南宁老虎岭林区多种外来植物入侵程度与影响因子模型解释
2020-06-08黄小荣
黄小荣
(广西壮族自治区林业科学研究院,南宁 530002)
外来物种(alien species)是指人类有意或无意地在其过去或现在自然分布之外引入的物种[1~2]。随着人们对入侵物种、入侵影响和入侵控制的日益重视[3],探索外来物种入侵机理成为当前生态学研究热点[4~5];通常用与入侵有关的环境因子来解释入侵机理,包括道路条件、林冠郁蔽度、林下植被高度和乡土物种丰富度等因素。道路是外来入侵物种传播的主要途径之一[6],道路修建干扰生态环境使路边更容易被外来植物拓殖[7],植物种子附着在车轮上可沿路传播数米至数千米[8~9]。对于森林群落,林冠郁蔽度和林下植被高度可能影响外来入侵,光照条件较差限制喜光入侵植物的种子萌发和生长繁殖,林下植被高度可能阻碍风传型植物种子落地生根。土著物种丰富度与外来入侵的关系存在一定的争议,在不同生境可能出现正相关、负相关或不相关[10]。自然群落通常存在多个外来种和多种环境因子,但大多数生物入侵研究只注重单一物种入侵[11],或只考虑单个因子和主效应;现实生态系统过程的复杂性,导致至关重要的生态效应往往不是主效应,而是与其他效应的互作[11]。为了有效地管理外来物种,在众多外来物种持续存在的情况下,我们必须了解多个外来物种与多个影响因子之间的互作效应。
广西具有沿边沿海地理位置和温暖湿润的气候,是外来植物入侵重灾区。老虎岭位于广西南宁市北郊,近些年各种基因收集种质资源库的建立、林区水泥路的铺设、附近苗圃育苗、养蜂等活动,为外来植物的传播、生长和繁衍提供了机会,外来植物的种群数量和分布范围持续增加,改变了原有的植被风貌。老虎岭林区分布有各种林龄、树种和抚育时间的林分,为比较外来植物入侵与林区各种环境因子的关联性提供了天然的条件。本研究在老虎岭进行了分区样方调查,拟比较不同林型、林龄、抚育时间、路边与林内的外来植物入侵程度,探讨多种因素对入侵的影响,为多因素对多个外来植物入侵的互作效应分析提供一个具体而详细的范例。
1 材料和方法
1.1 研究区概况
研究区在老虎岭林区一、二林班,调查样地南起广西林科院树木园,北至老虎岭水库西北部的珍贵树种任豆资源库,南北、东西相距均为1 600 m,中心坐标108°20.4′E,22°55.8′N,海拔135~217 m。研究区地处南亚热带季风气候区,年均降雨量1 304 mm。大部分土壤为砂页岩发育而成的砖红壤性红壤,低山丘陵地貌。广西林科院自1956年建院以来,在老虎岭陆续营建了松杉人工林、引种树木园以及加勒比松(Pinuscaribaea)、湿地松(P.elliottii)等国外松资源林;至1974年,老虎岭的栽培乔灌木达684种。1995年在高速路桥边原松树皆伐迹地建立了马尾松(Pinusmassoniana)优质种源林,2016年底进行了疏伐、砍灌和补植大叶栎。2009~2015年,在老虎岭水库西南部、西部和西北部的松杉林皆伐迹地上分别营建了斑皮桉(Eucalyptusmaculata)、邓恩桉(E.dunnii)和珍贵树种基因库;2016年将该区的土道铺筑成水泥路;林分的抚育工作外包给同一包工队,在每年1~2月砍灌割草。老虎岭林区树木基本连片,相对僻静和冷清,除造林和抚育施工、苗圃育苗和养蜂之外其他干扰甚少。
