不同光合途径植物红砂和珍珠猪毛菜幼苗对氮沉降及降水变化的光合响应
2021-01-09田艳丽种培芳陆文涛贾向阳
田艳丽,种培芳,陆文涛,贾向阳
(甘肃农业大学林学院,甘肃 兰州 730070)
全球人口的急剧增长、工农业生产迅猛发展、化石燃料的燃烧和氮肥在农田中的大量施用导致大气中排放的含氮化合物浓度迅速增加,这些氮化物通过氮沉降的形式重新进入地下生态系统,且氮输入量呈逐年上升趋势[1]。氮素是植物重要的营养元素,在植物生命活动过程中具有重要的生理调节功能,主要促进植物营养生长、提高光合能力,保证其正常生长发育[2]。全球气候变暖,也导致降水格局发生改变,局部地区降水量增加或减少[3],我国西北干旱和半干旱区在未来100年内降水将增加30~100 mm[4]。降水是植物光合作用、生长、氮循环以及植被恢复等主要生理生态过程的重要限制因素[5]。而氮和水分是影响植物光合作用和限制植物生长及生产力的两个重要因素,二者之间存在显著的耦合效应[6-7]。研究表明施氮对水分胁迫造成的光合作用的减弱具有显著的补偿作用[8],且净光合速率随施氮量的增加而增大[9]。孙群等[10]研究得出水分胁迫下随着施氮量的增加,叶绿素含量增加,并认为氮素能够增加水分胁迫下叶绿素的稳定性。可见,水分和氮素的变化对许多植物的光合特性均产生影响。在荒漠地区,水资源和氮素是限制荒漠植物生长的主要因素,水分和氮素的变化必将对其光合作用产生重要影响。因此开展氮沉降和降水变化对典型荒漠植物光合生理影响的研究,对揭示全球气候变化背景下荒漠植物响应和适应机制具有重要的意义。
高等植物分为C3,C4和景天酸代谢(Crassulacean acid metabolism,CAM)3种光合碳同化途径,相应的植物因CO2固定的最初产物不同,分别称为C3,C4和CAM植物[11]。与C3植物相比,C4植物叶片中有“花环结构”(Kranz type)[12],对CO2的吸收固定能力强,尤其是在CO2浓度低的环境下,两者的净光合速率、光呼吸速率相差更是悬殊[13]。研究表明,C3和C4植物的光合特征具有极大的可塑性[14],在特定环境条件下,植物的形态结构和生理生化功能会发生相应的改变[15],这是植物增强生存能力和竞争能力的需要[16]。氮素和降水变化对不同光合碳同化途径的植物的生长具有重要的意义。研究表明,土壤氮供应量的增加降低了C3禾本科/C4豆科作物间作体系的净产量优势,但是提高了C3禾本科/C3禾本科作物间作体系的净产量优势和补偿效应[17]。Monson等发现干旱的冬天和潮湿的夏天促进C4植物的生长,而潮湿的冬天和干旱的夏天则促进C3植物的生长[18]。因此,研究氮沉降和降水变化下C3植物和C4植物的光合特性,对探究未来气候变化对不同光合途径植物的影响具有重要意义。
在许多生态系统中,不同光合途径植物(如C3和C4)往往以混生形式共存,植物群落可通过地上光合生理调节来实现种间互惠[19]。C3植物红砂(Reaumuriasoongorica)和C4植物珍珠猪毛菜(Salaolapasserina)是干旱荒漠和荒漠草原区分布最广的植物种,在自然分布群落中,有时红砂和珍珠猪毛菜各自单独生长,有时它们紧密生长在一起[20]。红砂通过维持较高净光合速率、较高蒸腾速率和低水分利用效率来生存,属于避旱型;而珍珠猪毛菜则通过高水分利用效率来更好地生存于水资源不太丰富的荒漠,属于耐旱型,这种不同的资源利用策略是红砂和珍珠猪毛菜混生助长的机制之一[21]。