桃果皮毛长度性状的评价及其与几个主要农艺性状的关系
2020-08-17陈旭瑞王力荣朱更瑞方伟超陈昌文王新卫
陈旭瑞,谭 彬,王力荣,朱更瑞,方伟超,陈昌文,王新卫,曹 珂
(1.中国农业科学院 郑州果树研究所,郑州 450000;2.河南农业大学 园艺学院,郑州 450000)
表皮毛是覆盖于植物地上部、特异突起的表皮细胞,根据腺体的存在与否,可分为无腺体和有腺体表皮毛两类;而根据细胞数量的多少,又可分为单细胞和多细胞表皮毛[1]。果实上的果皮毛长短可以影响果实的光合与呼吸作用[2],同时也会在高光强时起到保护作用。也有报道表明果实的表皮毛可以降低病虫害对果实的侵害[3]。
在模式作物拟南芥(Arabidopsisthaliana)上,关于表皮毛的研究比较深入。拟南芥的表皮毛分布于茎、叶片、花瓣、莲座叶和根上,为无腺体、单细胞类型[4]。研究发现,不少转录因子在拟南芥表皮毛的发育过程起着作用[5],如Glabrous1(GL1)[6-7]、GL2[8-9]、Transparent Testa Glabra1(TTG1)[10]、TTG2[9]、Werewolf(WER)[11]、Triptychon(TRY)、Caprice(CPC)[12]以及GL3[13]等。其中3个转录因子TTG1、GL1/WER和GL3对于根毛和表皮毛的发育启动是必须的。上述基因之间通常存在互作,如研究发现TTG1对TTG2具有上位效应,TTG2和GL2在控制表皮毛生长时有重叠作用[9,14],而CPC和TRY却对拟南芥表皮毛发育具有负向调节作用。另一个对表皮毛研究较多的物种为黄瓜,其地上部不同组织均有表皮毛的存在,在黄瓜果实上被称为果刺。果刺性状的突变体gl1果实光滑无刺瘤,且其茎、花萼、子房、卷须和叶片表面都没有表皮毛的覆盖;而另外一个gl2突变体则表现为在果柄、花柄、花萼上有稀疏绒毛,果实亦有稀少果瘤的存在,但叶片、叶柄及茎均光滑无毛[15]。随后,研究者还发现茎叶覆软毛、果实光滑无刺瘤且植株无卷须的黄瓜突变株[16-17]。
