鸽源新城疫病毒辽宁分离株LNPPMV17全基因测序与遗传演化分析
2020-06-08郑百利刘承惠徐静怡李冰
郑百利,刘承惠,徐静怡,李冰
(锦州医科大学畜牧兽医学院,辽宁 锦州 121001)
鸽新城疫是一种以腹泻和神经紊乱为主要特征的病毒性传染病[1]。其病原是一种抗原变异的新城疫病毒(Newcastle disease virus,NDV),又称禽副黏病毒Ⅰ型(paramyxovirus 1,APMV-1)[2]。1978年在中东地区首次分离到了鸽源禽副黏病毒Ⅰ型,随后传播到北非、欧洲等地,不久该病毒迅速传播至世界各地[3]。1985年我国首次在香港分离到鸽副黏病毒,随后广东、福建、四川等多省份相继出现鸽副黏病毒的相关报道[4-5]。鸽新城疫的流行给对我国肉鸽和赛鸽的饲养都造成了严重的经济损失,该病成为威胁养鸽业的重要疫病之一[6]。
NDV属于副黏病毒科(Paramyxoviridae)、禽腮腺炎病毒亚科(Avulavirinae)、正禽腮腺炎病毒属(Orthoavulavirus)[7],基因组为单股负链不分节段的RNA病毒,包含6个结构基因,按照3′-NP-P-M-F-HN-L-5′的顺序编码结构蛋白,另外还有2个非结构蛋白(V和W)[8]。根据全基因组序列长度不同,将NDV分为ClassⅠ和ClassⅡ两类,其中ClassⅠ类毒株基因组全长15 198 nt,ClassⅡ类毒株为15 186 nt或15 192 nt。NDV只有1个血清型,但根据F蛋白全基因序列和最新分类命名体系, ClassⅠ类NDV仅有1个基因型(含3个亚型);ClassⅡ类毒株至少可分为20个基因型和48个亚型[9]。禽副黏病毒分为9个血清型,即APMV-1~9,引起鸽发病的毒株多数属于APMV-1型。另外,APMV-7型也可在鸽群中流行,并引起火鸡和鸵鸟发病[10]。虽然引起鸽发病的副黏病毒F蛋白上具有强毒株特点的胰蛋白酶裂解位点,但是鸽副黏病毒对鸡的致病力弱,甚至没有致病力[11]。