丹波黑大豆GmDREPP基因的克隆与耐铝性
2020-06-24魏运民余世田韩蓉蓉颉永红刘卢生蒋曹德玉永雄
王 燚,魏运民,余世田,韩蓉蓉,颉永红,刘卢生,蒋曹德,玉永雄,*
(1.西南大学 动物科技学院,重庆 400715; 2.重庆市高校草食动物工程中心,重庆 400715)
酸性土壤(pH<5.0)约占全世界潜在可耕种土地的40%~50%[1],植物在酸性土壤中生长受到铝毒、酸害、低磷等因子胁迫,而铝毒害是酸性土壤中限制作物生长和产量的关键因子[2]。研究表明,铝在酸性土壤中主要以离子形式存在,并富集在植物根系的分生区和伸长区[3]。游离的Al3+能通过诱导脂质过氧化反应引起活性氧损伤[4]、破坏细胞骨架[5]等过程阻止细胞分裂,从而抑制根的生长。目前生产上主要采用大量施加石灰来缓解土壤酸性的方法,成本高且容易造成土壤板结,相比之下,理解植物适应铝胁迫的分子机制,利用抗铝基因培育耐铝性强的农作物新品种是解决酸性土壤铝毒问题的长效策略[6]。
多年研究表明,植物主要有2种耐铝机制[7-8]:(1)铝耐受机制,即铝进入植物后被解毒和隔离于液泡,如铝敏感蛋白3(aluminum sensitive protein 3, ALS3)是植物耐铝所必需的蛋白[9],能将进入细胞的铝离子转移至液泡,减轻铝毒害;(2)铝排斥机制,目前最典型的铝排斥机制是铝诱导根系分泌有机酸(organic acid,OA),螯合根际Al3+形成无毒化合物,防止铝离子进入根尖细胞。柠檬酸是铝离子的强效螯合剂,其分泌受多药与毒性复合物的排出转运蛋白(multidrug and toxic compound extrus-ion protein,MATE)影响[10]。
发育调节质膜多肽(DREPP)是一类植物特异性蛋白,参与调控植物生长发育与逆境应答过程,在植物抗逆过程中发挥重要作用[11-12]。Kochian等[13]发现,拟南芥(Arabidopsisthaliana)中DREPP家族蛋白AtPCaP2(plasma membrane-associated cation binding protein 2)在干旱胁迫下对种子的萌发有正向调节作用,且过表达该基因能通过参与生长激素调节增强拟南芥对干旱胁迫的适应性。低温胁迫下AtPCaP2也可通过激活cbf介导的转录调控网络,在寒冷胁迫应答中发挥重要作用[14]。水稻(OryzasativaL.)中DREPP同源基因OsDREPP2的表达与盐胁迫相关,盐胁迫下OsDREPP2在耐盐品种中的转录水平显著高于盐敏感品种[15]。DREPP蛋白家族成员PtBP1(peaT1 binding protein 1)可以诱导植物的防御反应[16]。但有关DREPP基因在植物耐铝方面的作用还未见报道。
丹波黑大豆是原产于日本的耐铝大豆品种[17]。本研究室在前期试验中测定的丹波黑大豆在酸性土壤条件下根尖芯片组数据和铝胁迫条件下转录组数据中发现,GmDREPP基因的表达量均明显上调,由此推测该基因与植物耐铝相关,但其耐铝功能尚不明确。为此,本研究以日本丹波黑大豆为试验材料,克隆GmDREPP基因,构建pCXSN-GmDREPP过表达载体,遗传转化烟草,并研究鉴定该基因的耐铝性,以期为培育耐铝新品种提供基因资源。
1 材料与方法
1.1 材料
1.1.1 植物材料
本研究所用的耐铝品种丹波黑大豆由日本引进,转基因用烟草为实验室保存的无菌苗。
1.1.2 主要试剂