张海娜[22]对红砂-珍珠猪毛菜混生群落光合特征进行研究,发现混生红砂的光合速率、蒸腾速率以及光系统II的实际光化学效率增强,而珍珠猪毛菜的Pn,Tr降低。研究表明红砂和珍珠猪毛菜混生群落适应极端环境的能力要高于单生群落,这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用[23]。有研究[24-25]指出植物间的互作类型及互作强度与其所处的环境条件之间存在着密切的关联,植物个体间相互作用的类型会随着外在环境恶劣程度的变化而产生相应的动态改变。但Maestre等[26]发现植物间互惠作用没有随着非生物因素胁迫程度的加深而增加。那么在未来氮沉降和降水变化下,不同光合碳同化途径植物红砂和珍珠猪毛菜在光合生理方面也将表现出显著差异,且因生长方式不同而有所不同。为此,本研究通过人工模拟氮沉降和降水处理,研究C3植物红砂和C4植物珍珠猪毛菜不同生长方式下的光合特性,以期进一步了解不同生长方式下荒漠植物应对环境变化的适应能力和适应机制,为深入探讨我国干旱荒漠区植物群落的更新与建设提供科学依据。
1 材料与方法
1.1 试验材料
2019年4—7月在甘肃农业大学实验基地的人工遮雨棚内进行盆栽试验,以前期培育好的同一批民勤种源1年实生苗为试验材料。2019年4月初从苗圃选取生长一致的幼苗移栽到上口径18 cm,底口径10 cm,高25.5 cm的花盆内培育,每盆栽种2株。盆内用土均取采自民勤种源地红砂-珍珠猪毛菜灌木林下0~20 cm去杂后混合均匀的均质土壤。生长方式分为:单生红砂(HH:2株红砂为1盆)、单生珍珠猪毛菜(ZZ:2株珍珠猪毛菜为1盆)和混生红砂-珍珠猪毛菜(HZ:红砂和珍珠猪毛菜各1株为1盆)。2019年5月1日选取长势一致(红砂和珍珠猪毛菜株高10 cm左右)的幼苗进行施氮和降水控制试验。
1.2 试验设计
研究结果表明,我国西北氮沉降速率在0~1.0 g·(m2·a)-1,且近10年来局部地区沉降速度达4.0 g·(m2·a)-1,大气氮沉降中的氮元素主要是以铵态氮和硝态氮形式存在,但在河西地区主要以铵态氮为主[27]。鉴于此,施氮分为N0(对照)、N1(中氮4 g·(m2·a)-1)、N2(高氮8 g·(m2·a)-1)这3种处理,氮肥由(NH4)2SO4提供,5—7月每月月初将氮肥溶于100 mL水中施入土壤,对照施入等量的水。依据民勤1961—2008年气象资料显示,该区多年平均降水量为116.7 mm,降水量较高的年份为154.2 mm左右,比多年平均水平高出约30%,降水量最低为81.5 mm,比多年平均水平低30%[28]。因而试验期间降水设置为:降水减少30%(W-)、对照(W)和降水增加30%(W+)3个梯度。以红砂生长旺盛期(5—7月)每月的降水量为基准对照,换算为各处理每月的总灌水量,分10次施入(每3天1次)。试验处理期间HH,ZZ和HZ的氮沉降和降水交互处理各9个处理(N0W-,N0W,N0W+,N1W-,N1W,N1W+,N2W-,N2W,N2W+),每个处理取6盆,共计162盆。
1.3 测定指标与方法
1.3.1光合色素含量测定 2019年7月15日早上6:00取叶片鲜样装入液氮罐迅速带回实验室并置于超低温冰箱,用于光合色素含量测定。采用乙醇丙酮浸提法[29]测定两种植物的光合色素含量,测定指标有:叶绿素a(Chlorophyll a,Chla)、叶绿素b(Chlorophyll b,Chlb)、叶绿素(a+b)(Chlorophyll (a+b),Chl(a+b))、类胡萝卜素(Carotenoid,Car)。