1.2 样地设置和测量
老虎岭林区的外来物种分6个区(A~F)进行样方调查,区间间隔200 m以上。每个区沿样线调查20个路边小样方和20个林内小样方,A、C和F区在道路一侧、样线长200 m,B、D和E区在道路两侧、样线长100 m;林内样线在距林缘滴水线10 m处设置;沿样线间隔10 m取一个小样方,尺寸均为1 m×1 m;每个区的第一个小样方随机起点,随后样方等距间隔,不因人为偏好而移位[12]。6个区共240个小样方,分为15个组:路边A、路边B、路边C、路边D、路边E、路边F、A区马尾松、B区红锥(Castanopsishystrix)、B区尾巨桉(Eucalyptusurophylla×E.grandis)、C区邓恩桉、D区西桦(Betulaalnoides)、D区任豆(Zeniainsignis)、E区麻栎(Quercusacutissima)、E区国外松和F区斑皮桉。样方调查在2018年6~7月进行,记录每个小样方内植物名和盖度,以小样方中心为圆心的直径5 m样圆记录高度>3 m的林冠郁蔽度,分3个类型记录小样方位置高于林道、齐平林道或低于林道。
1.3 外来植物入侵的定量
为了避免主观地划分外来种和入侵种,本文的外来物种定义包括林下所有外来植物[6]。根据万方浩[13]、林建勇[14~15]、唐赛春[16]等编目的名录确定哪些是外来植物,包括林下高度≤5 m的非禾本科草本、禾本科草本、灌木以及藤本植物;不包括人工挖坎种植的引进树种;以小样方中所有外来植物占小样方植被总盖度的百分比作为外来植物占比(alien ratio),代表外来植物入侵程度(invasion)。
1.4 其他因子的获取
咨询相关人员和查阅广西林科院内部资料,确定各林分的林龄。将抚育时间分为4个等级:之前4~6个月抚育(M1)、之前6~20个月抚育(M2)、之前20~36个月抚育(M3)、之前36个月以上无抚育(M4)。根据调查数据确定每种植物的最大株高。以小样方的土著物种个数作为土著物种丰富度。将盖度0~1%、1%~5%、5%~25%、25%~50%、50%~75%、75%~95%、95%~100%转换成1~7级多度[17],用R-FD包计算每个小样方的多度加权优势最大株高,它是样方内林下植被潜在最大株高与多度乘积的加和[18]。
1.5 数据分析