2017年4—6月,辽宁地区某赛鸽公棚幼龄鸽陆续发生疑似ND病例,发病率约20%,病鸽主要表现为精神沉郁、食欲下降、排绿色水样稀便、头颈歪斜;剖检病鸽可见脑部水肿、消化道黏膜轻微出血。本研究对辽宁地区分离的鸽副黏病毒进行全基因测序,并将其与国内外典型的毒株进行序列比对,分析遗传演化特点,为辽宁地区鸽源禽副黏病毒Ⅰ型的分子流行病学研究提供有价值的参考资料。
1 材料与方法
1.1 材料
鸽副黏病毒辽宁分离株由锦州医科大学畜产品质量与安全重点实验室分离保存;大肠杆菌DH5α感受态、快速琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒购自北京天根生物科技有限公司;pMD18-T载体购自宝生物工程有限公司;血液总RNA快速提取试剂盒(离心柱型)购于北京百泰克生物技术有限公司;2×TaqMaster Mix(含染料)购自康为世纪生物科技有限公司。
1.2 方法
1.2.1 病毒RNA的提取及cDNA的合成
按RNA快速提取试剂盒使用说明,从细胞培养物中提取总RNA,并按常规反转录方式进行反转录,反转录反应条件:65 ℃ 5 min,42 ℃ 1 h,70 ℃ 5 min。将反转录获得的cDNA置于-20 ℃保存备用。
1.2.2 引物的设计与合成
根据GenBank中鸽副黏病毒毒株Pi/SH/CH/0168/2013(登录号:KT163263)设计16对引物,对分离病毒的全基因序列进行扩增,引物信息详见表1,由上海生物工程股份有限公司合成。
表1 全基因引物信息
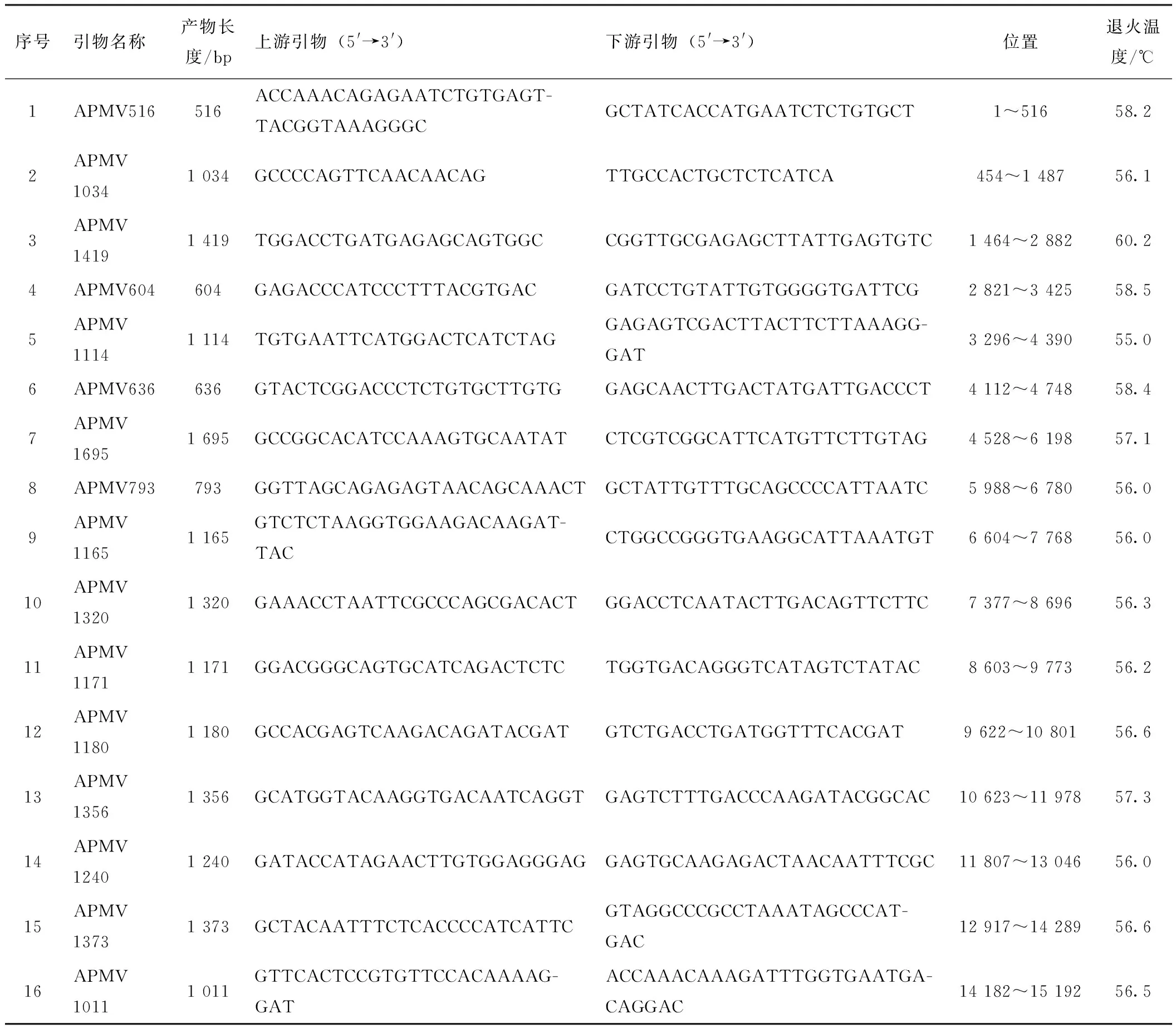
序号引物名称产物长度/bp上游引物(5'→3')下游引物(5'→3')位置退火温度/℃1APMV516516ACCAAACAGAGAATCTGTGAGT-TACGGTAAAGGGCGCTATCACCATGAATCTCTGTGCT1~51658.22APMV10341 034GCCCCAGTTCAACAACAGTTGCCACTGCTCTCATCA454~1 