谷子bZIP转录因子家族的全基因组鉴定与生物信息学分析
2020-09-14卢平武懿茂武强强李雪垠
卢平,武懿茂,武强强,李雪垠
(1.山西农业大学生命科学学院,山西太谷030801;2.山西农业大学农学院,山西太谷030801)
谷子(Setaria italica)属于1年生草本植物,为二倍体禾本科作物(2n=18),起源于我国,具有悠久的栽培历史,我国的黄河中上游为主要栽培区。谷子是C4植物,耐旱、耐贫瘠,是发掘农作物抗旱基因和研究作物抗旱分子机制的优良种质资源[1]。2012年谷子的基因组测序已经完成,为谷子功能基因组学研究提供了数据平台[2-3]。目前,谷子逐渐成为理想的模式作物之一,主要应用于禾本科植物的基因组学研究。谷子脱壳后称为小米,小米富含蛋白质、脂肪、铁、类胡萝卜素、纤维素和维生素等,其营养价值很高;还可作为药物[4],具有清热、滋阴、治水泻等功效;又可以用来酿酒。谷子的茎叶又可以用作牲畜的优等饲料。因此,其越来越受到人们的重视。
转录因子能调控下游基因的时空特异性表达,是真核生物中重要的调控蛋白。在植物中,转录因子是调节生长发育和胁迫应答过程的关键因子。碱性亮氨酸拉链(Basic leucine zipper,bZIP)转录因子广泛分布于真核生物基因组,是数量最大、最保守的基因家族之一,在植物的生长发育、病原防御、高温以干旱胁迫应答等生物和非生物胁迫反应中起着重要的调控作用[5]。因此,bZIP转录因子的基因鉴定和功能分析等研究在后基因组时代中是较为活跃的研究领域。bZIP转录因子是以共有的bZIP保守结构域为依据而被命名的。bZIP结构域包括一个C端的碱性氨基酸区域和一个N端的亮氨酸拉链区域,bZIP保守结构域包含了60~80个氨基酸,其中,碱性区域含有一个保守基序N-x7-R/K-x9,该保守基序由16~20个氨基酸残基组成,这个区域参与细胞核的定位与DNA识别[6-7];亮氨酸拉链结构域由若干个两亲α螺旋构成,在α螺旋中每隔6个氨基酸就出现一个亮氨酸或其他疏水性的氨基酸,例如缬氨酸、甲硫氨酸、异亮氨酸和苯丙氨酸。
在植物中,bZIP转录因子调节包括病原防御、光反应和胁迫信号、种子成熟和花发育等过程[8]。在很多植物中,bZIP转录因子家族成员已经被鉴定或预测完成。在模式植物拟南芥和水稻中,分别包含75个和89个bZIP家族的成员[9-10]。在拟南芥中,bZIP转录因子能够通过ABA途径调节植物生长发育和非生物胁迫等过程[11];在水稻中,bZIP转录因子与干旱胁迫和花粉发育相关[12-13];在小麦中,有48个bZIP基因与花药发育有关[14];在辣椒中,bZIP转录因子能够抵御病原感染和非生物胁迫[15]。迄今为止,bZIP转录因子在拟南芥和水稻中的抗逆性研究较为深入,但在谷子中的研究却相对较少[16]。