用SPSS 24.0的图表构建器进行数据的初步探索,有意义的关联使用均值比较或一般线性模型单因素分析进一步量化,事后多重比较方差齐性时用LSD法、非齐性时用T2法。为了寻找多种因素影响外来植物入侵的最佳子集,将可能对入侵产生影响的所有因子进行SPSS自动线性建模,包括林龄、林冠郁蔽度、优势最大株高、植被总盖度、土著物种丰富度、是否路边、抚育时间、分区等8个因子。之后用R-lme4量化最佳子集对入侵的影响,以分区为随机截距,最佳子集其他因子为解释变量建立混合效应模型,逐步简化模型至所有项都显著[19]。调用effects包allEffects函数提取混合效应模型中各项的效应值,用plot(allEffects(model))命令绘制互作效应面板图;当提取混合效应模型中某一项的效应时,其他各项处于其各水平样本数加权平均值[20];在R3.5.2[21]上进行混合效应模型分析、互作效应提取和作图。
2 结果与分析
老虎岭林区共调查240个植被样方,记录到171种植物,隶属于116科和162属,外来植物24种,土著植物147种;其中禾本科植物最多,有21种,其次是菊科15种;外来植物丰富度样方均值为1.35,盖度均值26.0%;土著植物丰富度样方均值4.54,盖度均值39.6%。样方中出现的外来植物包括9种一年生草本、9种多年生草本和6种木本植物。9个林内分组的林冠郁蔽度估算总均值53%,按降序排列为B区红锥、D区西桦、A区马尾松、E区麻栎、E区国外松、C区邓恩桉、B区尾巨桉、D区任豆、F区斑皮桉,分别为86%、71%、62%、60%、58%、55%、47%、26%和10%,T2多重比较结果为a、ab、b、abcd、abc、b、cd、de和e。15个分组的优势最大株高平均值分别为:B区尾巨桉2.00 m、E区麻栎1.79 m、C区邓恩桉1.76 m、A区马尾松1.63 m、D区任豆1.47 m、E区国外松1.22 m、B区红锥1.19 m、路边C 1.09 m、路边A 1.06 m、路边E 1.00 m、路边D 0.91 m、路边B 0.90 m、D区西桦0.81 m、F区斑皮桉0.76 m、路边F 0.56 m。
2.1 不同分组的外来植物入侵程度比较
外来植物入侵对15个分组的一般线性模型单因素分析方差非齐性,从T2多重比较和均值95%置信区间可知,F区外来植物占比最高;除F区斑皮桉和E区麻栎之外,路边的外来植物入侵显著高于其他所有林内分组(P<0.05);斑皮桉是引进树种且林下外来植物入侵较多,但同为引进树种的尾巨桉和国外松中外来植物入侵很少,邓恩桉林下完全没有外来植物(见图1),因此并未发现引进树种造林与乡土树种造林的林下外来植物入侵有差异。图1中15个分组的外来植物占比估算均值从上到下分别为90.1%、62.5%、61.2%、57.5%、50.2%、49.1%、37.5%、19.7%、19.5%、5.2%、5.0%、4.5%、3.1%、2.2%和0。F区的路边和林内入侵均较高,外来植物分布可能与分区关联较与造林树种关联更密切,路边多于林内。
2.2 林龄和抚育时间是否显著影响外来植物分布格局
研究区林分包括6个林龄段:国外松和麻栎40 yr,马尾松22 yr,邓恩桉9 yr,任豆、红锥和西桦6 yr,尾巨桉和斑皮桉3 yr,路边0 yr;6个林龄段的外来植物占比估算均值分别为11.3%、4.5%、0、9.0%、34.5%和59.8%;T2多重比较的结果为BC、C、C、C、B和A,路边的外来植物入侵极显著多于其他所有林龄段分组(P<0.01),其次是3 yr林分和40 yr林分。外来植物入侵随林龄的变化是非线性的。