大肠埃希菌(Escherichiacoli)感受态DH5α(北京天恩泽生物技术有限公司);农杆菌(Agrobacteriumtumefaciens)感受态GV3101(唯地生物技术有限公司);RNA提取试剂RNAiso Plus、反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa,大连);半定量PCR酶Taq、高保真酶Prime STAR、荧光定量PCR酶SYBR Premix ExTaqⅡ、T4DNA Ligase、限制性内切酶XcmⅠ(TaKaRa,大连);胶回收试剂盒Gel Extraction Kit(Omega,美国);质粒提取试剂盒Plasmid Mini Kit I(Omega,美国)。
1.2 方法
1.2.1 植物材料培养
25 ℃避光萌发丹波黑大豆(Glycinemaxcv. Tamba)种子,待其发芽2 d后转至霍格兰营养液进行培养。转基因试验选用本实验室保存的烟草无菌苗叶片为外植体。耐铝比较试验选取长势一致的野生型和转基因烟草植株进行扦插水培,生根后培养与丹波黑大豆一致。
1.2.2 目的基因克隆与生物信息学分析
根据基因组序列设计引物(表1),PCR扩增采用高保真酶Primer STAR,回收片段连接T载体后转化大肠埃希菌DH5α,挑取阳性菌落摇菌送出测序并确认。
将获得的GmDREPP基因的CDS序列进行生物信息学分析(表2),并进行BLAST比对,挑选同源性较高的序列比对后用MEGA7.0构建进化树。
1.2.3 铝胁迫下丹波黑大豆GmDREPP的表达分析
用0.5 mmol·L-1CaCl2对培养2周的丹波黑大豆预处理24 h,然后选取长势一致的植株进行试验,重复3次。在50 μmol·L-1AlCl3(pH 4.5、0.5 mmol·L-1CaCl2)条件下进行不同组织、不同根段和不同处理时间3个试验:(1)不同组织试验,处理24 h后取植株根、茎、叶、子叶测定;(2)根的不同区段试验,处理24 h后取距根尖0~2、2~4、4~6 cm的3个区段测定;(3)不同处理时间试验,处理0、3、6、9、12、15、18、21、24 h分别取根测定。另外,进行不同浓度试验:分别用0、25、50、75、100 μmol·L-1AlCl3(pH 4.5、0.5 mmol·L-1CaCl2)处理24 h后取根测定。采取的根、根段或组织用液氮速冻备用。然后分别提取RNA,用于qRT-PCR。
表1 引物序列及其用途
Table 1 Primer sequence and purpose

引物Primer 引物序列(5′—3′)Primer sequence (5′—3′)用途PurposeGmDREPP-FGmDREPP-RATGGGTTATTGGAAGTCTAAGGTCAAGGCTTTGGTGGTTCTG基因克隆、半定量RT-PCR、载体构建Gene cloning, semi-quantitative RT-PCR, vector constructionqRTGmDREPP-FCAGTCAAGGAATTGTGACGTTT荧光定量qRTGmDREPP-RCAAACAGGACTTCCATAATGGCFluorescent quantitation18S rRNA-FATGATAACTCGACGGATCGC大豆内参18S rRNA-RCTTGGATGTGGTAGCCGTTTSoybean reference geneβ-actin-FTCCGGCGACGGTGTCTCACA烟草内参β-actin-RCGCGGACAATTTCCCGTTCAGCTobacco