1.3.2光合气体交换参数测定 在2019年7月中旬晴朗的上午8:00—11:00,用LI-6400光合作用测定仪(LI-COR,Lincoln,USA)测定红砂和珍珠猪毛菜的净光合速率(Net photosynthesis rate,Pn)、气孔导度(Stomatal conductance,Gs)、胞间二氧化碳浓度(Intercellular CO2concentration,Ci)和蒸腾速率(Transpiration rate,Tr)等气体交换参数,并以Pn/Tr计算瞬时水分利用效率(Water use efficicency,WUE)[30]。
1.3.3叶绿素荧光参数测定 于2019年7月中旬使用便携式调制叶绿素荧光仪Imaging-PAM-2500(德国WALZ公司)测定叶片叶绿素荧光参数。测定时,每个处理选取健康、长势一致、光照相似的相同叶位处的叶片,先将叶片置于黑暗中适应20分钟,于早上9:00—11:00测定,每次随机选取叶片重复测定3次。测定参数包括:初始荧光(Minimal Fluorescence,Fo)、最大荧光(Maximal Fluorescence,Fm)、最大光化学效率(Maximal photochemical efficiency of PSⅡin the dark,Fv/Fm)和潜在光化学效率(Potential photochemical efficiency of PSⅡin the dark,Fv/Fo),其中Fv = Fm -Fo。
1.4 数据处理
采用Excel 2010软件对数据进行统计和制表,用统计分析软件SPSS 22进行三因素方差分析检验氮、降水、生长方式及其交互对红砂和珍珠猪毛菜幼苗的影响,利用单因素方差分析检验同一氮沉降和降雨不同生长方式下各指标的差异,不同水平间的多重比较用Duncan法,采用Pearson法对5个光合气体交换参数进行相关性分析。
2 结果与分析
2.1 两种植物光合色素对不同氮、水及生长方式的响应比较
方差分析表明(表1),氮沉降、降水变化、生长方式单因素下对红砂和珍珠猪毛菜的叶绿素a、叶绿素b、叶绿素(a+b)和类胡萝卜素均有极显著影响(P<0.01);两者交互或三者交互处理下,红砂的叶绿素a和叶绿素(a+b)呈极显著差异(P<0.01),而类胡萝卜素差异不显著,珍珠猪毛菜的叶绿素(a+b)和类胡萝卜素差异显著(P<0.05)或极显著(P<0.01)。
图1分析表明,增水和施氮交互均使不同生长方式下的红砂和珍珠猪毛菜的光合色素含量增加,且在降水增加30%和施氮量8 g·(m2·a)-1时达最大,与对照(N0 W)相比,单生红砂的叶绿素a、叶绿素b、叶绿素(a+b)、类胡萝卜素最大分别增加48%,6.03倍,1.19倍,71%;混生红砂分别增加21%,3.63倍,70%,78%;单生珍珠猪毛菜分别增加41%,79%,52%,1.19倍;混生珍珠猪毛菜最大分别增加93%,99%,95%,85%。同一氮沉降和降水处理下,红砂和珍珠猪毛菜混生显著提高了红砂的光合色素含量(P<0.05),而珍珠猪毛菜相反。与单生红砂相比,混生红砂的叶绿素a在N0W处理下增加最大,为44%,在N0W-处理下增加最小,为6%;叶绿素b在N0W+处理下增加最大,为1.04倍,在N1W处理下增加最小,为1%;叶绿素(a+b)在N0W+处理下增加最大,为47%,在N1W-下增加最小,为12%;类胡萝卜素在N1W+处理下增加最大,为31%,N0W+下增加最小,为4%。与单生珍珠猪毛菜相比,混生珍珠猪毛菜叶绿素a在N0W处理下减少最大,为30%,在N2W处理下减少最小,为3%;叶绿素b在N0W+处理下减少最大,为50%,在N1W-下减少最小,为5%;叶绿素(a+b)在N0W+处理下减少最大,为30%,在N2W+下减少最小,为7%;类胡萝卜素在N2W+下减少最大,为34%,在N2W处理下减少最小,为7%。不同处理下,红砂的光合色素含量始终高于珍珠猪毛菜。