本研究利用生物信息学方法对谷子bZIP转录因子进行全基因组鉴定,并分析其家族成员的系统进化关系、基因结构、蛋白质理化性质、保守基序、启动子顺式作用元件和表达量水平,旨在为进一步研究谷子bZIP基因功能和分子机制奠定理论基础。
1 材料和方法
1.1 全基因组鉴定和蛋白质理化性质预测
根据Pfam(http://pfam.xfam.org/)数据库中谷子bZIP转录因子的Pfam编号,在Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)v12数据库中筛选出85个bZIP转录因子基因,并下载它们的基因组序列、编码区序列(Coding sequence,CDS)和蛋白质序列。根据bZIP转录因子在染色体上的位置命名,谷子bZIP转录因子基因的染色体定位和基因组位置信息来源于Phytozome。利用ExPASy-Compute pI/Mw(https://web.expasy.org/compute_pi/)网站计算谷子bZIP转录因子蛋白质的理论等电点和分子量。
1.2 系统进化和基因结构预测
利用MEGA 7.0软件对85条谷子bZIP转录因子的氨基酸序列进行多序列比对(Clustal W);采用Neighbor-Joining法构建系统发育树,参数Bootstrap值设为1 000,其余参数默认。利用Gene Structure Display Server 2.0(http://gsds.cbi.pku.edu.cn/)网 站预测谷子bZIP转录因子基因结构,包括外显子、内含子和上下游区域。
1.3 蛋白质模体和基因启动子顺式作用元件预测
利用网站MEME(http://meme-suite.org/)上传氨基酸序列,分析谷子bZIP转录因子的蛋白质模体,模体数目设置为15个。
利用Phytozome网站的数据库Setaria italica v2.2下载bZIP转录因子基因的5′UTR上游1.5 kb序列(包括5′UTR)作为谷子bZIP转录因子基因启动子序列。利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)网站在线预测谷子bZIP转录因子基因启动子上的顺式作用元件。
1.4 基因组织特异性和环境诱导表达预测
利用TBtools,根据谷子bZIP转录因子基因在不同组织中和不同环境中的表达数据绘制热图。谷子bZIP转录因子基因不同组织、不同条件下的表达量数据来源于Phytozome Setaria italica v2.2数据库。