reference geneNtMATE-FCAGCATTGTGTTCTTGCTCATTC荧光定量NtMATE-RGCCTATCCTCCCGAAACCAFluorescent quantitationNtALS3-FTCGCGATGACATCAAGATACAA荧光定量NtALS3-RCAGGAGATAGAGCAATAACCAACGAFluorescent quantitation
表2 生物信息学分析工具与数据库
Table 2 Bioinformatics analysis tools and databases

项目Item名称Name网站Website同源序列Homologous sequenceBLASThttps://blast.ncbi.nlm.nih.gov/Blast.cgi理化性Physical and chemical propertiesProtParamhttps://web.expasy.org/protparam/二级结构Secondary structureSOPMAhttps://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html三级结构Tertiary structurePDBhttps://www.rcsb.org/跨膜区Transmembrane areaTMHMMhttp://www.cbs.dtu.dk/services/TMHMM/信号肽Signal peptideSingalPhttp://www.cbs.dtu.dk/services/SignalP/功能结构域Functional domainCDDhttps://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi
1.2.4 构建过表达载体
用XcmⅠ对pCXSN载体进行酶切并回收。以丹波黑大豆cDNA为模板,DREPP-F、DREPP-R为引物,高保真酶Primer STAR进行扩增,回收片段进行加A实验(72 ℃,1 h)后再回收。回收产物与酶切回收的pCXSN载体用Solution I在16 ℃连接12 h,连接产物转化大肠埃希菌DH5α。根据菌落PCR检测结果,挑取阳性菌落培养提取质粒pCXSN-GmDREPP,酶切检测后送出测序验证,确认获得植物表达载体pCXSN-GmDREPP。
1.2.5 遗传转化烟草并筛选转基因植株
将构建好的植物表达载体pCXSN-GmDREPP通过冻融法转入农杆菌GV3101,待形成菌落后进行菌落PCR检测,挑取阳性菌落培养后侵染烟草。选取在筛选培养基上长势良好的烟草植株,先后提取其DNA进行PCR扩增,RNA作为模板反转录的cDNA进行PCR扩增,筛选转基因烟草植株。
1.3 转基因烟草的耐铝性鉴定
1.3.1 烟草根相对伸长率的测定
将野生型(wild type,WT)和转基因烟草1代种子用75%乙醇和30%次氯酸钠溶液消毒灭菌后置于1/2MS培养基上至萌发,挑取萌发后长势一致的幼苗转移至含50 μmol·L-1AlCl3的1/2MS培养基上生长,2周后观测根长并记录。相对伸长率(%)=50 μmol·L-1AlCl3(pH 4.5、0.5 mmol·L-1CaCl2)条件下根的伸长量/无铝胁迫条件下根的伸长量×100。
1.3.2 过氧化物酶(POD)、超氧化物歧化酶(SOD)活性测定