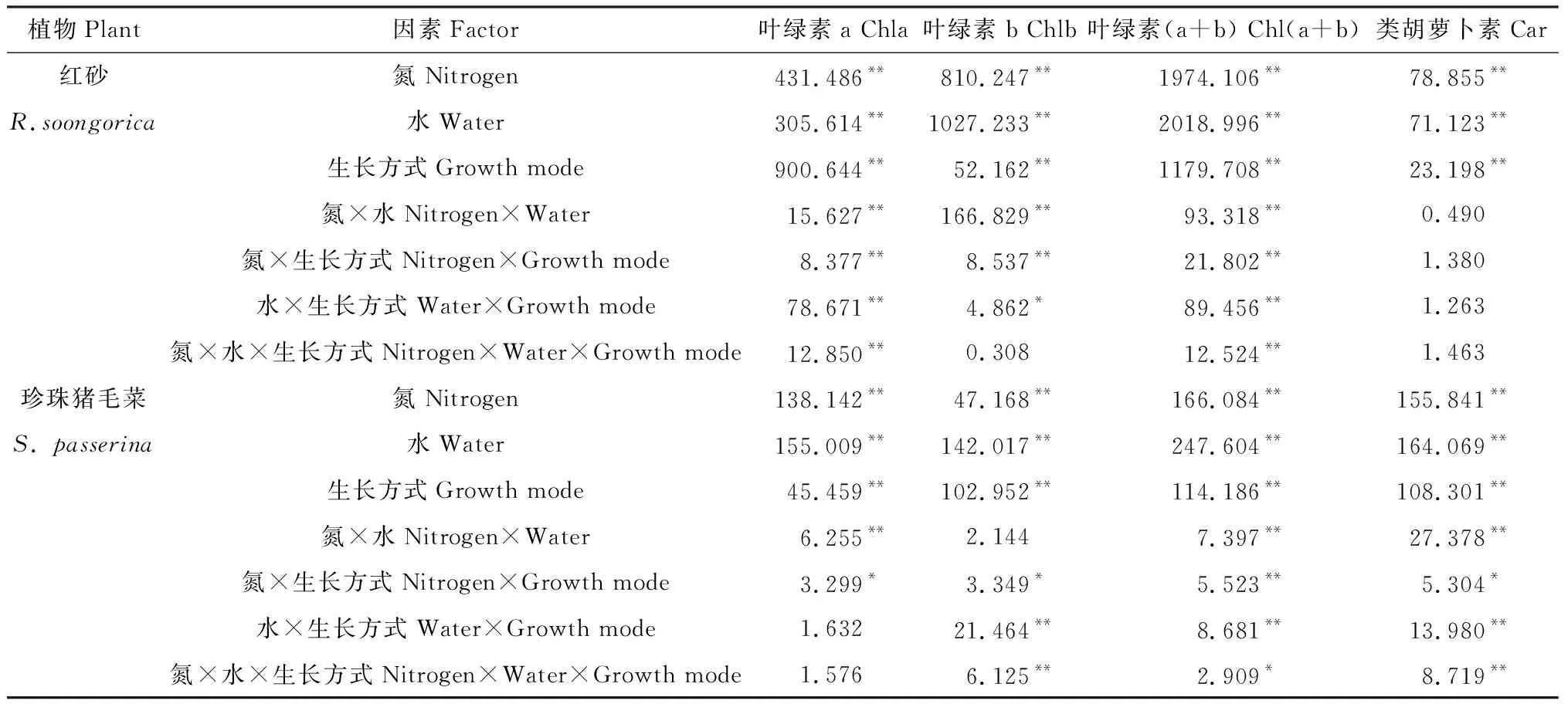
表1 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜光合色素含量影响的方差分析结果Table 1 Variance analysis results of the effects of nitrogen settlement,precipitation,growth mode and their interaction on photosynthetic pigment content in R. soongorica and S. passerina
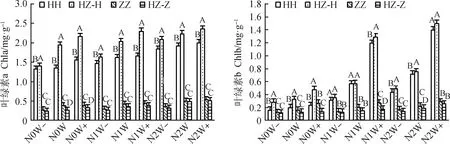
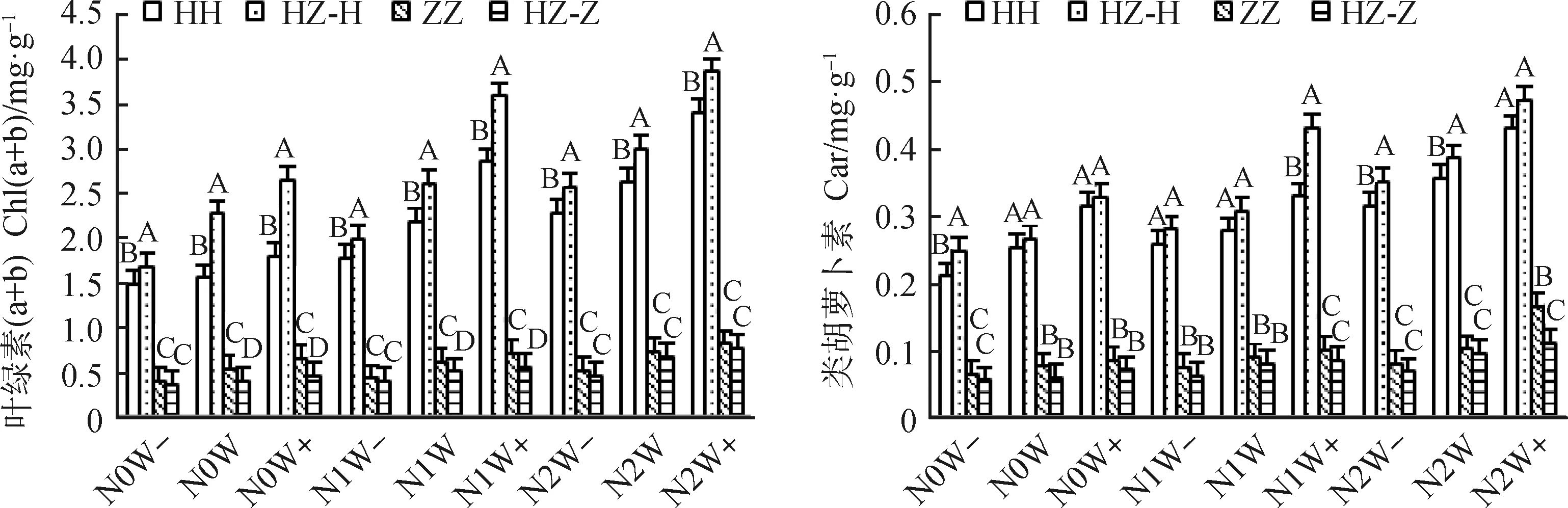
图1 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜光合色素含量的影响Fig.1 Effects of nitrogen settlement,precipitation,growth mode and their interaction on photosynthetic pigment content in R. soongorica and S. passerina注:N0,对照;N1,中氮4 g·(m2·a)-1;N2,高氮8 g·(m2·a)-1;W-,降水减少30%;W,对照;W+,降水增加30%;HH,单生红砂;ZZ,单生珍珠猪毛菜;HZ-H,混生红砂;HZ-Z,混生珍珠猪毛菜;不同大写字母表示同一施氮和降水水平下不同生长方式差异显著(P<0.05),下同Note:N0,Contrast;N1,Medium nitrogen 4 g·(m2·a)-1;N2,High nitrogen 8 g·(m2·a)-1;W-,Precipitation decrease 30%;W,Contrast;W+,Precipitation increase 30%;HH,Solitary R. soongorica;ZZ,Solitary S. passerina;HZ-H,Mixed R. soongorica;HZ-Z,Mixed S. passerina;Different capital letters indicate significant differences in different growth mode under the same nitrogen application and precipitation level at the 0.05 level,the same as below
2.2 两种植物光合气体交换参数对不同氮、水及生长方式的响应比较
表2方差分析表明,氮沉降、降水、生长方式单因素下对红砂和珍珠猪毛菜的Pn,Gs,Ci,Tr,WUE影响均极显著(P<0.01);两者交互或三者交互处理对红砂的Pn,Tr影响均极显著(P<0.01),对珍珠猪毛菜的WUE影响均不显著。