桃(PrunuspersicaL.)是一种备受消费者欢迎的时令水果,普通桃果实密布无腺体、单细胞的表皮毛,因其存在会引起部分人群出现过敏症状,影响其食用价值。在很长一段时间内,桃果皮毛一直被作为一个质量性状进行研究[18],即根据其果皮毛的有无将桃分为有毛的普通桃和蟠桃以及无毛的油桃和油蟠桃两大类。Dirlewanger等[19]将桃果皮毛有/无位点定位在第5连锁群上,位于2个限制性片段长度多态性标记AG46和FG之间,与扩增片段长度多态性标记AC-CAA1共分离。俞明亮等[20]研究发现,桃有毛性状与1个简单重复序列标记UDP96-018连锁,遗传距离4.5cM。最近,研究者利用‘Contender’与‘Ambra’的F2杂交群体,将桃果皮毛位点定位到第5连锁群上,区间约1.1cM(635kb物理距离)。根据桃基因组注释结果,研究发现基因PpeMYB25(ppa023143m)是桃果实表皮毛的候选基因,重测序表明该基因的外显子上有一个反转座子插入可能是导致无毛性状形成的原因[21]。
受消费观念的影响,有毛的普通桃在今后一段时间内仍是市场中主要类型。因此,近年来,不少育种者试图选育果皮毛短的新品种,以克服果皮毛对食用不便带来的影响。为此,本试验首先分析了果皮毛长度在6份不同类型桃种质果实发育过程中的变化,测定了238份桃种质果实成熟期的果皮毛长度,探讨果皮毛长度与果实类型和来源之间的关系,对培育果皮毛短的品种以及研究果皮毛发育的生理机制具有重要参考价值。
1 材料与方法
1.1 材 料
第1类材料包括普通桃、蟠桃和油桃种质各2份,果实成熟期基本一致,基本性状见表1。第2类材料为238份有毛的普通桃。所有供试材料均来自中国农业科学院郑州果树研究所国家果树种质桃资源圃,树体约15 a生,砧木为毛桃,栽培措施一致。
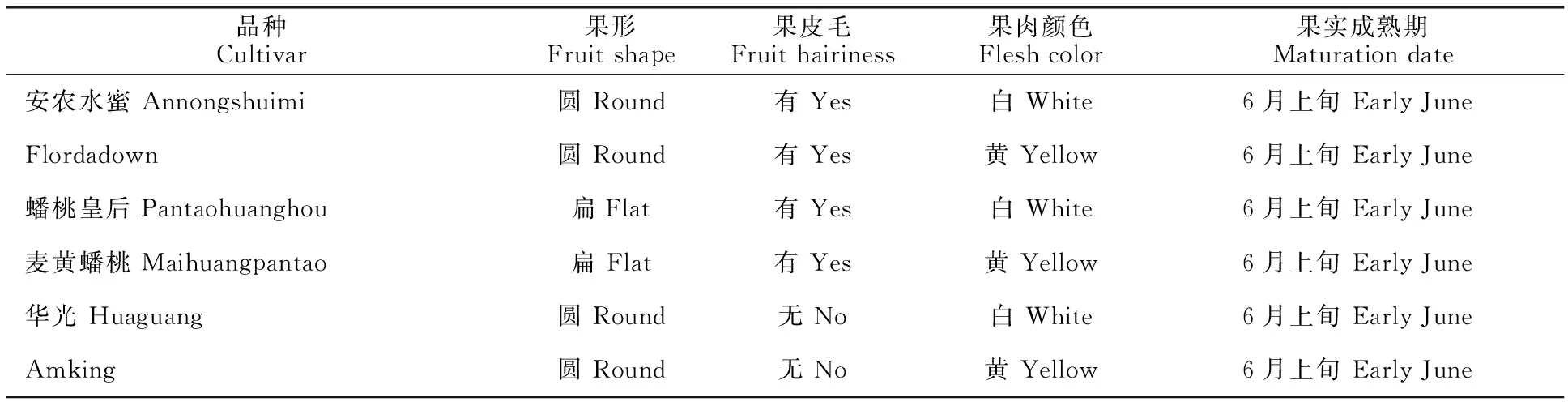
表1 6份供试桃种质及其主要性状Table 1 Key agronomic traits of six peach samples
1.2 果皮毛长度测定
在第1类材料的不同果实成熟期,即从盛花后15 d时至果实成熟,每隔15 d取树冠外围中部果实,再取果实中部缝合线两侧果皮,徒手将其切成3~4 mm厚的薄片,置于Olympus BH-2型(Olympus,日本)显微镜观察果皮毛长度并拍照,放大倍数固定。用Imagetool软件(The University of Texas Health Science Center in San Antonio)分析果皮毛的长短,重复6次。此外,取第2类试材,在不同的果实成熟期按上述方法进行果皮毛的拍照和测量。
1.3 其他主要农艺性状的评价