48756.13APMV14191 419TGGACCTGATGAGAGCAGTGGCCGGTTGCGAGAGCTTATTGAGTGTC1 464~2 88260.24APMV604604GAGACCCATCCCTTTACGTGACGATCCTGTATTGTGGGGTGATTCG2 821~3 42558.55APMV11141 114TGTGAATTCATGGACTCATCTAGGAGAGTCGACTTACTTCTTAAAGG-GAT3 296~4 39055.06APMV636636GTACTCGGACCCTCTGTGCTTGTGGAGCAACTTGACTATGATTGACCCT4 112~4 74858.47APMV16951 695GCCGGCACATCCAAAGTGCAATATCTCGTCGGCATTCATGTTCTTGTAG4 528~6 19857.18APMV793793GGTTAGCAGAGAGTAACAGCAAACTGCTATTGTTTGCAGCCCCATTAATC5 988~6 78056.09APMV11651 165GTCTCTAAGGTGGAAGACAAGAT-TACCTGGCCGGGTGAAGGCATTAAATGT6 604~7 76856.010APMV13201 320GAAACCTAATTCGCCCAGCGACACTGGACCTCAATACTTGACAGTTCTTC7 377~8 69656.311APMV11711 171GGACGGGCAGTGCATCAGACTCTCTGGTGACAGGGTCATAGTCTATAC8 603~9 77356.212APMV11801 180GCCACGAGTCAAGACAGATACGATGTCTGACCTGATGGTTTCACGAT9 622~10 80156.613APMV13561 356GCATGGTACAAGGTGACAATCAGGTGAGTCTTTGACCCAAGATACGGCAC10 