不同网箱混养模式对日本黄姑鱼生长性能的影响
2020-12-21张鼎元柴学军阮泽超张燕王跃斌
张鼎元 柴学军 阮泽超 张燕 王跃斌


摘 要:研究了不同网箱混养模式对日本黄姑鱼(Nibea japonica)幼鱼生长、摄食以及网箱网衣附着情况的影响。平均体重(36.33±1.03)g的日本黄姑鱼在1 m×1 m×1 m网箱内单养27尾42 d后成活率100%,平均体重达到(85.92±2.87)g;而单养21尾日本黄姑鱼体重从(35.52±1.79)g上升到(100.10±1.90)g。本試验中梭鱼(Liza heamatocheilus)无法与日本黄姑鱼混养,三个实验组在42 d的养殖周期内梭鱼均全部死亡。日本黄姑鱼与点篮子鱼(Siganus guttatus)在网箱混养模式下有着较好的生长指标,混养27尾平均质量(36.83±0.65)g的日本黄姑鱼与18尾点篮子鱼,经过42 d日本黄姑鱼体重达到(98.32±2.90)g;而混养21尾平均质量(35.38±2.12)g的日本黄姑鱼与54尾点篮子鱼,经过42 d体重达到(120.13±2.63)g,增重率达到了(239.17±5.26)%。结果表明,在本试验条件下,与点篮子鱼混养模式下的日本黄姑鱼的存活率与单养情况下基本一致,生长及摄食情况都要优于单养模式,增重速度快,且摄食率低而饲料转化效率高。同时,优化日本黄姑鱼与点篮子鱼的混养比例可以进一步提高日本黄姑鱼的生长及摄食情况;另外,日本黄姑鱼与点篮子鱼混养情况下网箱网衣藻类附着程度也较低。
关键词:网箱混养;日本黄姑鱼(Nibea japonica);点篮子鱼(Siganus guttatus);生长;摄食
中图分类号: S917
近几十年来,我国水产养殖业发展迅速,产量不断上升,但目前我国海水鱼类养殖的主导品种多为肉食性鱼类,每年需消耗400万t左右的冰鲜鱼,这不仅对渔业幼体资源造成极大的破坏和浪费,而且由于养殖自身污染导致养殖水域环境恶化、病害频发,浒苔、赤潮等暴发频率上升,严重影响海水鱼类养殖的可持续发展。并且随着养殖集约化的发展,动物蛋白饲料日益匮乏、水域资源受限、养殖成本不断攀升等问题日趋突出。日本黄姑鱼(Nibea japonica)属鲈形目,石首鱼科,黄姑鱼属,分布于中国东海、日本南海,往北不超过北纬35°,为大型食用鱼类[1-2]。日本黄姑鱼因其肉味鲜美,营养丰富,适应不良环境和抵御疾病能力强,生长迅速,饲料系数低,是目前正在开展深水网箱养殖的优良鱼种。
梭鱼(Liza heamatocheilus)及点篮子鱼(Siganus guttatus)皆为典型的植食性鱼类,他们主要以水生植物为食,在能量转化、生态与资源方面都占据了一定的优势[3-4]。植食性海水鱼类主要以海水中的初级生产者如浮游植物、浮游动物、小型海藻等为食。在养殖中可充分利用水体食物链中的初级生产者,将初级生产者固定的能量转移到整个生态系统中,填补生态位空白,有效实现植物蛋白到优质动物蛋白的转化,既衔接了物质能量流动,又解决了动物蛋白饲料问题,其物质和能量转化优势十分突出[5]。同时,植食性海水鱼类通过与其他养殖品种的合理搭配,可优化养殖水域生态环境,构造完整生态链条,为高效养殖模式的设计与构建提供了选择。此外,植食性鱼类养殖水体具有自清洁效果,环境排放压力小,更加突出了发展该类品种养殖的环境生态优势。