对于第2类试材,同时引用果形、果肉颜色、果实粘离核、果实风味、果实成熟期、种群分类、地理类群和育成品种来源等信息,均引自《中国桃遗传资源》[22]。
2 结果与分析
2.1 桃不同发育时期果皮毛长短的变化
对6份桃种质果实不同发育期的果皮进行观察,普通桃、蟠桃麦黄蟠桃和油桃各1个品种的显微观察结果如图1所示,其中油桃(图1-C)在整个发育期均没有观察到果皮毛的存在。
有毛的4份种质的果皮毛长度统计结果见图2,结果表明4份种质的果皮毛长度与果实发育的变化趋势一致,即在花后30 d较花后15 d的下降程度明显,随后直至果实成熟呈缓慢下降趋势。
2.2 不同种质果实成熟期的果皮毛长度评价
对供试的238份有毛的桃种质果皮毛长度进行评价,果皮毛长度分布在106.3~343.3 μm间,平均值为193.9 μm,变异幅度为24.77%。果皮毛最短的种质为‘霞晖2号’,最长的为‘白芒蟠桃’。
2.2.1 桃果形与果皮毛长度间的关系 根据果形的不同,桃可以分为扁和圆2种类型。本试验对221份圆桃和17份扁果形桃果皮毛长度进行测定(图3-A),发现圆桃平均果皮毛长度为191.1 μm,显著低于扁果形桃(230.3 μm)。圆桃果皮毛长度分布较为集中,变异幅度为23.99%,低于扁果形桃的26.65%。
2.2.2 桃果肉颜色与果皮毛长度间的关系 根据果实肉色的不同,桃可以分为白肉和黄肉2种类型。研究表明白肉桃平均果皮毛长度为192.2 μm,而黄肉桃平均果皮毛长度为196.9 μm,2种类型间无显著差异(图3-B)。2种类型桃果皮毛长度的变异幅度也类似,分别为24.81%(白肉桃)和24.76%(黄肉桃)。
2.2.3 桃果实粘/离核与果皮毛长度的关系 根据果实粘/离核的不同,桃可以分为粘核和离核。研究表明离核桃平均果皮毛长度为185.9 μm,而粘核桃平均果皮毛长度为199.2 μm,2种类型间差异显著(图3-C)。分析2种类型桃果皮毛长度的分布,离核桃果皮毛长度为107.5~335.3 μm,变异幅度为25.73%;而粘核桃果皮毛长度分布在106.3~343.3 μm,变异幅度为24.33%。

A.安农水蜜;B.麦黄蟠桃;C.华光
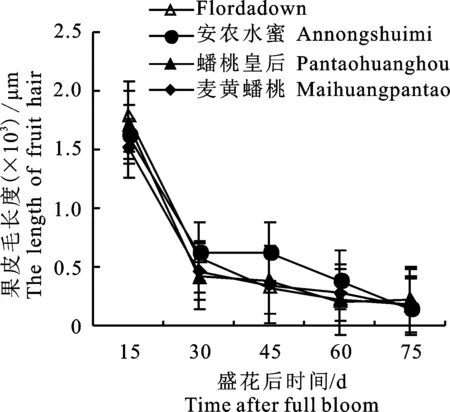