通过相关性分析结果得知(表3,4),在不同生长方式下,红砂和珍珠猪毛菜的光合气体交换参数间存在着不同程度正负相关关系。其中,单生红砂和混生红砂的Pn,Gs,Ci,Tr相互之间呈极显著正相关关系(P<0.01);单生红砂的WUE与Pn,Gs,Ci,Tr相关性均不显著;混生红砂的WUE与Tr呈极显著负相关(P<0.01)。单生和混生下,珍珠猪毛菜的Pn,Gs,Ci,Tr相互之间均呈极显著正相关关系(P<0.01);单生珍珠猪毛菜的WUE与Tr呈显著负相关(P<0.05)。
图2结果表明,不同生长方式下水氮耦合使红砂和珍珠猪毛菜的Pn,Gs,Ci,Tr值变化与光合色素变化一致,与对照相比,单生红砂分别最大增加1.19倍,83%,15%,1.04倍,混生红砂分别最大增加54%,65%,23%,39%;单生珍珠猪毛菜分别最大增加73%,1.21倍,17%,55%,混生珍珠猪毛菜分别最大增加1.13倍,1.20倍,19%,99%。而WUE值随降水减少和施氮量增加而增大,且在降水减少30%和增施氮肥8 g·(m2·a)-1交互处理时达最大,与对照相比,单生和混生红砂分别最大增加18%,25%;单生和混生珍珠猪毛菜分别最大增加34%,26%。同一氮水处理下,红砂和珍珠猪毛菜混生显著提高了红砂的Pn,Gs,Ci,Tr,WUE值(P<0.05),而珍珠猪毛菜反而减小。与单生相比,混生红砂的Pn在N0W处理下增加最大,为61%,N2W+下增加最小,为13%;Gs在N2W-处理下增加最大,为28%,N2W下增加最小,为8%;Ci在N2W+处理下增加最多,为14%,N2W-下增加最少,为4%;Tr在N0W下增加最大,为52%,N2W+下增加最小,为4%;WUE则在N0W-处理下增加最大,为16%,N0W+下增加最小,为2%。混生珍珠猪毛菜的Pn在N0W-处理下减小最多,为29%,在N2W下减小最少,为7%;Gs在N1W-下减小最多,为33%,N1W+下减小最少,为6%;Ci在N2W-处理下减小最多,为4%,在N2W+下减少最少,为1%;Tr在N0W下减少最多,为24%,在N2W+处理下减少最小,为3%;WUE则在N0W-处理下减少最大,为11%,N0W处理下减少最小,为2%。不同种处理下红砂的Pn,Gs,Ci,Tr值均大于珍珠猪毛菜,而珍珠猪毛菜的WUE略高于红砂。
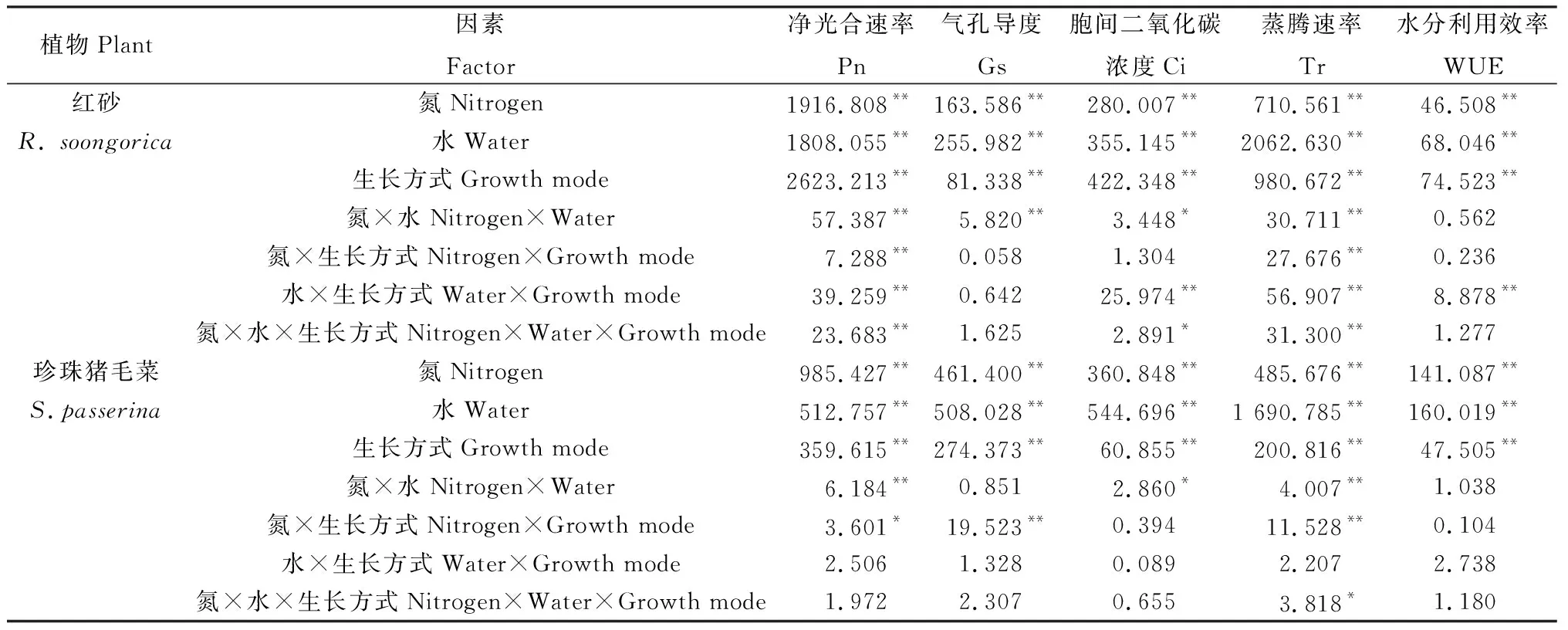
表2 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜光合气体交换参数影响的方差分析结果Table 2 Variance analysis results of the effects of nitrogen settlement,precipitation,growth mode and their interaction on the photosynthetic gas exchange parameters of R. soongorica and S. passerina

图2 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜光合气体交换参数的影响Fig.2 Effects of nitrogen settlement,precipitation,growth mode and their interaction on the photosynthetic gas exchange parameters of R. soongorica and S. passerina