623~11 97857.314APMV12401 240GATACCATAGAACTTGTGGAGGGAGGAGTGCAAGAGACTAACAATTTCGC11 807~13 04656.015APMV13731 373GCTACAATTTCTCACCCCATCATTCGTAGGCCCGCCTAAATAGCCCAT-GAC12 917~14 28956.616APMV10111 011GTTCACTCCGTGTTCCACAAAAG-GATACCAAACAAAGATTTGGTGAATGA-CAGGAC14 182~15 19256.5
1.2.3 全基因序列的扩增及克隆测序
采用RT-PCR方法对全基因序列进行分段扩增。按照常规方法进行反转录,PCR反应体系(30 μL):模板cDNA 3 μL;引物2 μL(上下引物各1 μL);dH2O 10 μL;2×TaqMaster Mix 15 μL,按照表1中退火温度对16条引物进行扩增,扩增出目的条带后纯化PCR产物,连接到pMD18-T载体上并转入DH5α感受态细胞中,涂于含氨苄青霉素(100 μg/mL)的LB培养皿上进行筛选,培养16 h后挑取单个菌落,经PCR验证为阳性,用质粒小提试剂盒提取质粒,放于-20 ℃保存,每个样品重复3次,将所提质粒送至上海生物科技有限公司进行测序。
1.2.4 全基因序列比对分析
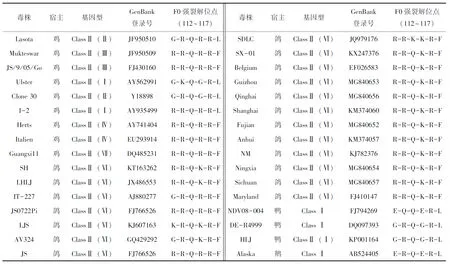
测序结果经NCBI-BLAST比对正确后,根据各片段之间相互重叠的区域,用DNAman拼接获得完整的基因序列。采用MEGA 7.0和DNAStar 7.0软件将分离毒株的F蛋白、M蛋白和全基因序列与GenBank中公布的代表性毒株的基因序列及氨基酸序列进行同源性比对,并制作进化树;参照文献[9]提出的新城疫病毒分类命名体系对分离毒株进行分型。用于比对分析的序列信息见表2。
表2 全基因序列比对分析的代表毒株序列信息
2 结果与分析
2.1 辽宁分离株全基因各片段PCR扩增