烟草水培2周,选取根生长一致的WT和转基因烟草,50 μmol·L-1AlCl3(pH 4.5,0.5 mmol·L-1CaCl2)条件下处理24 h,取烟草根尖,用过氧化物酶(POD)、超氧化物歧化酶(SOD)活性试剂盒(上海优选生物科技有限公司)分别测定转基因烟草和野生型烟草的过氧化物酶、超氧化物歧化酶活性。
1.3.3 柠檬酸分泌量测定
预培养烟草2周,取根生长一致的WT和转基因烟草在50 μmol·L-1AlCl3(pH 4.5,0.5 mmol·L-1CaCl2)条件下处理24 h,测烟草根质量。用柠檬酸(CA)ELISA试剂盒(江西江蓝纯生物试剂有限公司)测定处理液中的柠檬酸含量,即为转基因烟草和野生型烟草的柠檬酸分泌量。
1.3.4 丙二醛(MDA)含量测定
烟草预先水培2周,选取根生长一致的WT和转基因烟草,在50 μmol·L-1AlCl3(pH 4.5,0.5 mmol·L-1CaCl2)条件下处理24 h,取烟草根尖,用丙二醛(MDA)含量试剂盒(上海优选生物科技有限公司),分别测定转基因烟草和野生型烟草根中的MDA含量。
1.3.5 过表达烟草中耐铝相关基因的表达分析
为了解过表达GmDREPP对烟草耐铝重要基因NtALS3和NtMATE的影响,根据烟草基因组序列设计引物(表1),以长势一致的转基因烟草和野生型烟草的根为样品,提取RNA反转录得到cDNA,进行qRT-PCR。
1.3.6 伊文思蓝和苏木精对烟草根尖染色
选取长势一致的水培WT和转基因烟草苗,在50 μmol·L-1AlCl3(pH 4.5,0.5 mmol·L-1CaCl2)溶液中处理24 h,分别在伊文思蓝和苏木精染液中染色30 min,待洗去根尖表面的染液后置于体视镜下观察拍照。
2 结果与分析
2.1 GmDREPP基因的克隆与序列分析
扩增所得目的片段与预测大小基本一致,目的片段经纯化回收,连接pMD19-T 载体后测序,结果如图1所示。该序列全长624 bp,编码207个氨基酸,是一个完整的开放阅读框。根据BLAST比对结果推测该基因是丹波黑大豆DREPP基因,命名为GmDREPP。
Prot Param在线工具预测分析,GmDREPP蛋白分子式为C1028H1646N254O346S1,相对分子质量23 131.93,理论等电点4.95;氨基酸组成中谷氨酸占比最高(21.3%),赖氨酸和丙氨酸占比17.4%、10.1%,精氨酸和色氨酸占比最低(0.5%)。

atg,起始密码子;-,终止密码子。atg, Initiation codon; -, Termination codon.图1 GmDREPP的核苷酸序列及其编码的氨基酸序列Fig.1 Nucleotide and predicted amino acid sequences of GmDREPP
该蛋白的正、负电荷的残基均为0;不稳定系数50.26(>40),表明GmDREPP为不稳定蛋白。
2.2 生物信息学分析结果
SingalP软件预测GmDREPP蛋白存在信号肽的可能性极低,为0.04%。经TMHMM软件预测GmDREPP蛋白无跨膜区,该蛋白1~207位氨基酸均位于细胞表面。CDD在线分析认为GmDREPP蛋白第1~185位氨基酸形成其唯一一个功能结构域,且与DREPP家族蛋白功能结构域基本吻合。SOPMA软件预测GmDREPP蛋白的二级结构,其中α-螺旋占比58.94%,延伸链占比4.83%,β转角占比2.90%,无规则卷曲占比33.33%(图2)。利用PDB软件预测GmDREPP蛋白三级结构,结果如图3所示。