表3 单生红砂(对角线上)和混生红砂(对角线下)光合气体交换参数间的相关分析Table 3 Correlation analysis of photosynthetic gas exchange parameters between solitary R. soongorica(on the diagonal) and mixed R. soongorica (under the diagonal)

表4 单生珍珠猪毛菜(对角线上)和混生珍珠猪毛菜(对角线下)光合气体交换参数间的相关分析Table 4 Correlation analysis of photosynthetic gas exchange parameters between solitary S. passerina (on the diagonal) and mixed S. passerina (under the diagonal)
2.3 两种植物叶绿素荧光参数对不同氮、水及生长方式的响应比较
表5方差分析表明,氮沉降、降水及生长方式单因素对红砂和珍珠猪毛菜的叶绿素荧光参数Fv/Fo和Fv/Fm均有极显著影响(P<0.01),氮、水、生长方式两两交互及三者交互对红砂的Fv/Fm和珍珠猪毛菜的Fv/Fo影响不显著。
图3表明,不同生长方式下,降水增加和增施氮使红砂和珍珠猪毛菜的Fv/Fo和Fv/Fm值增大,且在降水增加30%和施氮量8 g·(m2·a)-1交互处理下达最大,单生和混生红砂的Fv/Fo和Fv/Fm最大分别增加71%,11%,83%,8%,单生和混生珍珠猪毛菜最大分别增加96%,16%,1.62倍,29%。同一氮水处理下,混生红砂的Fv/Fo和Fv/Fm值显著大于单生红砂(P<0.05),而珍珠猪毛菜相反。与单生相比,混生红砂的Fv/Fo在N2W处理下增加最大,在N0W-处理下增加最小,分别增加79%,37%;Fv/Fm在N1W-处理下增加最大,在N2W+处理下增加最小,分别增加11%,7%。而混生珍珠猪毛菜的Fv/Fo,Fv/Fm分别在N1W-,N0W处理下减少最多,分别减少36%,12%;均在N2W+处理下减小最少,分别减少10%,2%。不同处理下红砂的Fv/Fo和Fv/Fm值均高于珍珠猪毛菜。

表5 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜叶绿素荧光参数影响的方差分析结果Table 5 Variance analysis results of the effects of nitrogen settlement,precipitation,growth mode and their interaction on chlorophyll fluorescence parameters of R. soongorica and S. passerina
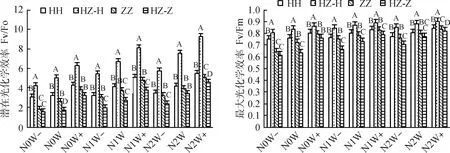
图3 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜叶绿素荧光参数的影响Fig.3 Effects of nitrogen settlement,precipitation,growth mode and their interaction on chlorophyll fluorescence parameters of R. soongorica and S. passerina