图2 4份种质果皮毛长度Fig.2 Fruit hair lengths during fruit development in four peach germplasms
2.2.4 桃果实风味与果皮毛长度间的关系 在本研究中,通过对果实风味的感官评价,可以将桃分为浓甜、甜、酸甜和酸4种类型。由图3-D可知,不同类型间的平均果皮毛长度相差不大。比较4种类型的变异幅度发现,甜和酸甜2种类型的变异幅度较高,分别为26.23%和26.86%,其次为酸类型果实的种质(22.02%),而甜类型果实的种质果皮毛长度分布最为集中,变异幅度仅为 20.58%。
2.2.5 桃果实成熟期与果皮毛长度间的关系 由图4-A所示,成熟期在7月和8月的中晚熟桃品种的平均果皮毛长度(分别为220.9和226.9 μm)显著高于成熟期在5月和6月的早熟桃平均果皮毛长度(分别为173.3和167.8μm)。但早熟桃和晚熟桃的果皮毛长度变异幅度差异不大,分别在19.16%~20.41%和19.56%~22.09%之间。
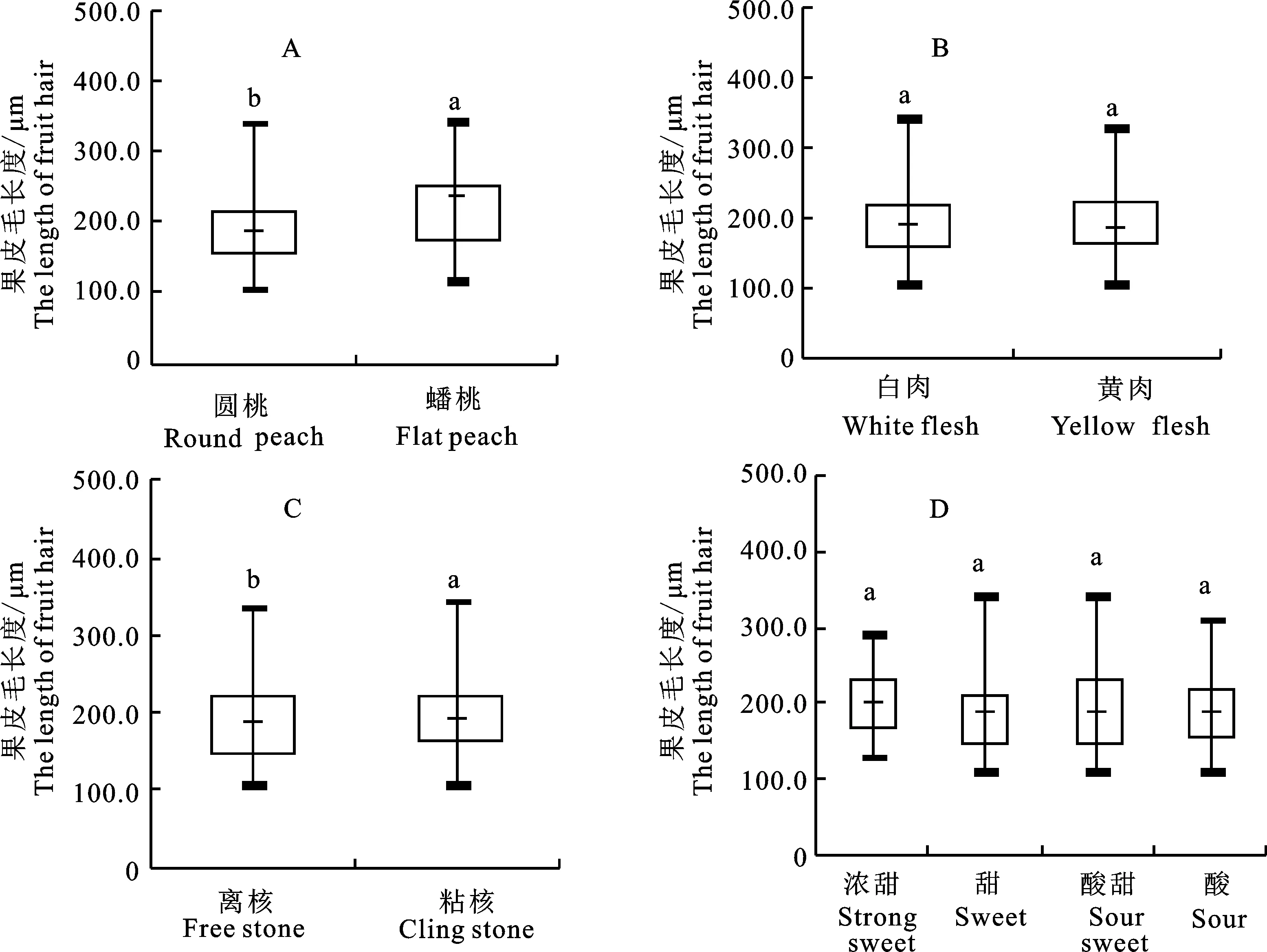
箱形图上不同字母表示组之间差异显著(P<0.05),下同

图4 桃果皮毛长度与成熟期、种群、地理类群以及育成品种类群间的关系Fig.4 Relationship between fruit hair length and fruit maturation date,populations, geographic subpopulations and improvement subpopulations
2.2.6 桃种群与果皮毛长度间的关系 根据桃选育方式的不同,可以将其分为地方品种和育成品种。从图4-B可以看出,地方品种与育成品种的平均果皮毛长度无显著差异,分别为199.2 μm和191.7 μm。育成品种的果皮毛长度分布较为集中,但是有异常高的种质,总体变异幅度为 23.68%,低于地方品种27.72%的变异幅度。
2.2.7 桃不同地理类群与桃果皮毛长度间的关系 在地方品种中,根据其来源的不同可以将供试的桃种质划分为4种地理类群,即华北品种群、长江中下游品种群、西北品种群和云贵高原品种群。由图4-C可知,长江中下游、西北和云贵高原品种群种质的果皮毛长度分别为217.0、209.5和187.6 μm,其中长江中下游品种群果皮毛长度显著高于华北品种群种质。华北、西北和云贵高原品种群的桃果皮毛长度分布集中,变异幅度分别为21.90%、21.45%和15.41%;而长江中下游品种群的种质果皮毛长度分布较为分散,变异幅度高达33.26%。