2.3 GmDREPP蛋白的进化树分析
用BlastP比对GmDREPP的氨基酸序列,用软件MEGA 7.0取同源性大于70%的氨基酸序列构建进化树(图4)。如图所示,丹波黑大豆的蛋白序列与大豆(Glycinemax)、菜豆(Phaseolusvulgaris)和豇豆(Vignaunguiculata)的DREPP蛋白序列在进化分枝中亲缘关系较近,且与另外15种豆科植物聚为一枝,表明GmDREPP在同一科中同源性很高。

图中蓝色线条表示α-螺旋,红色线条表示延伸链,绿色线条表示β-转角,紫色线条表示无规则卷曲。Blue lines represented alpha helixs, red lines represented extend strands, green lines represent beta turns, and the purple lines represented random coils in the figure.图2 GmDREPP蛋白的二级结构预测Fig.2 Secondary structure prediction of GmDREPP
图3 GmDREPP 蛋白质三维结构预测图Fig.3 Predicted 3D structure of GmDREPP
2.4 GmDREPP基因表达分析
实时荧光定量结果表明,不同浓度AlCl3处理24 h后,随铝浓度的升高,丹波黑大豆根系中GmDREPP的表达量随铝浓度的升高而上升,且Al3+浓度高于50 μmol·L-1后其表达量无显著变化(图5-A)。50 μmol·L-1AlCl3(pH 4.5,0.5 mmol·L-1CaCl2)条件下处理24 h后,GmDREPP在丹波黑大豆的根、茎、叶、子叶中的表达量存在显著差异,表达量由高到低分别为茎、根、子叶和叶(图5-B),根的3个区段中又以根尖0~2 cm表达量最高(图5-C),且根系GmDREPP的表达量在处理12 h后趋于稳定(图5-D)。

红色方框处是丹波黑大豆的GmDREPP蛋白。GmDREPP of Tamba Black soybean was showed in the red box.图4 GmDREPP蛋白的进化树分析Fig.4 Phylogenetic tree of GmDREPP

A,不同浓度Al3+处理24 h大豆根中GmDREPP的表达量;B,GmDREPP在大豆根、茎、叶和子叶中的相对表达水平;C,GmDREPP在根不同区段的相对表达水平;D,50 μmol·L-1 Al3+处理下不同时间大豆根尖GmDREPP表达量。不同柱上无相同小写字母表示差异显著(P<0.05),无相同大写字母表示差异极显著(P<0.01)。下同。A, Expression of GmDREPP in soybean roots under different concentrations of Al3+ for 24 h; B, Relative expression level of GmDREPP in roots, stems, leaves and cotyledon of soybean; C, Relative expression level of GmDREPP in roots different segments; D, Time course of GmDREPP expression in