2 结果与分析
2.1 全基因组鉴定及其蛋白质基本理化性质分析
表1对谷子bZIP转录因子的基因编号、染色体定位、基因组位置、基因长度、CDS长度、氨基酸个数、蛋白质的理论等电点和分子量进行了统计,利用Phytozome网站在谷子全基因组水平鉴定了bZIP转录因子基因共85个,根据bZIP转录因子在染色体和基因组位置上的分布,按顺序依次命名为SibZIP1~SibZIP85。谷子bZIP转录因子基因在1~9号染色体上的数量分别是10、15、13、10、12、7、4、3、11个,分布不均匀。

表1谷子bZIP转录因子基因基本信息及其蛋白质基本理化性质

续表1
在谷子bZIP转录因子的蛋白质中,氨基酸个数最多的蛋白质是SibZIP35,有645个;最少的是SibZIP52,有140个。谷子bZIP转录因子家族蛋白质的理论等电点介于4.69~11.97,分子量在15.507~68.096 ku。蛋白质等电点偏高通常与其较高的碱性氨基酸(如精氨酸、赖氨酸、组氨酸)含量有关。例如SibZIP38、SibZIP61、SibZIP78、SibZIP83蛋白质等电点较高,分别为11.41、11.97、11.03、10.17,因为这几个SibZIP转录因子蛋白质有较多的碱性氨基酸,因此等电点也较高。
2.2 系统进化树的构建和基因结构分析
对谷子85条bZIP转录因子的氨基酸序列进行多序列比对,并构建系统进化树如图1(左)所示,部分SibZIP转录因子家族成员亲缘关系较为相近,如SibZIP19/SibZIP68、SibZIP85/SibZIP79、SibZIP74/SibZIP70、SibZIP23/SibZIP80、SibZIP1/SibZIP51、SibZIP35/SibZIP22、SibZIP4/SibZIP44、SibZIP33/SibZIP60、SibZIP34/SibZIP48、SibZIP74/SibZIP70。因此,它们可能是旁系同源基因,由同一个bZIP转录因子祖先基因复制产生。图1(右)为谷子bZIP转录因子基因的外显子-内含子的结构情况分析。由图1可知,85个谷子bZIP转录因子中亲缘关系较为相近的基因结构也较为相似,如SibZIP19/SibZIP68、SibZIP85/SibZIP79、SibZIP74/SibZIP70、SibZIP23/SibZIP80,每组外显子的个数都相等、长度相似,说明亲缘关系较近的SibZIP基因的基因结构具有较强的保守性。
2.3 蛋白质模体和启动子顺式作用元件分析
