,根土比呈现增加趋势,人工草地仍然处于正向恢复演替。
4.3 不同建植期土壤酶活性的变化
土壤酶类是土壤中具有生物活性的蛋白质,主要来自于土壤微生物释放、植物根系的分泌以及土壤动植物残体腐解过程的释放(王启兰等,2010;范珍珍等,2018),是反映生态系统养分循环的重要指标(Acosta et al.,2018)。土壤酶活性的变化可以用来指示高寒草甸的演替阶段和退化程度(胡雷等,2014)。本研究中,6种土壤酶活性随人工草地恢复演替均呈现“N”字型变化规律,表现为建植9年的土壤酶活性显著低于建植5、6和13年,这是由于:(1)建植 9年,地上生物量和凋落物生物量最低,输入的土壤养分,尤其是SOM含量显著降低导致酶作用底物的减少,抑制了土壤酶的活性(Acosta et al.,2007;林玉等,2019);(2)土壤含水量在建植9年也达到最低,在一定程度上降低了土壤动物和微生物活性,从而对土壤酶活性产生抑制作用;(3)土壤酶活性与土壤 pH显著相关,如土壤酸性磷酸酶受土壤pH值的影响较大(张焱华等,2007),土壤 pH越接近中性,酸性磷酸酶活性越高(黄宇等,2008)。在建植 6—9年,土壤pH从7.34降到6.25,导致0—10 cm土层土壤酸性磷酸酶活性降低。土壤深度对酶活性影响也显著,酶活性(除硝酸还原酶活性)随土层加深而降低,与多数研究结果一致(文都日乐等,2010;Ananbeh et al.,2019)。一方面,植物0—10 cm土层的根系分布显著高于10—20 cm土层,根系的分泌物是微生物丰富的能源物质(王长庭等,2009)。其次,土壤表层的凋落物和腐殖质累积较多,有机质含量高,土壤有机质作为酶合成的前体,能增加土壤中的酶源(如:土壤微生物生物量),在酶物理稳定性中具有重要作用(Acosta et al.,2007)。随着土层加深,根系分布、土壤有机质含量和微生物数量及活性的降低是导致酶活性降低的重要原因。
4.4 植物群落特征与土壤环境的协同关系
人工草地恢复演替过程中,地上植物群落、土壤理化性质和土壤酶活性均呈现相似的“N”字型动态变化特征,证明了地上植物群落和土壤环境因子之间具有协同效应,且与草地恢复演替存在明显的关联性。人工草地建植初期的施肥等人为干预措施,增加了土壤养分供给,充足的养分资源促进了植物群落的生长(Chapin et al.,1987),特别是土壤氮素的增加显著提高了植物生物量,间接促进了植被盖度和后期地表凋落物的增加,从而提高了SWC和土壤养分的有效性。随着演替的进行,由于管理不当(过牧),土壤肥力和SWC下降,土壤pH显著增加,从而抑制了生物群体的养分供应能力(Brady et al.,2002),建植7—9年植物地上生物量和丰富度显著降低。同时,土壤养分的限制改变了植物对养分资源的分配模式(Gedroc et al.,1996),加大了对根系的投资力度。且由于植物群落中各物种对于环境变化响应的对策不同,群落优势种发生改变,莎草科等通过根系分蘖进行无性繁殖的物种取代禾本科成为草地优势种。建植10—13年,人工草地正向演替的主导因素由土壤养分逐渐转变为植物根系。地下根系的增加,有利于保持土壤水分,提高根系对养分的吸收效率,并增加土壤酶源。土壤酶类通过参与碳氮磷等的循环增加了植物多样性(胡雷等,2014),保证了植物光合作用的进行和根系的发育(戴凌,2014;De et al.,2020),土壤-植物复合系统的功能得以恢复。
5 结论
(1)建植 5—6年,土壤理化性质与植物生物量、盖度及凋落物量等植物群落特征的协同演替促进了人工草地的恢复,土壤养分和水分是此阶段恢复演替的主导因子。
(2)建植 7—9年,草地土壤养分、酶活性、植物生产力和丰富度显著降低,但由于植物根系不断增加,人工草地仍处于正向演替。
(3)建植 10—13年,人工草地正向演替的主导因素由土壤养分转变为植物根系,草地也由有性繁殖主导转变为无性繁殖主导,开始自下而上的恢复,植物地上生物量和丰富度随之增加,草地向正向演替发展。