soybean roots in response to 50 μmol·L-1 Al3+ treatment. Different lowercase and uppercase letters above the columns represent significant differences among treatments at P<0.05 and P<0.01, respectively. The same as below.图5 GmDREPP的相对表达量Fig.5 Expression analysis of GmDREPP gene
2.5 表达载体的构建与转基因烟草的筛选
将构建好的质粒pCXSN-GmDREPP(图6)经单酶切电泳检测后送华大基因测序,验证结果表明已获得全长CDS序列。将构建好的载体转入农杆菌GV3101,待形成菌落后进行菌落PCR检测,挑取阳性菌落培养后遗传转化烟草,培养获得烟草植株42株。根据提取DNA进行PCR检测,获得7株阳性植株(图7-A)。提取阳性植株RNA反转录所得cDNA进行RT-PCR检测,获得5株有表达活性的植株(图7-B),并从中选取3个表达水平较高的植株,即图7中9、6、4号转基因株系GmDREPP-2、GmDREPP-4、GmDREPP-5,测定相对表达量(图8)并进行耐铝性分析。
2.6 耐铝性检测
2.6.1 转基因对铝胁迫下烟草根系的影响
无铝胁迫条件下,转基因烟草GmDREPP-2、GmDREPP-4、GmDREPP-5与WT烟草根系生长状况良好,根的伸长量无明显差异(图9-A)。铝胁迫下,烟草根系生长均受抑制,但3个转基因株系的根明显较WT长(图9-B),根的相对伸长量显著(P<0.05)高于野生型(图9-C),GmDREPP过表达的烟草植株较野生型植株高大。这一结果表明,GmDREPP在烟草中的过表达减轻了铝胁迫对烟草根系生长的抑制作用。
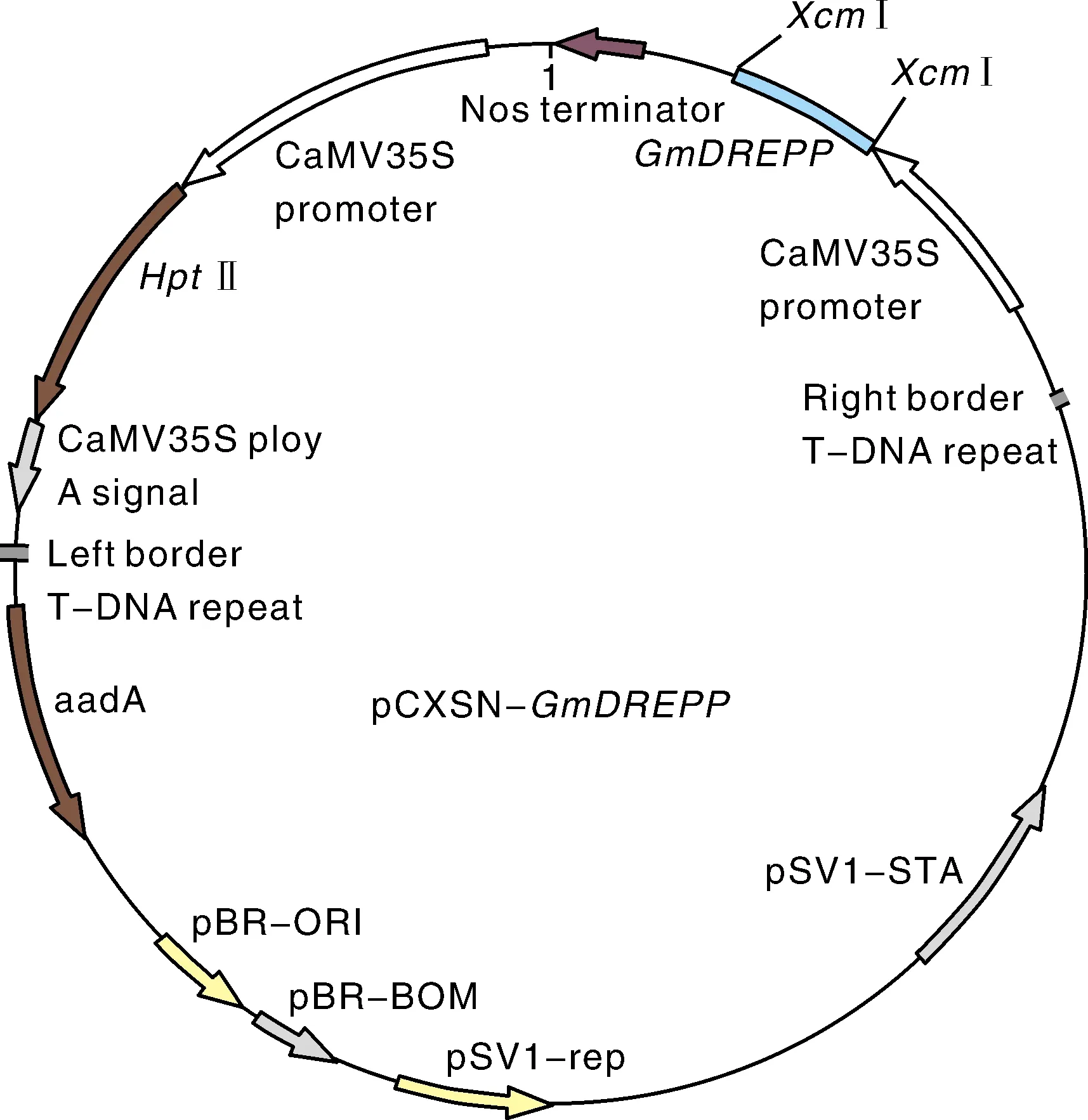
图6 植物过表达载体pCXSN-GmDREPP结构示意图Fig.6 Diagram structure of plant overexpression vector pCXSN-GmDREPP

A,GmDREPP基因DNA水平的检测;B,GmDREPP基因mRNA水平的检测。1,转pCXSN载体烟草;2,WT;3~9,转基因烟草株;9,GmDREPP-2;4,GmDREPP-4;6,GmDREPP-5。A, Detection of DNA level of GmDREPP; B, Detection of mRNA level of GmDREPP. 1, Transgenic tobacco with pCXSN vector; 2, WT; 3-9, Transgenic tobacco lines; 9, GmDREPP-2; 4, GmDREPP-4; 6, GmDREPP-5.图7 转基因烟草阳性植株筛选Fig.7 Screening of transgenic tobacco plants

图8 转基因烟草GmDREPP的相对表达量Fig.8 Relative expression of GmDREPP in transgenic tobacco
2.6.2 铝胁迫对转基因植株根系抗氧化的影响
如图10所示,与WT相比,铝处理24 h后转基因株系的POD、SOD活性显著(P<0.05)上升,产生MDA的量更少。这种变化说明,铝胁迫下烟草过表达GmDREPP能提高根系POD、SOD活性,降低根尖MDA含量。