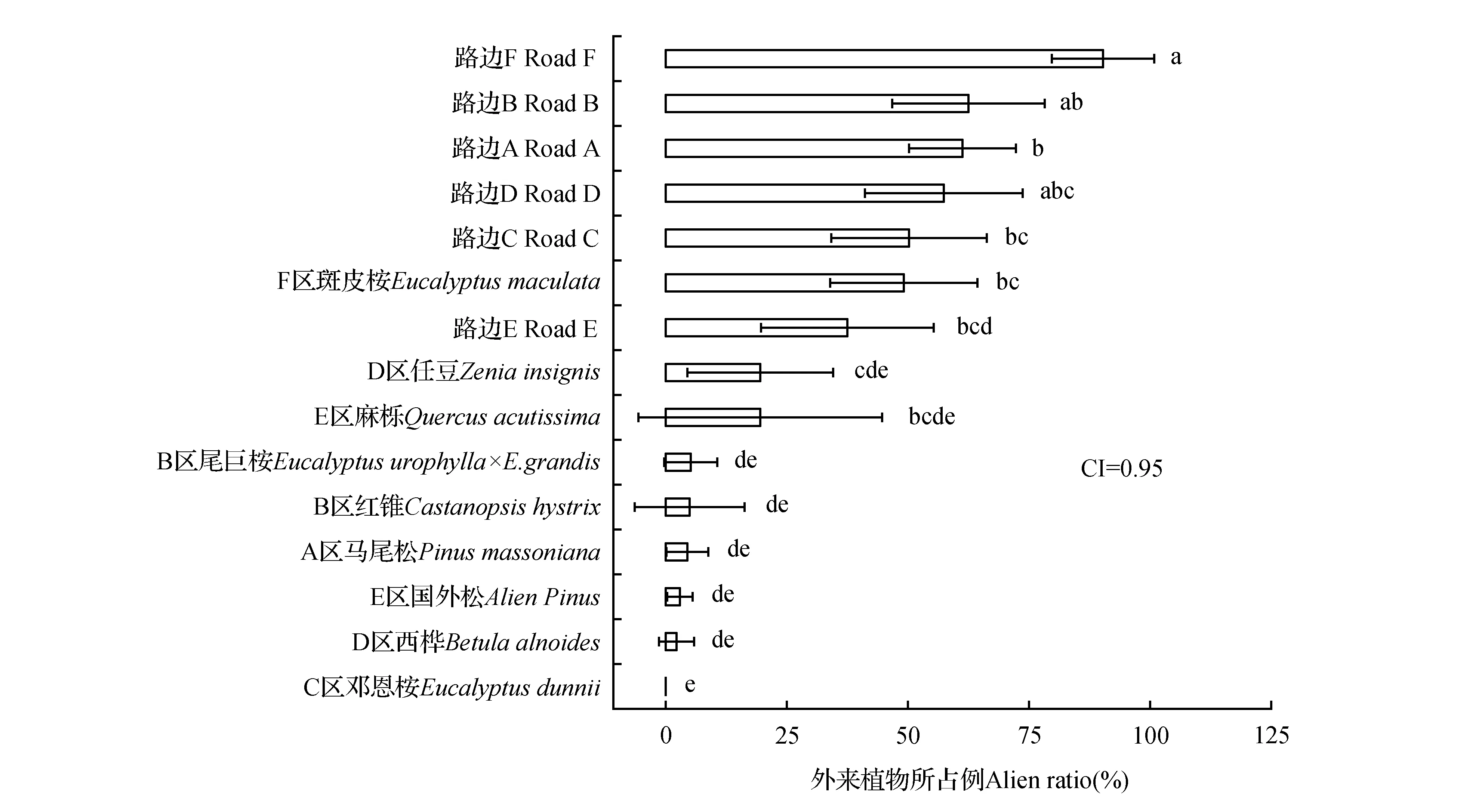
图1 不同分组的入侵平均值、95%置信区间和SPSS一般线性模型T2多重比较结果 T2多重比较无相同字母表示在P<0.05水平上差异显著。Fig.1 Means,95% confidence intervals and T2 multiple comparison results of invasions in 15 groups calculated by SPSS GLM-GEN Not sharing letter implied significant difference at P< 0.05 level.
之前4~6个月抚育(M1)、之前6~20个月抚育(M2)、之前20~36个月抚育(M3)、之前36个月以上无抚育(M4)等4种抚育时间的外来植物占比估算均值分别为33.0%、37.7%、4.5%和47.1%,4种抚育时间的入侵差异极显著(F=35,df=5,P=0.000),多重比较表明M3与其他3种抚育时间的入侵差异极显著,其他3种之间无显著差异;只有A区马尾松抚育时间为M3。虽然只比较抚育时间单个因子的方差分析表明不同抚育时间的入侵差异极显著,从图1中可见B区红锥、B区尾巨桉、D区西桦和E区国外松的外来植物入侵与A区马尾松的不相上下,但M1、M2和M4的其他组分拉高了它们的平均值;只比较单个因子均值、只分析主效应可能产生误导的结论。
2.3 植被样方位置对入侵的影响
植被样方位置高于林道、齐平林道、低于林道对外来植物入侵影响极显著(n=240,F=43,df=2,P=0.000),估算入侵均值分别为6.1、53.6和22.5,T2多重比较结果C、A和B,3种位置之间入侵差异均达极显著。我们用SPSS“选择个案”屏蔽所有路边样方,只对120个林内样方进行样方位置单因素的一般线性模型分析,样方位置对入侵仍然影响极显著(n=120,F=7,df=2,P=0.001),估算入侵均值分别为5.3、5.0和21.5,T2多重比较结果B、B和A;低于林道林内样方的外来植物入侵极显著高于其他两种位置的林内样方。
2.4 多因素的混合效应模型分析
以外来植物入侵为目标变量、其他有关因子为预测变量,利用SPSS自动线性建模从多种因素中筛选出解释目标变量的最佳子集;中选模型准确性为65.2%,最佳子集包括是否路边、林冠郁蔽度、土著物种丰富度、分区、林龄、优势最大株高,分别占解释率的37%、21%、19%、10%、7%和6%。以分区为随机截距,最佳子集的其他因子为解释变量建立混合效应模型,简化后的最小适当模型见表1。在分区效应为平均截距、其他固定效应处于其均值的情况下,路边对入侵的主效应为正且极显著(P=0.000),是否路边的外来植物入侵相差42.5%;林冠郁蔽度对入侵主效应为负(P=0.000),林冠郁蔽度每增加10%,外来植物入侵减少8.4%;样方的优势最大株高对入侵的主效应也为负,样方的优势最大株高每增加21 cm,外来植物占比减少5%(见表1)。