2.2.8 桃不同育成品种类群与果皮毛长度间的关系 在育成品种中,根据其选育国家的不同,又可以将供试的桃种质划分为中国、日韩和欧美品种群3类。由图4-D可知,3个育成品种类群的平均果皮毛长度无差异,分别为194.9、192.2和192.0 μm。其中,日韩育种品种的果皮毛长度分布较集中,而来自欧美的育成品种果皮毛长度变异幅度较大,达到26.16%。
2.2.9 桃单果质量与果皮毛长度间的关系 本试验同时分析了果皮毛长度与单果质量之间的关系,在该研究中,为了排除果形对单果质量的影响,只选择了圆桃种质。从图5可以看出,单果质量和果皮毛的线性相关系数(r)达到0.321 3 (P<0.01),暗示随着单果质量的增加果皮毛呈上升趋势,但仍然存在不少品种单果质量较高而果皮毛较短,如‘冈山早生’单果质量为249.23 g,但果皮毛长度仅为147.5 μm;‘和谐’单果质量 255.13 g,果皮毛长为181.3 μm。而‘早霞露’却相反,单果质量仅为90.64 g,但果皮毛长度却为241.3 μm。
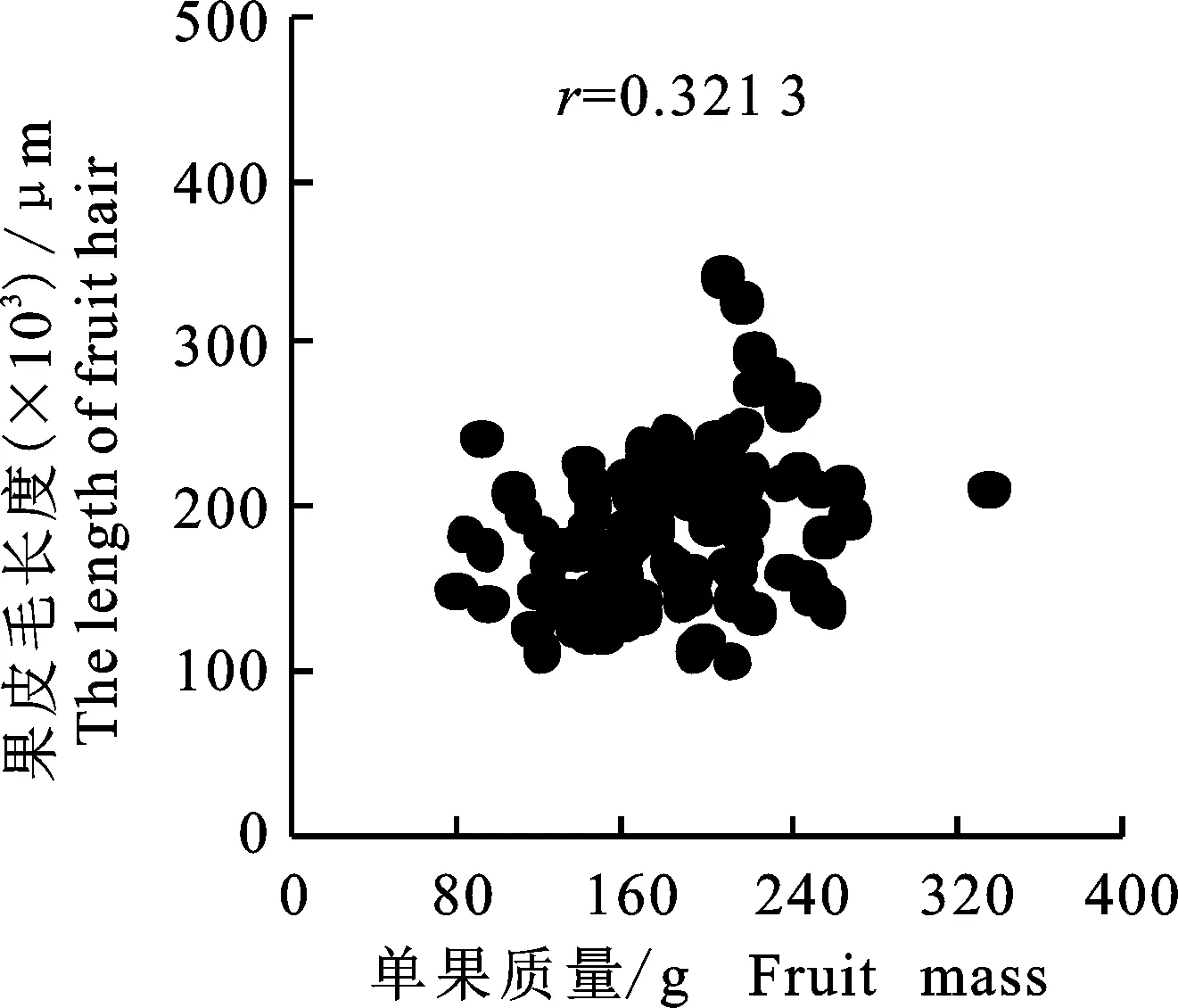
图5 桃果皮毛长度与单果质量性状间的关系Fig.5 Relationship between fruit hairlength and fruit mass traits
3 结论与讨论
果皮毛有/无是桃的一个重要的质量性状,目前对其基因定位和发掘有不少的研究,但果皮毛长度作为一个数量性状,研究其果皮毛长度与其他性状的关系以及是否存在调控基因的报道较少。本试验首先对6份桃种质果实发育期的果皮毛进行了观察,发现果皮毛长度在果实发育期并不是一成不变的,而是在花后时间为30 d时有一个明显的下降,之后下降趋势变缓直至成熟。在猕猴桃‘海沃德’上,花后时间为28 d时雌蕊子房上的果皮毛长度为11.9 μm,直至果实成熟长度增至247.0 μm[23],与桃果实发育过程中果皮毛的变化相反。对于果皮毛长度变化的原因,在拟南芥[24]和番茄[25]上的研究表明,乙烯被认为可以正调控根毛的发育。而在桃上,乙烯含量随着果实成熟而上升,该结果表明在桃上果皮毛长度的变化与乙烯的关系与拟南芥根毛发育的调控机制有明显差异。由于在成熟果实上(图1),显微观察并未发现存在表皮毛断裂现象。同时在猕猴桃上,笔者认为成熟果实上较短的果皮毛可能是较长果皮毛的早期发育形态[26],因此推测,桃果皮毛随发育时期不断下降可能与果皮毛的更新有关,即成熟的果皮毛不断脱落,而新的果皮毛虽然不断产生,但受激素调控其发育受到抑制。