利用RT-PCR方法扩增出了鸽副黏病毒辽宁分离株全基因各片段,产物用1.5%的琼脂糖凝胶电泳鉴定,目的片段与预期片段大小基本一致,电泳结果见图1。
2.2 测序结果及全基因序列的拼接
根据测序结果重叠区域用DNAman进行拼接、校正,经BLAST验证得到鸽源禽副黏病毒Ⅰ型全基因序列,命名为LNPPMV17。分离毒株全基因序列共15 192个核苷酸,有6个结构蛋白,各结构蛋白的具体位置详见表3。将鸽源禽副黏病毒Ⅰ型辽宁分离毒株LNPPMV17的全基因序列提交GenBank,经检验后获得登陆号MK335466。
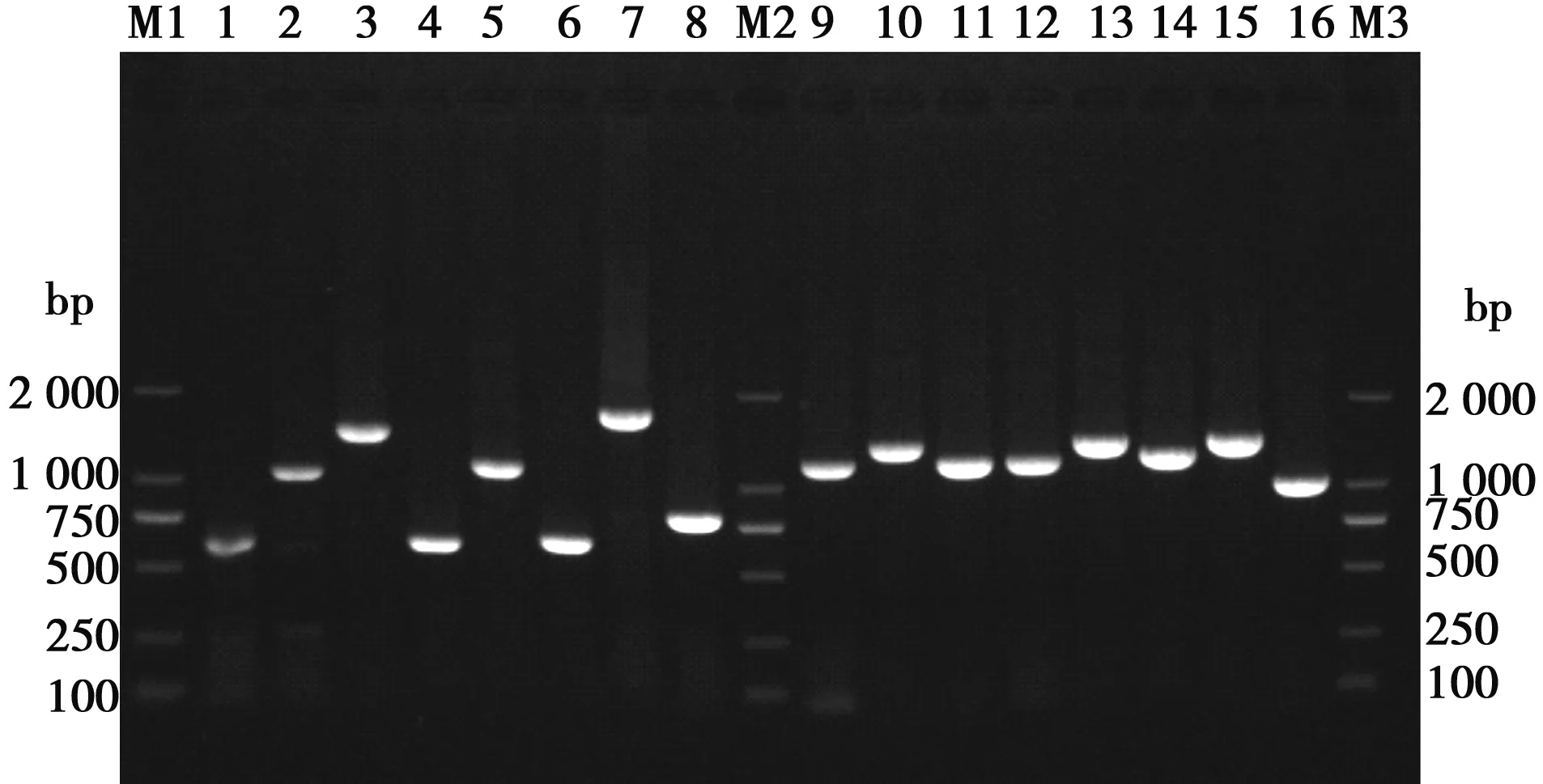
M1、M2、M3. DL2000 DNA marker;1~16. PCR产物
表3 辽宁分离株LNPPMV17结构蛋白基因信息
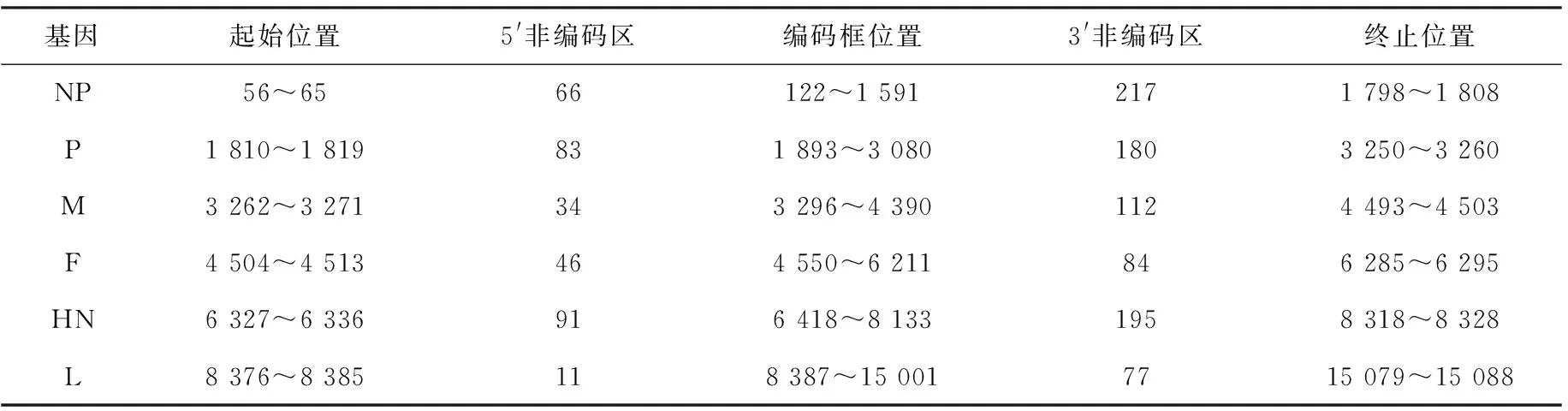
基因起始位置5'非编码区编码框位置3'非编码区终止位置NP56~6566122~1 5912171 798~1 808P1 810~1 819831 893~3 0801803 250~3 260M3 262~3 271343 296~4 3901124 493~4 503F4 504~4 513464 550~6 211846 285~6 295HN6 327~6 336916 418~8 1331958 318~8 328L8 376~8 385118 387~15 0017715 079~15 088
2.3 基于F蛋白基因序列分析
LNPPMV17辽宁分离株F基因全长1 792 bp,开放阅读框的长度为1 662 bp,编码553个氨基酸。分离毒株F0裂解位点的氨基酸序列为112R-R-Q-K-R-F117,第116位氨基酸为精氨酸(R),符合禽副黏病毒特点,第112位点、113位点、115位点均为碱性氨基酸,并且第117位氨基酸为亮氨酸(F),符合强毒株特征,详见图2。