本试验研究了日本黄姑鱼与梭鱼及点篮子鱼网箱混养下的生长和摄食情况,为日本黄姑鱼网箱的健康养殖提供科学的参考依据。
1 材料与方法
1.1 试验鱼与试验条件
本试验于2019年9—10月在舟山市普陀区登步岛优辰水产养殖专业合作社的网箱内进行,试验养殖网箱规格为1 m×1 m×1 m;试验用日本黄姑鱼为人工繁殖培育的子一代,平均体重为(36.33±0.43)g,体长为(14.30±0.45)cm;试验用梭鱼为海捕个体,平均体重(12.21±0.37)g。海捕梭鱼在本试验开始前已于网箱内驯养1个月以上,鱼体情况稳定;试验用点篮子鱼购买自水科院东海研究所,平均体重(12.79±0.16)g。本试验所用网箱和鱼种放养前进行消毒处理,饲养期间若发生死鱼情况,及时清理并进行无害化处理,以免传染其他鱼体。
1.2 试验方法
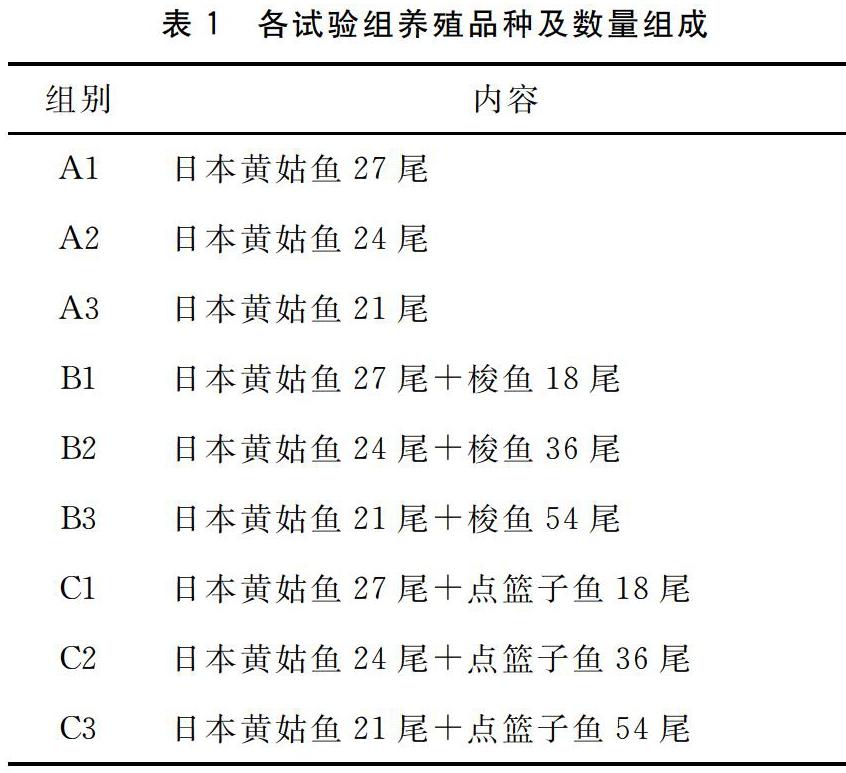
试验设9个组,每组设3个平行,试验组组成见表1。试验中所用饲料为瑞斯可海水鱼类配合饲料,每天在7:30和16:00投喂两次,每次投喂都以鱼达到饱食状态,即投入饲料后不见有鱼摄食饲料时为止。饲料主要成分分析如下:粗蛋白(492 g/kg)、粗脂肪(84 g/kg)、干物质(848 g/kg)、粗灰分(111 g/kg)和总能量(17.29 MJ/kg)。
1.3 数据测定
试验前停食1 d,然后称重(精确至0.01 g),随机将试验鱼分配至试验网箱,每个试验网箱中的鱼数量见表1。试验时间为42 d,每天记录摄食情况,每14 d检测1次生长结果。试验结束后停食1 d,然后测量体重。
1.4 数据统计
数据通过SPSS20与Excel2007进行分析统计和作图,对所得数据进行单因素方差分析(One-way ANOVA)和Duncan多重比较,以P<0.05作为差异显著水平。
参数计算依据及公式为:
2 结果
2.1 不同混养模式下日本黄姑鱼的生长与存活情况
经过42 d 的养殖后,各网箱内的鱼存活情况如表2所示,9个试验组中的日本黄姑鱼均无死亡情况,而在日本黄姑鱼与梭鱼混养情况下各试验组梭鱼皆全部死亡,另外除了C3组的点篮子鱼存活率为96.3%,C1与C2中的点篮子鱼都未出现死亡的情况。
日本黄姑鱼单养与混养条件下的生长情况情况如表3所示,1 m×1 m×1 m的网箱内单养日本黄姑鱼27尾、24尾、21尾42 d平均体重分别达到了(85.92±2.87)g、(90.23±3.16)g与(100.10±1.90)g。日本黄姑鱼的生长速度与网箱内的养殖密度逆相关,随着日本黄姑鱼养殖密度的增加其生长速度显著下降;另外,在同样的养殖密度下,混养点篮子鱼能够明显提升日本黄姑鱼的生长水平,C3组内的鱼试验结束后增重率是A3组的1.3倍。
2.2 不同混养模式下日本黄姑鱼的摄食情况
混养情况下随着日本黄姑鱼密度减少、点篮子鱼密度增加,其饲料系数有所下降,饲料转化效率和摄食率皆有显著的提高,1 m×1 m×1 m的网箱内养21尾鱼时饲料系数最低,为1.41,但是在单养以及与梭鱼混养时并无此规律。见表4。
2.3 不同混养模式下网箱网衣附着情况