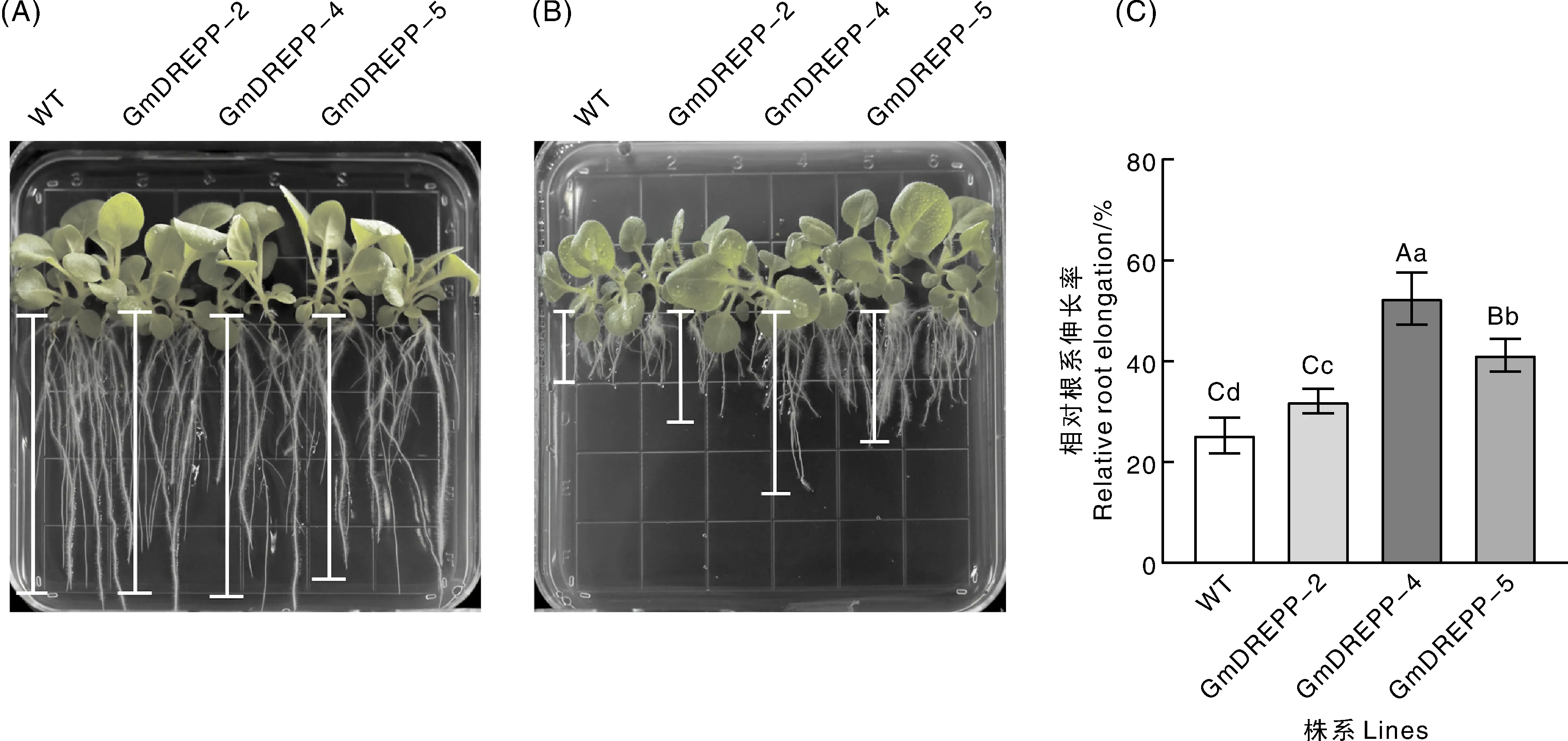
A,无铝条件下烟草根系伸长量;B,Al胁迫(50 μmol·L-1)下烟草根系伸长量;C,烟草相对根系伸长率。A, Root elongation of tobacco without Al; B, Root elongation of tobacco under aluminum stress (50 μmol·L-1); C, Relative root elongation of tobacco.图9 过表达GmDREPP对铝胁迫下烟草根系的影响Fig.9 Effects of overexpression GmDREPP on root system of tobacco under aluminum stress

数据以鲜质量计。下同。Data was detected based on fresh weight. The same as below.图10 铝胁迫对转基因植株根系抗氧化酶活性的影响Fig.10 Effects of aluminum stress on antioxidant enzyme activity of transgenic tobacco lines
2.6.3 铝胁迫对转基因植株根系柠檬酸分泌的影响
铝胁迫处理3个转基因烟草GmDREPP-2、GmDREPP-4、GmDREPP-5,以及野生型烟草12 h后,分别测定其根系柠檬酸分泌量。如图11所示,转基因烟草的根系柠檬酸分泌量极显著(P<0.01)高于野生型,为野生型的2.9倍以上,说明过表达GmDREPP能显著促进烟草分泌柠檬酸响应铝胁迫以缓解铝毒害。
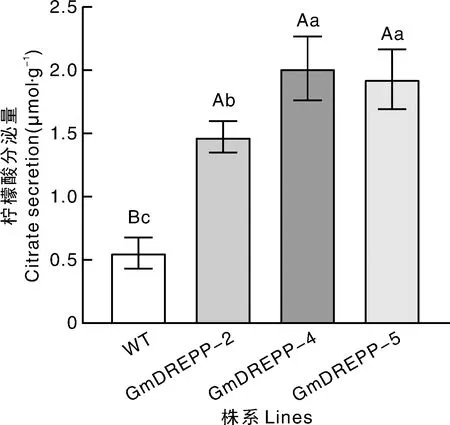
图11 铝胁迫下烟草根系柠檬酸分泌量Fig.11 Citrate secretion of tobacco roots under aluminum stress

图12 野生型和转基因烟草中NtALS3和NtMATE的相对表达量Fig.12 Relative expression levels of NtALS3 and NtMATE in WT and transgenic tobacco lines
2.6.4 过表达烟草中NtALS3和NtMATE的表达分析
NtALS3和NtMATE已经被证实是铝胁迫的响应基因。图12显示,无铝胁迫(CK)时,转基因株系GmDREPP-2中NtALS3基因的表达量显著高于WT和转基因株系GmDREPP-4、GmDREPP-5,WT和转基因株系GmDREPP-4、GmDREPP-5的NtALS3基因表达量无显著差异;同时,3个转基因株系的NtMATE基因表达量均显著高于WT。铝处理(pH 4.5,0.5 mmol·L-1CaCl2,50 μmol·L-1AlCl3)12 h后,野生型和转基因烟草中NtALS3和NtMATE的相对表达量均极显著(P<0.01)升高,且3个转基因株系的NtALS3和NtMATE基因表达量均极显著(P<0.01)高于WT。
2.6.5 铝胁迫对转基因烟草根尖的伤害
取铝处理(pH 4.5,0.5 mmol·L-1CaCl2,50 μmol·L-1AlCl3)12 h后的转基因烟草和野生型烟草根尖染色,结果如图13所示。由图可知,转基因烟草根尖着色明显较浅,说明转基因烟草根尖受损较轻,铝离子积累少。