3 讨论
3.1 氮沉降、降水和生长方式对红砂和珍珠猪毛菜光合色素的影响
植物的光合作用是一个极其复杂的生理过程,不仅受自身因素影响,还受外界因素的影响[31]。氮素是叶绿素的组成成分之一,与光合作用关系密切,而水分是植物生长和进行光合作用最重要的限制因素。氮素和水分对植物的影响除彼此孤立外,还存在相互依赖的关系,且还会因研究对象及条件的不同而使研究结果存一定的差异[32]。范阳阳[33]研究发现,刺槐(Robiniapseudoacacia)幼苗叶片中的叶绿素a、叶绿素b、类胡萝卜素含量均在干旱胁迫下降低,增施氮肥后,光合色素含量显著升高。水分胁迫下,可产生大量活性氧,其直接引发叶绿体的破坏,导致叶绿素明显下降,而氮素能够增加水分胁迫下光合色素的稳定性。本研究中,增水和施氮交互均使不同生长方式下的红砂和珍珠猪毛菜的光合色素含量显著增加,说明在西北干旱地区土壤中的有机氮含量较低,植物生长受氮素严重不足和干旱的胁迫,所以在充沛的水氮环境下,红砂和珍珠猪毛菜的光合色素含量显著增加。红砂和珍珠猪毛菜混生使红砂光合色素含量增加,珍珠猪毛菜光合色素含量减少。张璐和何新华[34]研究表明C4比C3植物氮素利用效率高,所以在本试验中氮和水交互有利于减小混生珍珠猪毛菜光合色素下降幅度。说明红砂和珍珠猪毛菜混生,对二者的光合色素含量产生影响,进而影响光合作用,氮沉降和降水增加,改善环境资源状况,使种间影响减弱。本研究中,任何处理下,红砂的光合色素含量始终高于珍珠猪毛菜,说明红砂以高含量的光合色素维持高效的光合能力,从而保持对光的竞争优势,而增水施氮并没有显著改变这种情况,说明红砂和珍珠猪毛菜光合色素受自身因素影响较大,也可能是处理时间太短,还未达到影响时间。
3.2 氮沉降、降水和生长方式对红砂和珍珠猪毛菜光合气体交换参数的影响
植物的净光合速率常作为评价内外因素对其光合作用影响程度的指标[35]。气孔导度是反映气孔运动的重要生理指标,能反映出植物传导二氧化碳的能力[36]。而胞间二氧化碳浓度作为植物用于光合作用的直接原料之一,通常会随着植物的净光合速率升高而降低,且还受到植物气孔导度及大气二氧化碳浓度等因素的共同影响[31]。蒸腾速率作为植物地上部分失水快慢及能否保持植物体内水分平衡的重要生理指标对外界环境变化的反应也极其敏感[37]。氮沉降与降水耦合作用的大量研究表明[38-39],适度的水氮耦合作用将提高植物的净光合速率、气孔导度及蒸腾速率,过量则会使其降低。水氮耦合提高植物光合速率的主要原因在于充足的水氮供应明显增大植物叶片面积,进而促进其叶片蒸腾速率增强、气孔导度增大以及其胞间二氧化碳浓度的降低[40]。本研究发现,水氮耦合作用显著提高了红砂和珍珠猪毛菜的净光合速率、气孔导度及蒸腾速率,胞间二氧化碳浓度并未随净光合速率升高而降低,可能是气孔导度增大,传导二氧化碳的能力增强,从而使胞间二氧化碳浓度增加。植物水分利用效率除作为反映植物耗水与其干物质生产之间的关系外,还是评价其生长适宜程度的综合生理生态指标,且凡对植物光合与蒸腾有影响的环境因子,其对植物的水分利用效率也均有影响[41]。对小麦(Triticumaestivum)叶片瞬时水分利用效率的试验结果表明,水分胁迫导致的叶片瞬时水分利用效率的减小可以通过增施氮肥得到部分补偿[42]。本研究中水氮耦合作用使红砂和珍珠猪毛菜的瞬时水分利用效率增加,但相同施氮水平下随降水增加而减小,可能是因为降水增加导致蒸腾速率大幅度增大,从而瞬时水分利用效率反而减小。