日本黄姑鱼在不同混养模式下网箱网衣的附着情况如图1(见封三)所示,日本黄姑鱼在单养以及与梭鱼混养模式下部分实验组网箱网衣上有明显的藻类生长,而日本黄姑鱼在与点篮子鱼混养情况下网箱网衣上不存在这种现象,这说明点篮子鱼在与日本黄姑鱼网箱混养下可以啃食网衣上的附着藻类,对于养殖环境有着一定的改善。 3 讨论
3.1 日本黄姑鱼与点篮子鱼网箱混养具有可行性
网箱养殖是进行水产养殖活动的重要形式之一,近年来我国的网箱养殖业存在着布局不合理、养殖密度过大、大量使用鲜杂鱼投喂、影响养殖水域环境等问题,严重制约了我国水产养殖业的可持续发展[6]。基于综合水产养殖的基本原理,对网箱养殖生态系统的结构优化,将不同食性的鱼类进行混养,是平衡水质、实现养殖水体资源充分利用、扩大网箱养殖容量的有效途径[7]。本研究结果显示,梭鱼无法和日本黄姑鱼混养,梭鱼性急贪食,和日本黄姑鱼在生态位上较为重叠[3]。本试验中日本黄姑鱼体重、规格显著高于梭鱼,故梭鱼摄食受到影响,在短时间内各个网箱内梭鱼均死亡。但是点篮子鱼和日本黄姑鱼混养不仅均有良好的成活率,而且生长和摄食情况较单养情况均有提升。同时,在混养情况下随着日本黄姑鱼密度减少、点篮子鱼密度增加,日本黄姑鱼的饲料系数有所下降,饲料转化效率和摄食率皆有显著性提高。
就生态习性而言,日本黄姑鱼最适生长温度为18~28 ℃,最适盐度为30‰~35‰,pH值一般为7.5~8.5[8-9];而点篮子鱼适应温度为18~32 ℃,最适盐度为31‰~34.5‰,适宜pH值一般为7.9~8.5[10-11]。两者都属于广盐、暖水性鱼类,具有相似的养殖生态环境。此外,日本黄姑鱼为肉食性鱼类而点篮子鱼属于杂食性偏草食性鱼类,根据两者食性的差异可以对网箱养殖水体中的饲料资源进行充分利用。
网箱网衣附着藻类大量着生是困扰网箱养殖从业者的主要问题之一,大量的附着藻类会导致网箱自身的重量增大,造成网具网眼堵塞,影响网箱内外水交换[6,12],严重时会阻碍养殖对象的生长,甚至导致死亡。因此,寻找一种合理、科学、高效的附着藻类清除的方法对网箱养殖具有积极的作用。本研究结果表明,点篮子鱼对网箱网衣附着藻类具有良好的清除能力,对网箱养殖环境起到了保护和修护的作用。点篮子鱼在和日本黄姑鱼混养模式下改善了日本黄姑鱼的生存环境,提高了日本黄姑鱼的摄食效率,降低了其饲料系数。这种混养模式对海水网箱生态养殖、高效养殖、科学养殖具有重要的参考意义。
3.2 日本黄姑鱼与点篮子鱼网箱混养有利于提高综合效益
该试验中A组、B组、C组内随着日本黄姑鱼密度的降低,其增重率和平均生长率会显著升高,这说明一定程度上降低日本黄姑鱼的网箱养殖密度对于提升其生长性能是有帮助的。同时,C组日本黄姑鱼的增重率和平均生长率要高于A 组和B组,并且随着日本黄姑鱼密度的减小及混养鱼密度的增大效果更加显著,这说明按一定比例将日本黄姑鱼与点篮子鱼混养能够提高日本黄姑鱼的生长性能。其原因可能是在混养模式下,点篮子鱼与日本黄姑鱼搭配合理,两者生态位上重叠小,日本黄姑鱼属于中下层鱼类而点篮子经常活动于水体底部,这种养殖模式在空间上充分利用了养殖水体[13]。同时在能量传递上这两种鱼对不同水层的资源进行充分利用,减少了水体中的藻类、残饵等有机质,起到综合调节水质的作用,优化了养殖水域的生态环境,这与戚正梁等[14]对大黄鱼与三疣梭子蟹池塘混养的研究有着相同的结果。