图2 辽宁分离株F0裂解位点的氨基酸序列
用DNAStar 7.0软件对辽宁分离株LNPPMV17与32个国内外参考毒株的F蛋白全基因序列进行比较,分离毒株与19株鸽源副黏病毒核苷酸同源性在91.8%~98.7%之间,与9株鸡源副黏病毒的同源性在82.7%~87.2%之间,与4株其他禽类副黏病毒的同源性68.7%~84.6%之间。用MEGA 7.0软件对F蛋白全基因序列进行分析并构建了系统进化树,结果显示,鸽源副黏病毒辽宁分离株与Qinghai株(MG840656)同源关系最近,核苷酸同源性为98.7%,核苷酸距离仅为1.4%(小于5%)。根据传统的基因分型方法,LNPPMV17分离株属于ClassⅡ类新城疫病毒,基因型为Ⅵ型,基因亚型为 k亚型;根据最新分类命名系统,LNPPMV17分离株属于Ⅵ.2.1.1.2.2,详见图3。
2.4 基于M蛋白氨基酸分析
辽宁分离株M蛋白基因全长1 240 nt,其开放阅读框长度1 095 nt,编码364氨基酸。运用DNAStar 7.0软件对LNPPMV17分离株的氨基酸序列与32个参考毒株进行同源性分析,辽宁分离株与参考株氨基酸同源性为88.5%~98.6%。氨基酸同源性比较分析发现:鸽源副黏病毒M蛋白第36位点氨基酸残基为精氨酸(R),其他禽类均为谷氨酰胺(Q);另外,鸽源副黏病毒第68位点氨基酸残基为异亮氨酸(I),其他禽类均为缬氨酸(V),详见图4。推测M蛋白第36位、68位氨基酸可能是禽副黏病毒Ⅰ型在鸽宿主体内进化、适应的关键氨基酸。
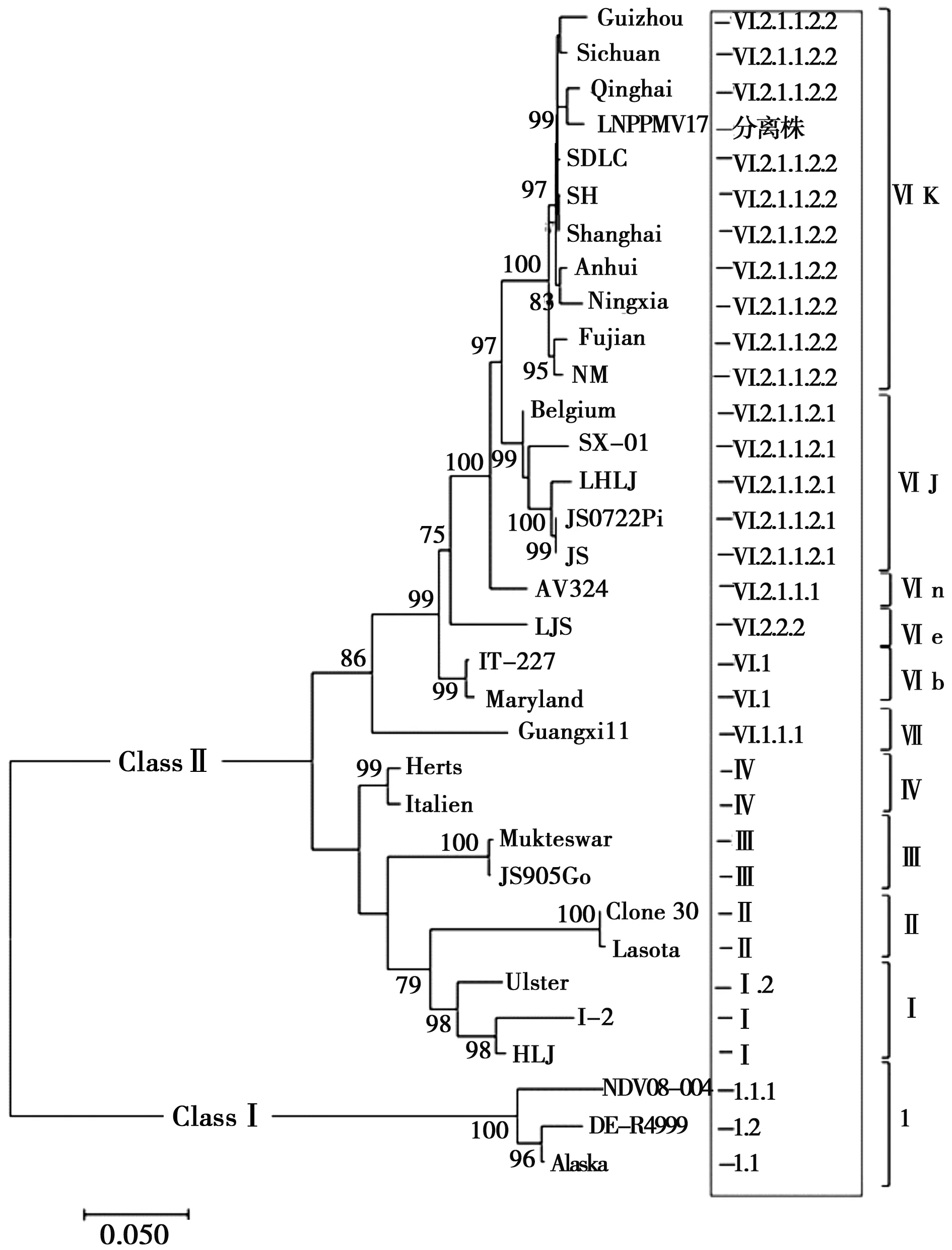
框内是根据文献[10]提出的最新分类命名标准查询指南对现有新城疫毒株的基因分型结果,中括号右边是2012年以来的分类方式
图3 F蛋白全基因序列系统进化树
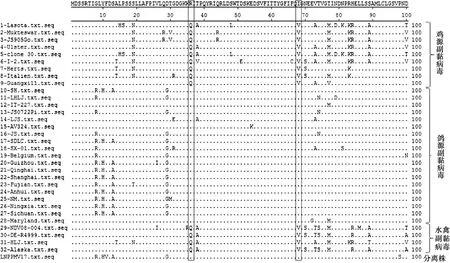
图4 M蛋白氨基酸比对结果
2.5 辽宁分离株全基因序列分析
2.5.1 辽宁分离株全基因序列同源性分析