表1 以入侵为响应变量、分区为随机截距、SPSS自动线性建模最佳子集为固定效应的混合效应模型分析结果
Table 1 Result of mixed effect model with alien ratio as response variable,minimum subset produced by SPSS automatic linear modeling as fixed effects,and district as random intercept
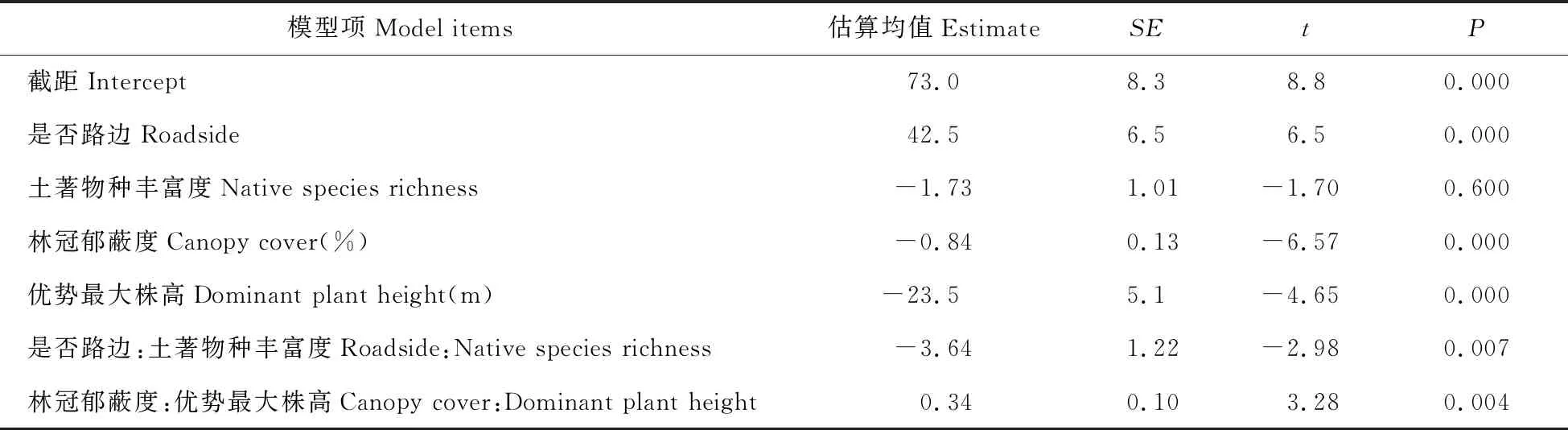
模型项Model items估算均值EstimateSEtP截距Intercept73.08.38.80.000是否路边Roadside42.56.56.50.000土著物种丰富度Native species richness-1.731.01-1.700.600林冠郁蔽度Canopy cover(%)-0.840.13-6.570.000优势最大株高Dominant plant height(m)-23.55.1-4.650.000是否路边:土著物种丰富度Roadside:Native species richness-3.641.22-2.980.007林冠郁蔽度:优势最大株高Canopy cover:Dominant plant height0.340.103.280.004

图2 混合效应模型中的两个互作效应 A.是否路边*土著物种丰富度;B.林冠郁蔽度*优势最大株高Fig.2 Two interactions in the mixed effect model A.Roadside*Snative(Native species richness); B.Canopy cover*Heightmax(Dominant plant height(m))
混合效应模型中包含两个极显著的互作项。在是否路边和土著物种丰富度对入侵的互作效应中(见图2a),当模型中其他项的效应为其均值时,不论土著物种丰富度多少,都呈现路边入侵多于林内的趋势。从互作效应提取数值中查看定住是否路边时外来入侵随土著物种丰富度的变化趋势;先看在林内的情况,当土著物种丰富度为0、3、6、9和10时,入侵为28.3%、23.1%、17.9%、12.8%、11.0%;再看路边的情况,土著物种丰富度为0、3、6、9和10时的入侵分别为70.8%、54.7%、38.6%、22.4%、17.1%;路边土著物种丰富度明显提升入侵抵抗性,但林内土著物种丰富度只能微弱增加入侵抵抗性;而当土著物种丰富度处于研究区均值(4.54)时,林内入侵约为20.5%,路边入侵约为46.4%,相差约26%。