利用网站MEME分析谷子bZIP转录因子蛋白质模体,结果如图2所示,谷子bZIP转录因子蛋白质预测了15个模体,分别为模体motif1~motif15;85个bZIP蛋白质都含有motif1,说明motif1在谷子bZIP蛋白质中分布最广,保守性最强;motif1有41个氨基酸,其中,5、10、17、19位的R(精氨酸)和27、34、41位的L(亮氨酸)具有较强的保守性,符合bZIP碱性亮氨酸拉链的特征。
部分bZIP转录因子蛋白质模体差异较大,一些亲缘关系较近的bZIP转录因子蛋白质模体也较为相似。如SibZIP23和SibZIP80都具有motif1、motif4和motif12这3个motif;SibZIP2和SibZIP50、SibZIP4和SibZIP44、SibZIP34和SibZIP48都仅有motif1;SibZIP25和SibZIP81、SibZIP57和SibZIP31都具有motif1、motif8、motif10、motif11。
bZIP转录因子在植物中参与生长发育、光的形态建成和防御与胁迫应答等生理过程。为了探究SibZIP转录因子基因启动子区域中顺式作用元件与谷子生理过程的关系,本研究利用PlantCARE网站分析85个bZIP基因的启动子顺式作用元件,部分结果如表2所示,其中,AuxRR-core、TGA-element和P-box元件功能涉及植物生长发育过程的激素调控;G-box、GT1-motif和Sp1元件功能涉及植物光反应调控;ABRE元件功能涉及植物ABA响应。ABA不仅能够抑制植物生长,还能够在干旱、寒冷、高温、盐渍和水涝等逆境中增强植物抗逆性。MBS元件是MYB结合位点,其功能涉及植物干旱诱导;TC-rich repeats元件功能涉及植物防御和应激反应;ABRE、MBS和TC-rich repeats元件涉及防御与胁迫应答。
SibZIP78基因启动子中含有16个G-box元件;SibZIP75基因启动子中含有11个G-box元件和1个Sp1元件;SibZIP78、SibZIP54基因启动子区域内含有较多的光响应相关顺式作用元件;SibZIP54基因启动子中含有1个AuxRR-core元件和3个TGA-element元件,SibZIP79基因启动子中含有1个AuxRR-core元件、1个P-box元件和1个TGA-element元件,SibZIP54、SibZIP79基因启动子富含激素调控相关的顺式作用元件;SibZIP78基因启动子中含有15个ABRE元件,SibZIP71基因启动子中含有14个ABRE元件,SibZIP2基因启动子中含有12个ABRE元件,SibZIP73基因启动子中含有9个ABRE元件和2个MBS元件,这4个基因启动子富含ABRE元件或MBS元件等与胁迫应答相关的顺式作用元件(表2、3)。