同时,本试验对238份桃成熟期果实表皮毛长度进行测定发现,果形为圆的桃果皮毛长度小于果形扁的桃,离核桃果皮毛小于粘核,早熟桃果皮毛长度小于晚熟桃,华北品种群小于长江中下游品种群。首先,应该注意到这种关系是否存在必然联系,抑或是种质份数较少导致的错误推定呢?例如在本试验中,果形为扁的桃种质仅有17份,远小于果形为圆的桃种质;而在其他性状的不同类群间没有明显的偏分离现象,例如早熟桃种质有116份,中晚熟桃有112份,因此,可以排除品种数量对结果的影响。同时,虽然在前人研究中发现陕北高原丘陵沟壑区6种优势物种的表皮毛长度随植物立地水分的减少而减少,表皮毛密度则相反[26];随海拔增加,紫椴果皮毛变短,表皮毛密度增大[27]。但在本试验中,所有的材料均来自同一果园,采用相同的栽培措施,因此可以排除外部环境对果皮毛长度的影响。
为进一步探讨不同性状之间的关系,本试验将上述4种性状分别选择其中1个为导致果皮毛长度变化的主要因子,进而判定其他3个性状与果皮毛的关系是否受到该主要因子的影响。如首先将果形扁作为导致果皮毛较长的主要因子,而在离核桃和粘核桃中,果形扁的桃分别占1.19%和10.96%,即粘核种质与长果皮毛间的关系可能是受到果形与果皮毛之间关系的影响;在早熟与晚熟桃中,果形扁的桃分别占6.03%和 9.01%,即果实成熟期性状对果皮毛长度的影响仍然受到果形的影响;在华北和长江中下游品种群中,果形扁的桃分别占到0%和31.82%,同样表明地理类群与果皮毛长度的关系可能是受到果形对果皮毛调控的影响。按照这种方法,将粘核作为导致果皮毛较长的主要因子,发现其与果形、果实成熟期和地理类群影响果皮毛长度之间并无内在联系;将晚熟性状作为导致果皮毛较长的主要因子,发现其与果形、粘/离核和地理类群调控果皮毛长度之间也无内在联系;将地理类群作为导致果皮毛长度变化的主要因子,发现其与果形调控果皮毛长度之间有关,而与粘/离核、果实成熟期性状调控果皮毛长度之间并无内在联系。综上所述,相比其他性状,果形可能与果皮毛发育具有一定的内在联系。