本研究还得出不同处理下红砂的净光合速率、气孔导度、胞间二氧化碳浓度及蒸腾速率均大于珍珠猪毛菜。严巧娣等[43]的研究结果也表明红砂的光合生理参数明显优于珍珠猪毛菜。从而证明C3植物红砂积极充分地利用有效资源进行光合作用,而C4植物珍珠猪毛菜则采用保守的方式利用有效资源满足生长。而孙彦坤等[44]研究得出,C4型入侵植物反枝苋(Amaranthusretroflexus)比C3型本地作物大豆(Glycinemax)具有更高的净光合速率、气孔导度、蒸腾速率及瞬时水分利用效率,说明C3和C4植物光合优势因植物不同而有所不同,应对不同C3和C4植物做更多研究。红砂和珍珠猪毛菜混生,其光合气体交换参数变化与光合色素变化一致,这与严巧娣等[43]研究结果相同,从而证明红砂可通过种间互惠更好地利用养分资源适应种间竞争,而珍珠猪毛菜竞争作用加强,水氮的添加可有效减弱竞争强度。
从相关性分析得出,红砂和珍珠猪毛菜在单生和混生下,净光合速率、气孔导度、胞间二氧化碳、蒸腾速率相互之间均呈极显著正相关关系,而红砂的瞬时水分利用效率在单生下与净光合速率、胞间二氧化碳表现出正相关关系,混生下则为负相关关系,珍珠猪毛菜的瞬时水分利用效率在单生下与气孔导度呈正相关关系,混生下则为负相关关系,说明不同生长方式对瞬时水分利用效率与其他光合气体交换参数相关性影响较大,因此两种植物在单生和混生下的瞬时水分利用效率差异显著。
3.3 氮沉降、降水和生长方式对红砂和珍珠猪毛菜叶绿素荧光参数的影响
叶绿素荧光参数Fv/Fm和Fv/Fo是研究植物光合生理生态的重要参数,Fv/Fm是指开放的PSII反映中心捕获激发能的效率,可以用于评价PSII原初光能转换效率[45];Fv/Fo表示光反应中心PSII的潜在活性,它值的变化可以衡量光合机构是否受到损伤,表明反应中心的活性大小[46]。已有研究表明,水分和氮素影响植物叶绿素荧光参数[47],并能使旱生植物通过调节叶绿素荧光效应来适应逆境[48]。钟小莉等[49]研究也得出氮素能提高干旱胁迫下胡杨(Populuseuphratica)幼苗最大光量子产量(Fv/Fm)。本研究得出,水氮耦合作用使红砂和珍珠猪毛菜的Fv/Fo和Fv/Fm值显著增大,表明水氮耦合促进PSⅡ的光化学反应活性,通过调节内在特点,提高光合能力,增强对环境资源的利用。与光合色素及光合气体交换参数一样,红砂的叶绿素荧光参数Fv/Fm和Fv/Fo在任何处理下均高于珍珠猪毛菜,说明在环境资源充足下,珍珠猪毛菜并不能发挥C4植物的优势。苏铭等[50]试验发现,与单生相比,红砂和珍珠猪毛菜混生时,混生红砂的Fv/ Fm值和Yield值均高于单生红砂,而混生珍珠猪毛菜的Fv/ Fm值和Yield值却低于单生珍珠猪毛菜,这与本试验研究结果一致。说明红砂和珍珠猪毛菜混生对红砂生长更有利,而在氮沉降和降水增加环境下竞争作用减弱。因此,在未来大气氮沉降和降水增加环境条件下,更有利于红砂和珍珠猪毛菜混生群落的发展。
4 结论
增水和施氮能不同程度提高单生和混生下红砂和珍珠猪毛菜的光合能力。红砂在混生下光合作用增强,珍珠猪毛菜在混生下因受竞争影响光合作用减弱。增水添氮能减弱混生下红砂和珍珠猪毛菜的竞争强度,从而增强珍珠猪毛菜的光合能力,即未来氮沉降和降水增加环境变化有利于红砂和珍珠猪毛菜的生长及二者混生生长。
