水稻东野型不育系异交不亲和性的鉴定和遗传分析
2020-11-23熊涛聂元元毛方明雷建国毛凌华朱珊黄仁良沈显华严松
熊涛 聂元元 毛方明 雷建国 毛凌华 朱珊 黄仁良 沈显华,* 严松,,*
(1 江西省农业科学院 江西省超级水稻研究发展中心 南昌 330200;2 江西省农业科学院 科技管理处 南昌 330200;3 江西省农业科学院 水稻研究所 南昌 330200;#共同第一作者;*通信联系人,E-mail: yans11@163.com;shen_xh20000913@126.com)
在自然界中,自交不亲和性广泛存在,超过60%的被子植物具有这种特征,它有利于异花传粉,从而促进遗传多样性。与自交不亲和对应的异交不亲和,其存在更为广泛。异交不亲和性主要存在于异种、异属间,亲缘关系越远,不亲和性越高,即异交不亲和是物种稳定的重要保障[1]。植物异交不亲和一般发生在远缘种间,种内异交不亲和则比较罕见。从应用角度来看,发掘种内异交不亲和资源并阐明其遗传机制,颇具应用前景。将异交不亲和基因导入自然品种中,可作为不含该基因的品种(包括转基因)生产繁殖的隔离材料使用;将不亲和基因和亲和基因分别导入不育系和恢复系,而周边种植不含亲和基因的材料,则可实现杂交种制种的生物学隔离。然而在杂交稻育种中,种内的异交不亲和却是影响繁种、制种产量以及生产成本最大的因素之一。
目前,在植物自交不亲和性研究方面已取得一系列重要进展[2-4],而对于异交不亲和性的研究还比较薄弱,主要因为异交不亲和性的遗传研究相对较难。如果不亲和在两个亲本中是双向的,就不能产生后代进行遗传研究,只有单向方可进行[5]。
在禾本科作物中,已在玉米和水稻上发现有单向异交不亲和现象。玉米上发现异交不亲和可以追溯到一百多年前,1902 年科学家Correns 发现第一个异交不亲和基因Ga1[6],1936 年Burnham 报道了Ga2[7],之后Ga3~Ga10及Tcb1等多个单向异交不亲和位点陆续被发现[5,8],目前仅Ga1被克隆报道。Zhang等对Ga1位点中的雄性控制基因ZmGa1P进行克隆和功能验证,发现ZmGa1P编码一个在Ga1-S和Ga1-M型玉米自交系花药中特异表达的果胶甲酯酶,ZmGa1P位于花粉管顶端,与另一个花粉管特异表达的 PME 蛋白互作,共同维持花粉管正常的甲酯化修饰程度,以保障花粉管在Ga1-S型花丝中的正常伸长,最终受精结实[9]。
作为自花授粉的水稻,异交不亲和主要发生在远缘野生种和栽培种以及不同染色体组的野生种之间,这也是生殖隔离的一种表现。2003 年Matsubara 等[10]报道了野生稻和栽培稻异交不亲和的遗传现象,通过正反交试验发现单向异交不亲和由来自母本 W593A 的Cif基因和来自父本 T65wx的cim互作产生,与玉米相似,Cif花粉能给隐性纯合的cim母本授粉结实,而cim花粉不能给显性纯合的Cif母本授粉结实,两者都被定位在第6 染色体上,分别位于短臂和着丝粒附近。2008 年,他们进一步研究发现Cif其实存在两个不连锁的位点Cif1和Cif2,都在第6 染色体上,Cif1被定位在标记R2291 和 RM204 之间 0.67 Mb 的区段上,Cif2定位在RG264 和RM3498 之间,两标记相距约18 cM,涵盖着丝粒区域,与Cim位置相近[11]。可能因分解难度大,之后未有新进展报道。