图13 野生型和转基因烟草根尖染色Fig.13 Staining at the root tips of WT and transgenic lines
3 讨论
本研究通过分子克隆技术从丹波黑大豆中克隆获得编码丹波黑大豆GmDREPP蛋白基因的CDS序列(全长624 bp,编码207个氨基酸),构建植物表达载体pCXSN-GmDREPP,并成功遗传转化烟草。根尖是识别铝毒害、积累铝离子的主要部位[18]。李季肤等[19]研究发现,地毯草铝响应基因AcABCG1在根中表达量较高且主要在根尖中表达,本研究的qRT-PCR结果与此一致,即铝胁迫条件下,根系中根尖的表达量最高,推测GmDREPP可能是铝胁迫的响应基因。
根系伸长是细胞分裂的过程,铝毒害对植物根系生长的抑制作用主要通过阻止细胞分裂实现[20]。本研究中,铝处理对烟草根系生长有明显抑制作用,但转GmDREPP烟草根的相对伸长率明显高于野生型,说明GmDREPP能缓解烟草根系生长受到的抑制。伊文思蓝和苏木精染色结果也与根部生长情况一致,进一步说明GmDREPP增强了烟草的耐铝能力。
SOD和POD是植物体内重要的抗氧化酶,在缓解逆境胁迫引起的氧化损伤中发挥重要作用[21-22]。经50 μmol·L-1AlCl3处理12 h后,转基因植株的SOD、POD活性明显高于野生型,与耐铝型杉木(Cunninghamialanceolata)幼苗叶片[23]和玉米(ZeamaysL.)[24]的SOD、POD活性变化趋势一致。由此可知,过表达GmDREPP基因能提高烟草中抗氧化酶的活性,这也是转GmDREPP烟草提高耐铝性的原因之一。
植物可以通过细胞膜上的特异受体和信号转导系统,接受并转导环境中的铝信号,激活下游相关基因(如MATE,ALS3等)表达抵御铝毒害[25-26]。MATE和ALS3是缓解铝胁迫的关键基因。ALS3蛋白能将进入植物细胞中的铝离子转运至液泡隔离[27],同时转运尿苷二磷酸葡糖(uridine diphosphate glucose,UDPG)至细胞壁合成纤维素固定铝离子,提高植物耐铝性[28-29]。植物抵御铝毒害最有效的机制之一是铝诱导植物根系分泌有机酸[30],其中柠檬酸能与铝络合形成稳定常数较大的复合物,被认为是强效的铝解毒剂。铝胁迫条件下,MATE蛋白通过促进植物柠檬酸分泌[31],使柠檬酸与铝络合形成稳定无毒复合物缓解铝毒害[32]。本研究中铝胁迫条件下,转GmDREPP烟草中NtALS3和NtMATE基因的表达量显著升高,推测过表达GmDREPP可能通过影响其体内次生代谢物或激素的变化间接影响NtALS3和NtMATE的表达[33-35],从而增强转基因烟草的耐铝性。但他们之间的具体关系目前还没有研究,这也是后期探索的目标。同时,铝处理条件下转基因烟草NtMATE的相对表达量极显著(P<0.01)高于野生型,且转GmDREPP后烟草根系柠檬酸的分泌量提升了1.5倍以上,说明过表达GmDREPP能通过上调MATE基因表达从而促进烟草分泌柠檬酸响应铝胁迫。
综上所述,转GmDREPP烟草显著上调了缓解铝胁迫的关键基因NtALS3和NtMATE的表达,增强了抵御铝毒害的能力,其中NtMATE的表达量升高促进了转基因烟草的柠檬酸分泌;同时提高抗氧化酶SOD、POD活性来降低铝胁迫对质膜的氧化损伤。过表达GmDREPP从以上2个方面增强了转基因烟草的耐铝性,证明该基因具有耐铝功能。