运用DNAStar 7.0软件对辽宁分离株LNPPMV17的全基因核苷酸序列与GenBank下载的32个参考毒株基因序列进行同源性分析。辽宁分离株与参考株的全基因核苷酸同源性为71.7%~98.7%,其中辽宁分离株与19株鸽源副黏病毒同源性为91.8%~98.7%,同源性在95%以上的有11株,与Qinghai株同源性最高(98.7%);与9株鸡源副黏病毒同源性为82.8%~87.3%;与其他禽类副黏病毒同源性为71.7%~84.6%。根据同源性比对分析结果,分离株与水禽的同源性较低,与鸭NDV08-004株同源性仅为71.7%。
2.5.2 基于全基因序列的系统进化分析
根据辽宁分离株LNPPMV17与32个参考毒株的全基因序列,应用MEGA 7.0软件(Maximum Likelihood)构建系统进化树。由图5可以看出,ClassⅠ群毒株与ClassⅡ群毒株的遗传关系较远,分别形成单独的分支,此结果与F基因构建的系统进化树一致;鸽源副黏病毒均在同一条分枝上,且亲缘关系较近,该结果与F基因构建的系统进化树结果基本一致,基因Ⅵ型中的亚型均在同一分支上,但核苷酸同源性略有差异;辽宁分离株LNPPMV17与Qinghai毒株的亲缘关系最近,其次为国内分离的Shanghai、SDLC毒株,而与疫苗株Lasota、Clone30的亲缘关系较远。
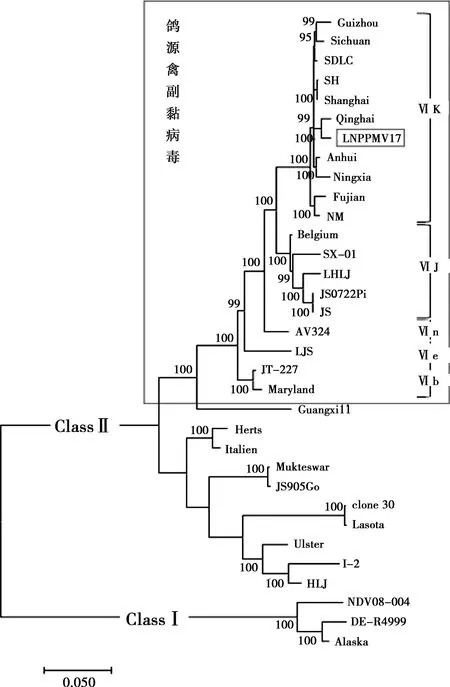
图5 基于各毒株全基因序列的系统进化树
3 讨论
禽副黏病毒有9个血清型,能引起鸽发病的主要是禽副黏病毒Ⅰ型(APMV-1),APMV-7型在鸽群中也有流行,并可引起火鸡和鸵鸟暴发感染。1996—2005年Liu等[12]从中国分离到14株鸽源禽副黏病毒Ⅰ型,其中10株属于基因Ⅵb型,3株属于基因Ⅶ型,1株是疫苗毒属于基因Ⅱ型。郭虹波[13]对2010—2012年从中国吉林、广东、辽宁、黑龙江分离到的8株鸽源禽副黏病毒Ⅰ型分析表明,8株均属于基因Ⅵb型。