我们在东乡野生稻利用过程中,也发现了异交不亲和现象。20 世纪80 年代,江西省农业科学院水稻研究所以东乡野生稻为细胞质源,采用野栽交和核置换的方法,培育出具有全新恢保关系的东野型雄性不育系[12],之后通过多年大量测交试验,直至近年实现三系配套[13]。然而,在东野型不育系培育过程中,发现部分保持系品种与不育系存在异交不亲和现象,这些品种在杂交或回交时,不育株系的异交结实率很低,甚至为零而不能继续转育。针对该现象开展了初步研究,通过构建不同核质背景的材料,探索东野型不育系异交不亲和性的形成原因,进而对该性状进行遗传分析和基因初步定位。
1 材料与方法
1.1 材料
以东野型不育系DY1A及其保持系早籼稻中早35[13]为主要材料。其余水稻材料有粳稻品种02428、秀水 123,籼稻野败型保持系Ⅱ-32B、金 23B、五丰B,恢复系华占、R668、广恢308 和93-11。
利用染色体单片段代换系研究异交不亲和性,该单片段代换系由 Ando 等[14]构建,以粳稻品种Sasanishiki 为轮回受体、籼稻品种Habataki 为供体,共有 39 个系[14]。
1.2 田间试验
2014−2019 年,在江西省农业科学院的江西南昌和海南三亚两处试验基地种植,每个材料种植 6行,每行8 株,株间距为15 cm×20 cm,田间常规水肥管理。
在不育系盛花期,傍晚将不育系颖壳剪去 1/3至 1/2,保留完整雌蕊,次日取父本花粉对它充分授粉,套袋防止串粉。对同一组杂交,每个父本做3 个套袋,每个杂交套袋用2~3 穗不育系。
1.3 性状考查
授粉后15~25 d,考查结实率。结实率(%)=实粒数/总粒数×100,取各穗结实率的平均值,利用Excel 2016 进行数据处理与分析。
1.4 基因定位分析
根据代换系群体遗传图谱,结合表型数据对基因进行定位,遗传距离用centiMorgan(cM)表示。
2 结果与分析
2.1 东野型不育系异交不亲和性的产生
我们通过分别构建同质异核系和同核异质系,初步探讨东野型不育系异交不亲和性形成的原因。
首先,我们进行了同质异核系杂交试验,发现所使用的东野型不育系DY1A,在与保持系中早35回交转育过程中未发现不亲和性。以DY1A 为胞质供体,利用 3 个粳稻(Sasanishiki、02428 和秀水 123)以及 8 个籼稻(Habataki、广恢 308、R668、93-11、华占、II-32B、金23B、五丰B)为轮回父本进行不育系转育。在杂交当代,Sasanishiki 对DY1A 表现显著的不亲和性,F0结实率仅1.5%,其余10 份材料与 DY1A 杂交结实正常(>75%)。值得一提的是,东野型败育胞质具有广保性,所有F1植株的花粉育性和结实率均为0。接下来,在回交第1 代,02428、秀水123、华占、93-11、广恢308、金23B和五丰B 这7 份材料对DY1A 的杂交结实率显著下降(回交时对每个父本只做了3 个套袋的回交,所选择的回交单株数不大于 3,各回交世代仅计算平均结实率,未能统计分离比);回交第2 代时,它们的杂交结实率继续下降。只有Habataki、R668 和Ⅱ-32B 这3 份材料能够正常转育,各回交世代结实正常(表1)。由此得知:1)东野型不育系存在异交不亲和现象;2)东野型不育系的异交亲和性受核遗传背景影响。
我们也进行了同核异质系杂交试验。在上述回交转育过程中,金23B 和五丰B 与DY1A 在BC2F0的结实率分别为5.3%和17.7%,进一步回交,BC3F0的结实率分别为4.5%和2.2%(表1),表现典型的异交不亲和性。而含有野败型败育胞质的同核异质系金23A 和五丰A 与各自保持系杂交结实正常,表明东野型不育系的异交不亲和性存在细胞质效应。