篮子鱼可以摄食藻类,陆忠康[15]研究发现在菲律宾篮子鱼能够和同样是植食性的遮目鱼混养。胡成硕等[16]实验结果表明,点篮子鱼幼鱼有啃食网箱网衣附着物的特点。本试验结果也发现类似的情况,将日本黄姑鱼和植食性鱼类点篮子鱼混养对养殖网箱网衣上藻类的附着情况有着一定的改善,这与点篮子鱼的食性关系密切,其可以充分利用网箱网衣上附着的藻类资源。这体现了日本黄姑鱼和点篮子鱼混养模式对养殖资源的高效利用,有效降低了养殖成本和减少对养殖区域周边水域环境的生态压力。
参考文献:
[1] 朱元鼎,张春霖,成庆泰.东海鱼类志[M].北京: 科学出版社,1963,277-288.
[2] 张其永,洪万树.福建沿海网箱养殖鮸状黄姑鱼的鉴别[J].福建水产,1997(02):6-10.
[3] 林长虹.梭鱼的生物学特性及人工养殖技术介绍[J].科学养鱼,2004(01):19-20.
[4] 闫文罡,章龙珍,庄平,等.点篮子鱼驯养与繁殖技术总结[J].现代渔业信息,2010,25(04):30-32.
[5] 丁福红,毛守康,雷霁霖,等.我国植物食性海水鱼类养殖发展前景[J].海洋科学,2014,38(11):131-137.
[6] 刘佳英,黄硕琳.我国网箱养殖存在的问题与对策[J].中国渔业经济,2006(05):28-32+36.
[7] PARSONS G,SHUMWAY S,KUENSTNER S et al.Polyculture of sea scallops(Placopecten magellanicus) suspended form salmon cages[J].Aquaculture International,2002,10(1): 65-77.
[8] 柴学军,徐君卓,吴祖杰.日本黄姑鱼全人工繁育技术研究[J].浙江海洋学院学报(自然科学版),2007,26(02):168-172.
[9] 许源剑,孙敏,柴学军,等.日本黄姑鱼胚胎发育及温度对其过程的影响[J].浙江海洋学院学报(自然科学版),2010,29(06):544-550.
[10] 刘鉴毅,宋志明,王妤,等.温度对点篮子鱼幼鱼生长、摄食和消化酶活性的影响[J].海洋渔业,20 15,37(05):442-448.
[11] 章龙珍,罗集光,赵峰,等.盐度对点篮子鱼血清渗透压、离子含量及鳃丝Na+/K+-ATP酶活力的影响[J].海洋渔业,2015,37(05):449-456.
[12] 王建军,黄宗国,李传燕,等.厦门港网箱养殖场污损生物的研究[J].海洋学报(中文版),1996(05):93-102.
[13] 吕旭宁,蒋增杰,方建光,等.黄斑篮子鱼(Siganus oramin)對北方养殖网箱网衣附着藻类的生物清除作用[J].渔业科学进展,2017,38(5):50-56.
[14] 戚正梁,何竺柳,唐舟凯,等.大黄鱼与三疣梭子蟹池塘混养技术研究[J].安徽农业科学,2016,44(26):73-75+78.
[15] 陆忠康.篮子鱼养殖(Rabbitfish Farming)现状及其发展前景[J].现代渔业信息,1996(03):20-23.
[16] 胡成硕,刘鉴毅,赵峰,等.点篮子鱼幼鱼摄食浒苔的习性及生长特征[J].海洋渔业,2015,37(04):349-356.
(收稿日期:2020-10-21)