在林冠郁蔽度和优势最大株高对入侵的互作效应中,当优势最大株高较矮时(0.2~1.0 m),随着林冠郁蔽度的减小入侵迅速增加;当样方的优势最大株高较高时(2 m),入侵随林冠郁蔽度的变化不大(见图2b)。当优势最大株高≥4 m,林冠郁蔽度越高外来植物入侵越多,但这一趋势的预测可靠性较低,因其置信区间较宽,可忽略不计。再从提取的互作效应数值中查看变化趋势;林冠郁闭度为0时,优势最大株高0.2、1.0和2.0 m对应的入侵分别为73.4%、54.6%和31.0%,负相关关系;林冠郁闭度为50%时,优势最大株高为0.2、1、2、4和5 m的入侵分别为34.7%、29.6%、23.2%、10.3%和3.9%,两者仍然负相关;当林冠郁闭度为70%,优势最大株高同样为0.2、1、2、4和5 m,外来植物入侵变为19.2%、19.6%、20.0%、20.9%和21.3%,几乎持平不变。
3 讨论
3.1 混合效应模型互作效应分析揭示共存因子的相互作用
利用混合效应模型明确地控制空间差异、具体地量化多因素的互作效应是本文的两大特色。由于不同分区在干扰、拓殖种繁殖体库、地形位置等方面有事先存在的差异,分区内的物种组成和植被结构存在自相关,入侵不仅与生物因子差异相关而且与区组作用相混淆,数量分析方法必需考虑影响变化的来源。本文的混合效应模型以分区作为随机效应,允许每个分区有不同的截距,以此控制来源于分区或造林树种的差别效应,多因素分析是在分区效应被均化后的结果。生物入侵研究目前仍难以提供可预测、可实用的知识,其中一个重要原因是大多数研究没有明确地控制空间差异[11]。
互作效应是一个熟悉的概念,但互作效应中各因子引起的响应变化方向一直是陌生而模糊的;本文借助专业软件的互作效应图形化和数据提取来解释互作效应。入侵研究通常分析单个环境因素的影响,忽略共存因素的互作效应,但实际上互作效应可能产生明显的非加性效应,导致意想不到的后果[22];本文是从互作效应中发现路边的土著物种丰富度明显提升入侵抵抗性,林内的土著物种丰富度只能微弱增加入侵抵抗性,以及林冠郁蔽度和优势最大株高在限制外来植物入侵上作用非可加。尽可能具体而清楚地了解的环境因子对外来入侵的互作效应,我们才能更有针对性地应对外来入侵威胁[23]。
3.2 模型解释结果与前人发现的比较
建立在系统设计的第一手调查数据基础之上的外来物种分布格局及其影响因子的研究目前还少见报道[24~25],较多前人研究是基于文献资料和非系统设计调查。我们从道路条件、物种多样性、林冠郁蔽度和优势最大株高、造林树种等几个方面,对比前人的研究结果来阐述本文的新发现。
道路是入侵植物进入自然生态系统的途径之一[26~27]。路边沟渠微生境往往有较好光照、水分、接触行走动物等条件,是入侵植物的最适宜定植点;土著植物通常对这些极端的条件比较敏感而不易定植,因此,路旁通常比周围地区分布更多的外来植物[6]。老虎岭林区的路边外来植物入侵显著高于林内;当其他因素效应处于其平均值时,路边入侵较林内入侵高26%;而且,不论土著物种丰富度增加或减少,均呈现路边入侵高于林内的趋势。