此外,针对上述Sasanishiki 与DY1A 在杂交当代不亲和,也进行了同核异质杂交试验,仍以Sasanishiki 为父本,DY1A 的异质保持系中早35 作母本,结果它们杂交结实正常,可见两者的细胞核是亲和的,进一步说明东野型不育系的细胞质影响了异交亲和性。
综合以上结果,明确了东野型不育系的异交不亲和性由核质互作产生。
2.2 东野型不育系杂交不亲和类型
根据上述杂交试验发现,东野型不育系的异交不亲和性存在两种类型,第一种类型(Ⅰ)发生在杂交当代,以Sasanishiki 为代表。第二种类型(Ⅱ)发生在回交世代,代表品种有 02428、华占等,在BC1F0时它们就表现出部分不亲和,且观察到结实率有分离,随着回交世代增加,不亲和性逐渐凸显。
2.3 利用单片段代换系研究异交不亲和性Ⅰ
将 Sasanishiki 和 Habataki 与 DY1A 杂交,两者的亲和性截然相反,前者不亲和,后者亲和。进而将它们衍生的一套染色体单片段代换系SL401-SL439 分别与DY1A 杂交,统计结实率。结果,筛选到2 个杂交亲和的代换系(SL426 和SL427),杂交结实率均在80%以上,完全正常(图1)。其余代换系与DY1A 杂交结实率均不足10%,大部分代换系杂交几乎不结实。表明Habataki 与DY1A 杂交的亲和性有主效因子作用,且位于 SL426 和SL427 所携带的代换片段上。
2.4 异交不亲和性Ⅰ的遗传方式
利用亲和代换系 SL426 与 Sasanishiki 的 F1及与Sasanishiki 回交的BC1F1分别与DY1A 杂交来分析异交不亲和性Ⅰ的遗传行为,结果,F1与DY1A杂交结实率为35.9%,表现中间型。BC1F1共35 株与DY1A 杂交,结实率表现为差(<5%)和中间型(30%~40%)两种情况,差和中间型两种表型株数分别是17 和18 株,符合1∶1 分离,初步推测该类型的异交不亲和性Ⅰ在细胞核内受单隐性基因控制,暂命名为CI1(t)。F1与 DY1A 杂交的结实率为中间型,说明CI1(t)的作用可能存在剂量效应。

表1 东野型不育系DY1A与11个品种杂交/回交的结实情况Table 1. Seed setting rates of the DW-type CMS DY1A crossing/backcrossing with 11 varieties.

图 1 DY1A 分别与 Sasanishiki、SL426 和 SL427 杂交的结实情况Fig. 1. Seed setting performances of DY1A crossing with Sasanishiki, SL426 and SL427, respectively.
2.5 初步定位CI1(t)
根据染色体代换系的遗传图谱[14],第8 染色体有4 个单片段代换系,依次为SL425、SL426、SL427和SL428,SL425 与后三者不重叠,SL427 与前后相邻的代换系SL426和SL428均有重叠片段。其中,SL426 含从 RM6838 到 RM3395 的片段,SL427 覆盖RM3395-RM5891,两者在RM3395 所在位置重叠;SL428 覆盖 RM5767-RM4997,与 SL427 除RM3395 以外的片段重叠。