2015年王林等[14]从北京分离到1株鸽副黏病毒(BJP13),遗传进化分析结果表明BJP13属于基因Ⅵb型。本试验从辽宁地区分离1株鸽源禽副黏病毒Ⅰ型,基因组全长15 192 nt,符合ClassⅡ病毒特征;根据F蛋白全基因序列遗传进化分析表明,分离毒株属于基因Ⅵ型,并且F0裂解位点的氨基酸序列为112R-R-Q-K-R-F117,符合目前我国鸽源禽副黏病毒Ⅰ型特点;根据全基因序列分析表明,辽宁分离株LNPPMV17与Qinghai株亲缘关系最近,同源性为98.7%。由此可见,辽宁分离株LNPPMV17属于新城疫ClassⅡ类病毒,基因型为Ⅵ型。
M蛋白是一种核质穿梭蛋白,在NDV复制周期中起定位作用[15]。目前研究发现鸽副黏病毒M蛋白第36位氨基酸为R,其他禽类副黏病毒均为Q;同时,构建了R→Q的变异毒株,发现变异毒株在鸽体内复制能力和致病力明显下降[16],推测禽副黏病毒Ⅰ型在鸽体内适应过程中,出现的独特氨基酸突变,也可能与宿主特异性有关。本试验分离的LNPPMV17毒株M蛋白第36位氨基酸也是R,与32个参考毒株进行氨基酸同源性分析发现第68位氨基酸为Ⅰ,其他禽类均为V。由此可见,鸽源禽副黏病毒Ⅰ型M蛋白第36位、68位氨基酸的差异可以作为区别其他禽类副黏病毒Ⅰ型的标志。
近年来,辽宁省养鸽业发展迅速,特别是肉鸽、赛鸽养殖量较大。由于养殖量的增加与鸽的流通频繁,鸽新城疫的危害越来越严重。虽然鸽源禽副黏病毒Ⅰ型与鸡新城疫病毒同属一个血清型,但分别属于2个不同的适应毒株,因此单纯用鸡新城疫疫苗免疫不能有效保护鸽免受鸽源禽Ⅰ型副黏病毒的感染[17]。辽宁分离株与疫苗Lasota株、Clone 30株全基因组同源性仅为83%,这可能是影响鸡新城疫疫苗对鸽保护效率不高的重要因素之一。另外,通过鸽源禽副黏病毒Ⅰ型全基因序列比对分析发现,与辽宁分离株同源性在98%以上的有6个病毒株,分别源于青海、上海、山东、安徽、云南和四川。由此可见,我国流行的鸽源禽副黏病毒Ⅰ型同源性较高,可能是近年来赛鸽也迅速发展,鸽流通增加而导致的。