于永辉等在广西的调查也发现林缘和林区道路的外来入侵植物种类最多[28];Wu等的研究结果显示在地区尺度上离公路距离与植物入侵显著相关[29]。除了认同路边与外来入侵密切相关这一科学共识之外,本文具体分析了相对于林道的样方位置高低对外来植物入侵的影响,发现在林道下方的林内样方较易被入侵,林道上方不易被入侵,与林道齐平的林内样方也不易被入侵;因此,在监测或防控林业外来植物时,重点应放在低于林道的森林。
物种多样性是一个地区生态系统健康稳定的基础[30~31]。通常认为土著物种丰富度高的地方对入侵的抵抗性强,但也有研究发现土著植物未对外来物种入侵发挥抵抗作用[23]或外来物种数量与土著种数量正相关[29]。混合效应模型分析中发现土著物种丰富度对入侵的主效应不显著,但土著物种丰富度与路边互作对入侵的效应极显著;在林区道路旁,土著物种丰富度强烈地抑制外来植物入侵;但在森林内部外来植物入侵普遍较少时,土著物种丰富度对入侵的抑制作用不甚明显。
混合效应模型中林冠郁蔽度和优势最大株高都极显著降低外来植物入侵。大多数小种子草本植物、树木和灌木在林冠郁闭情况下无法定植[32];稠密的林冠和较高的林下植被可阻挡种子空降,减少外来植物繁殖体压力;郁闭林分的建群种往往将资源利用得比较彻底,特别是光资源的利用;资源有限也使外来植物难以成功定植。Grime[32]将在较长时间尺度上才能产生的间接影响称为过滤器效应或奠基者效应,本文的林冠郁蔽度和优势最大株高是限制外来植物入侵的两大过滤器(filter),但两个因子的作用是非可加的,当林冠郁蔽度为零时优势最大株高对入侵的抑制效应强大,随着林冠郁蔽度的增加,优势最大株高对入侵的抑制效应减弱,当林冠郁蔽度达70%时优势最大株高对入侵的抑制效应几乎为零;反之,当优势最大株高较矮时,林冠郁蔽度对入侵的抑制效应明显,而优势最大株高≥2 m时林冠郁蔽度对入侵的抑制效应为零或转变为促进作用。
造林树种是群落优势种、建群种,优势种对群落结构和群落环境的形成有明显的控制作用,对生态系统的直接效应在很大程度上由优势种的性状和功能多样性决定[32]。本文的9个林内分组包含针叶林、干燥硬叶林、常绿阔叶林和落叶阔叶林;从单因素分析中可见,9种林内分组可分为高入侵(斑皮桉)、中等入侵(任豆、麻栎)、低入侵(尾巨桉、红锥、马尾松、国外松和西桦)和无入侵(邓恩桉)4个类型,两个极端都是干燥硬叶林,没有表现出哪类树种较易被入侵,引进树种造林与乡土树种造林对比在林下外来植物入侵方面也无明显差异。
本文以样方所有外来植物的相对百分比作为入侵程度指标,通过模型解释发现:当其他因素效应处于其平均值时,路边入侵较林内入侵高26%;样方相对于林道的位置高低影响入侵程度,林道下方的样方较易被入侵;土著物种丰富度对入侵的主效应不显著,但与路边的互作对入侵效应极显著;林冠郁蔽度和优势最大株高是限制外来植物入侵的两大因素,对入侵的主效应和互作效应均显著,但这两个因子对入侵的作用非可加;林龄和抚育时间对外来植物入侵的影响趋势不明显。