此外,在其他染色体上,SL426 还含有第 5 染色体 RM6517、SL427 含有第 6染色体RM5957、SL428 含有第2 染色体RM3316和第 3 染色体 RM5442 的小片段,这些片段来自Habataki。第 2、3、5 和 6 染色体的代换系以及 SL428与 DY1A 杂交,结实率表型都与轮回亲本Sasanishiki 相似,几乎不结实,仅 SL426、SL427及供体亲本Habataki 三者与DY1A 杂交结实正常,因此,将CI1(t)初步定位在 SL426 和SL427 的重叠区域,位于RM6838 和RM5767 两个标记之间,与RM3395 连锁(图2),前后两标记相距18.3 cM,物理距离约12 Mb,为精细定位奠定了基础。
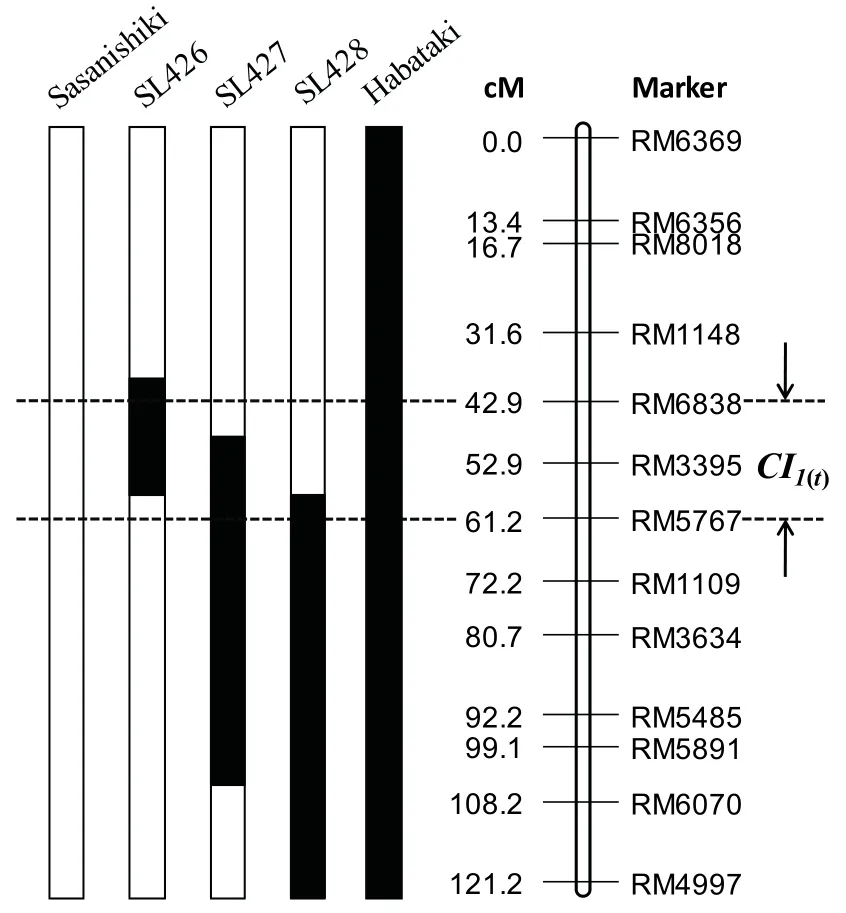
图2 CI1(t)在第8 染色体上的初步定位Fig. 2. Rough mapping of of CI1(t) on chromosome 8.
3 讨论
Matsubara 等[10]报道的水稻单向异交不亲和仅受核基因控制,授粉后种子发育退化或皱缩,本研究中的表型与它有所区别,东野型不育系的异交不亲和表现为结实率低或不结实,而且是一种新类型——由核质基因互作产生。我们熟知的植物远缘杂交不亲和,可能有一些就是由核质互作产生的。
此前我们利用分子标记通过对东乡野生稻居群检测发现,只有部分株系含有东野型败育细胞质[15],因而推测东野型不育系存在的异交不亲和性,可能是含此类细胞质的东乡野生稻血缘的一种自我保护机制,即这些野生稻株系拥有特定的细胞质基因,其他资源与它们杂交,需要带有与这类细胞质信号相匹配的亲和核基因,才能实现基因交流。
相比籼稻,我们发现粳稻在与东野型不育系杂交/回交的较早世代就表现不亲和性。在东野型三系杂交稻育种过程中,我们选用的粳稻品种(>50 份)转育东野型不育系最终几乎全部表现不亲和,而籼稻却有少数品种完全亲和,表明东野型三系体系不适用于粳稻,而在籼稻上距离生产应用仍有很长的路要走,当务之急是解决东野型不育系异交不亲和性的瓶颈问题。
本研究发现的两种不亲和性发生在不同杂交世代,说明它们各自的控制基因可能不同。第一种类型在杂交当代出现,受单隐性基因控制,并利用单片段代换系群体将其定位在第8 染色体SSR 标记RM6838 和RM5767 之间,覆盖着丝粒区域,对后续精细定位有一定难度。第二种类型的不亲和性在回交一代时初步体现,回交至第二或第三代时彻底显现,推测它的遗传方式也不复杂,在育种应用中对它们进行遗传改良应是可行的。
目前对水稻异交不亲和性的研究比较滞后,主要有两方面原因。一是主观因素,科研人员对这类性状认识不足,即便遇到也可能误以为杂交没有做好而忽视,因此相关研究报道非常少。众所周知,手工杂交的结实率比不上自交。本研究中,DY1A与不同品种杂交、回交的结实率变幅很大(0%~88.6%),其中不乏手工杂交没有做好的情况。实际上,影响杂交结实率的因素较多,比如授粉时的花粉数量和质量等,所以在后续基因定位过程中,将杂交做好从而准确鉴定表型十分关键。另一方面是客观原因,异交不亲和不同于其他农艺性状,它需要通过杂交试验来判断表型,比对一般农艺性状的观测多一道程序,若当代杂交失败就意味着要多一个世代来确定表型,而且对于纯合与杂合表型区分不开的情况还需要利用后代家系去鉴定。不管怎样,挖掘异交不亲和材料加以研究,无论对理论还是应用研究都很有价值。