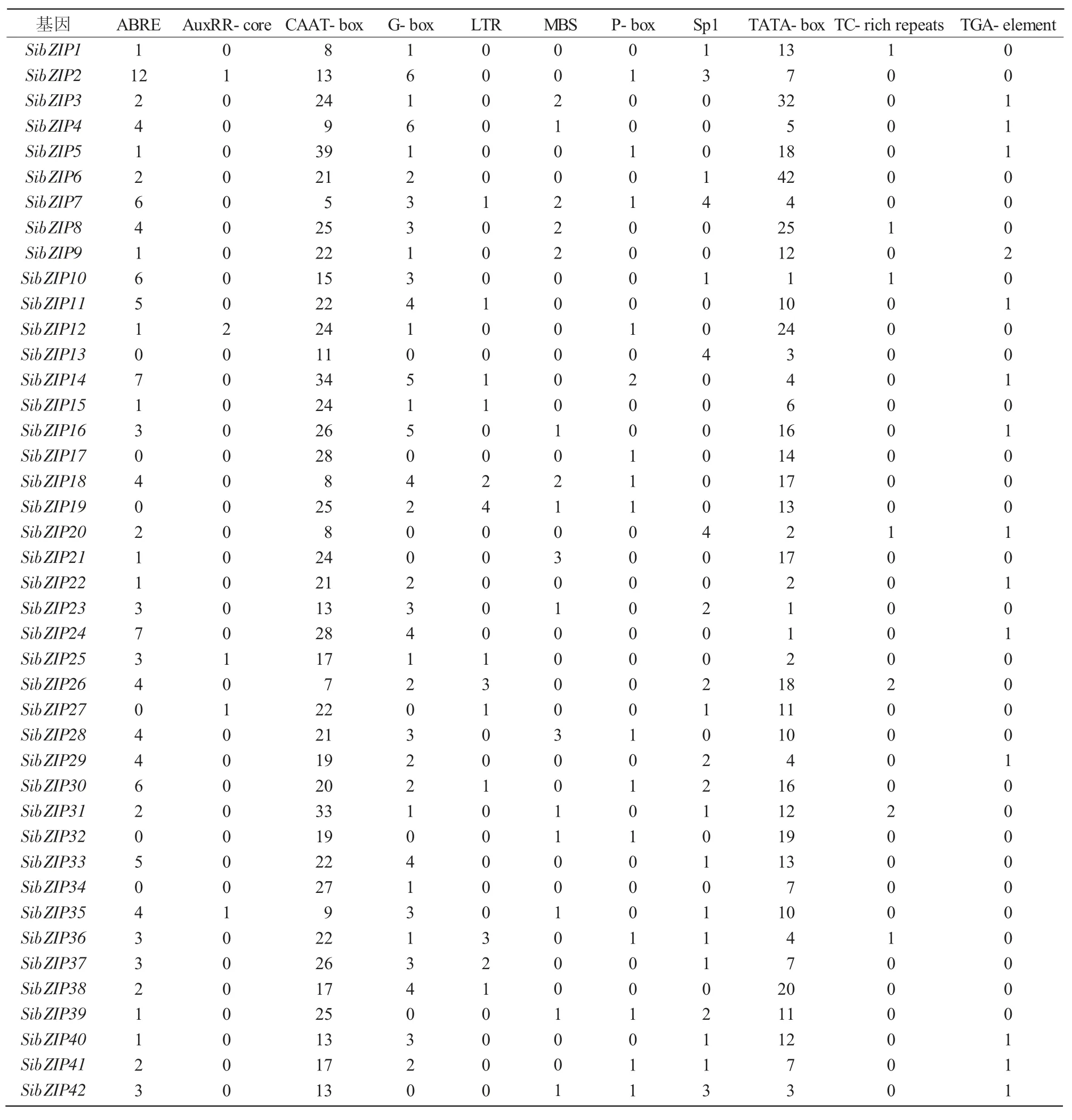
表2谷子bZIP基因启动子顺式作用元件种类与数目
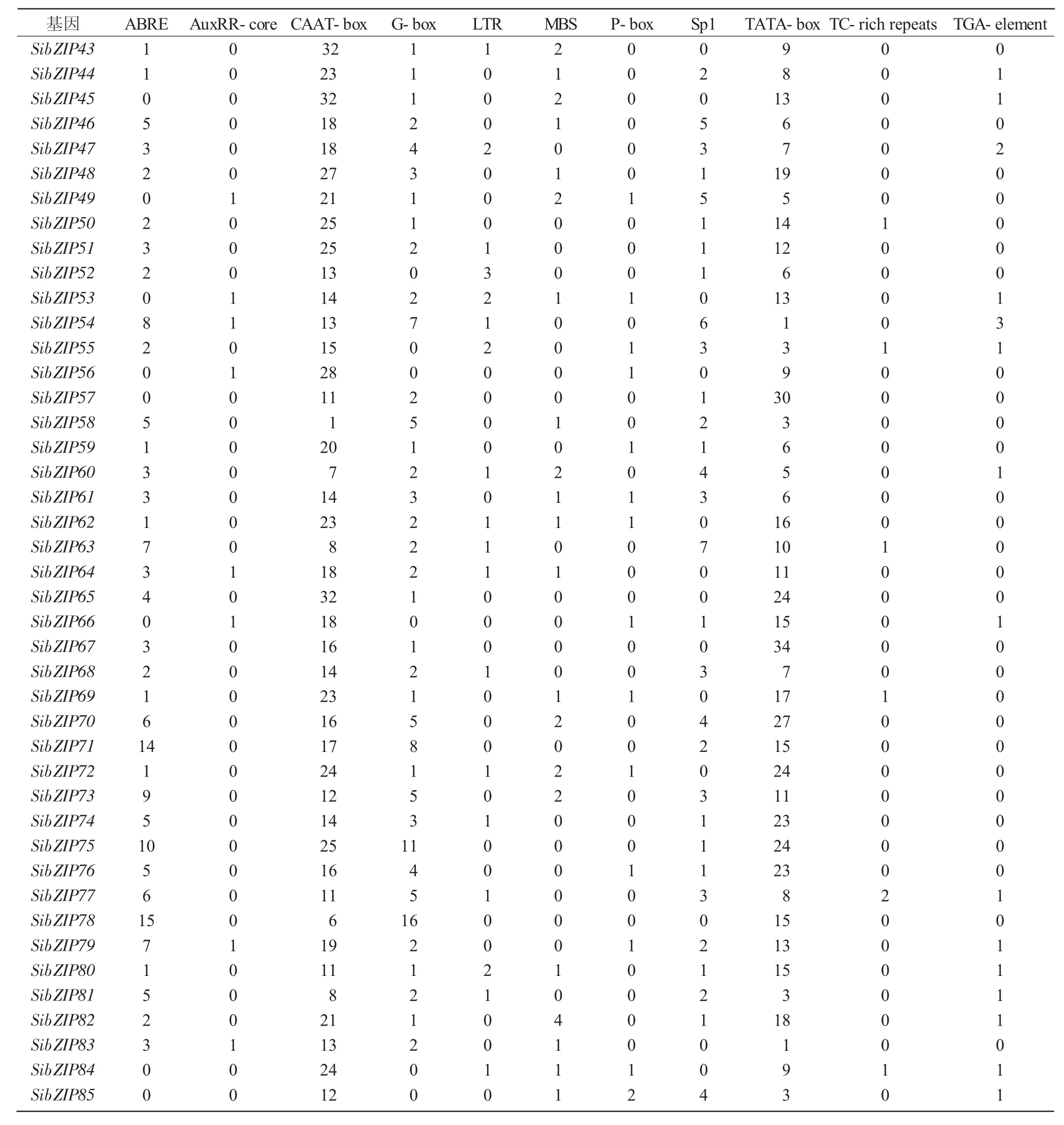
续表2

表3谷子bZIP基因启动子顺式作用元件核心序列与功能
2.4 组织特异性和环境诱导表达分析
利用TBtools软件制作热图,可以较为明显地显示出谷子bZIP转录因子基因在不同组织及不同环境诱导的表达情况(图3),表达情况相似的基因聚在一起。SibZIP3、SibZIP14、SibZIP48、SibZIP66等在不同条件下(10 d光照处理、干旱处理、硝酸处理、尿素处理、氨处理)的根中均有较高的表达量;SibZIP39、SibZIP75和SibZIP76等在圆锥花序中具有较高的表达量;SibZIP19、SibZIP20和SibZIP46等主要在光照下的叶片中表达;SibZIP21、SibZIP67、SibZIP79和SibZIP85均在黄化苗(Etiolated seedling)暗处理培养5 d的情况下具有较高的表达量;SibZIP31在远红光处理下特异表达;SibZIP17和SibZIP71在暗处理的地上部分中表达量较高;SibZIP11、SibZIP12和SibZIP72等在干旱处理的根中表达量较高。
3 结论与讨论
谷子是C4植物,具有耐旱、耐贫瘠、水分利用率高和蒸腾系数低等特性,对生长环境要求较低,耐逆境胁迫能力较其他作物更强。因此,对谷子的基础研究和抗逆基因资源的挖掘成为新的研究热点。bZIP转录因子在植物中不仅能够调节病原防御、胁迫应答等过程,还能够调控光的形态建成、激素信号传导、花发育和种子成熟与萌发等过程。植物的抗逆境胁迫反应机制非常复杂,因此参与调控植物抗逆胁迫反应的转录因子很多,除bZIP外,还有MYB、NAC、WRKY、HSF等[16-21]。
本研究利用生物信息学手段对谷子基因组中bZIP转录因子进行全基因组鉴定,并对所有家族成员进行了系统的生物信息学分析,经鉴定得到的85个SibZIP转录因子,分布在谷子的第1~9条染色体上,部分SibZIP转录因子家族成员具有一定保守性。亲缘关系相近的转录因子往往是由具有相似的基因结构和相似蛋白质模体组成,如SibZIP4/SibZIP44、SibZIP34/SibZIP48。SibZIP转录因子家族保守基序较多、较为复杂,并且SibZIP转录因子具有多种多样的生物学功能。SibZIP转录因子家族成员基因启动子顺式作用元件种类较多,包括与谷子防御胁迫相关的顺式作用元件和谷子生长发育过程相关的顺式作用元件。如SibZIP78、SibZIP75启动子区域内含有较多与光响应有关的顺式作用元件。因此,这2个基因可能参与谷子光反应调控。SibZIP54、SibZIP79基因启动子中含有AuxRR-core、P-box或TGA-element元件的SibZIP转录因子,可能参与植物生长素、赤霉素的调控;SibZIP2、SibZIP71、SibZIP73等转录因子基因启动子富含ABRE、MBS、TC-rich repeats或LTR元件的SibZIP转录因子,可能参与植物非生物胁迫调节、防御和应激反应。
通过对SibZIP转录因子家族成员表达量分析可知,SibZIP3/SibZIP14/SibZIP48、SibZIP39/SibZIP75/SibZIP76和SibZIP19/SibZIP20/SibZIP46分别在根、圆锥花序和叶片中具有较高的表达量。因此,推测这些bZIP转录因子在根、圆锥花序和叶片中特异表达,参与根、圆锥花序和叶片的生长发育。SibZIP21和SibZIP67均在黄化苗暗处理情况下具有较高的表达量,并且SibZIP21和SibZIP67基因结构和蛋白质模体较为相似。因此,推测它们功能相似,可能参与谷子暗反应。SibZIP30和SibZIP61在光照处理的根中具有较高表达量,说明这2个SibZIP基因可能参与谷子根部光反应。SibZIP31在远红光处理下特异表达,说明远红光能诱导SibZIP31基因表达。SibZIP17和SibZIP71在暗处理地上部分中表达量较高,说明黑暗能诱导SibZIP17和SibZIP71表达。SibZIP11、SibZIP12和SibZIP72等在干旱处理的根中表达量较高,说明干旱能够诱导其表达,这3个SibZIP基因可能参与谷子根部的干旱胁迫应答。
在植物中,bZIP转录因子的功能是多样的,不同的家族成员作用不同。因此,对bZIP转录因子的功能和作用机制的研究十分重要。高等植物中部分bZIP转录因子家族成员已经有了较为广泛的研究,接下来可以利用基因克隆、蛋白质组学等技术进一步研究bZIP转录因子在植物中的调控机制,从而更深入了解高等植物遗传进化、生长发育和胁迫应答的过程。
本研究依据生物信息学方法,对谷子bZIP转录因子进行全基因组鉴定,对基因结构、蛋白质理化性质、蛋白质模体、启动子顺式作用元件和表达量水平进行了预测分析,其结果为更深入研究谷子bZIP转录因子的生物学功能及其在谷子中的生长发育调控和逆境胁迫响应机制提供了参考。
