大丽轮枝菌β-1,4-内切木聚糖酶基因的鉴定及生物信息学分析
2020-06-04张迎春孙天歌张小雪梁其干李艳军
张迎春,孙天歌,张小雪,梁其干,李艳军,孙 杰
(石河子大学 农学院,新疆生产建设兵团绿洲生态农业重点实验室,新疆石河子 832003)
黄萎病被称为棉花的“癌症”,是制约中国棉花生产的主要因素之一。中国棉花黄萎病主要是大丽轮枝菌(Verticilliumdahliae)和黑白轮枝菌(Verticilliumalboatrum)侵染所致。大丽轮枝菌是一种土传性病原真菌,可侵染200多种植物,其寄主主要是双子叶植物[1-2]。大丽轮枝菌的微菌核能在土壤中存活30~50年,当感应到寄主植物根际释放的根系分泌物后,微菌核开始萌发产生感染性菌丝可定植于根皮层,进而扩散到木质部产生孢子,导致作物的维管束受到阻塞,出现枯萎症状,发育迟缓,黄化和坏死[3-5],严重影响棉花的产量和品质。因中国气候变化多样,大丽轮枝菌在不同环境条件下产生了许多具有丰富遗传多样性的生理小种,田间致病性易发生变异,出现强致病力的类型[6-7],大丽轮枝菌的防治目前仍无有效的生物及化学措施。大丽轮枝菌全基因组数据的公布为其致病分子机制的研究提供了基础,为抗病新品种的开发提供了新路线[8]。
植物细胞壁由纤维素、半纤维素以及果胶等多糖组成,这些组分与木质素一起形成致密的网状结构,形成植物的支撑结构[9]。在大丽轮枝菌侵染植物过程中分泌大量的细胞壁降解酶(cell wall degrading enzymes, CWDEs)[10]。细胞壁降解酶不仅可降解植物细胞壁中的多糖,而且也破坏植物细胞壁结构,为病原微生物的入侵和摄取营养提供条件[11]。细胞壁降解酶包括角质酶、多聚半乳糖醛酸酶、果胶酯酶、果胶裂合酶、木聚糖酶、内葡聚糖酶G1、β-1,4-葡萄糖苷酶和蔗糖非发酵蛋白激酶等[11]。角质酶VdCUT11水解植物细胞壁的角质层促进大丽轮枝菌的侵染[12]。果胶裂解酶VdPL1可诱导植物免疫反应,同时作为一个毒力因子参与大丽轮枝菌的侵染[13]。除角质酶和果胶裂解酶外,纤维素酶、聚半乳糖醛酸酶、木聚糖酶等在大丽轮枝菌侵染植物的过程中也发挥了重要作用[13]。
植物细胞的感染需要半纤维素的降解[14],木聚糖是植物细胞壁中半纤维素的主要成分。在苹果树烂腐病研究中,木聚糖酶被证明与致病性或毒力有关[15]。在灰霉病菌研究中,木聚糖酶基因xyn11A缺失突变体的致病力显著降低,但未完全丧失致病力[16]。木聚糖酶属于水解酶类,主要包括有β-1,4-内切木聚糖酶、β-1,4-外切木聚糖酶、β-1,4-木糖苷酶、α-L-呋喃阿拉伯糖苷酶、α-葡萄糖醛酸酶和乙酰木聚糖酯酶等[17]。其中,β-1,4-内切木聚糖酶是使木聚糖完全水解最关键的酶,其作用于木聚糖主链的木聚糖苷键,随机切断主链内的糖苷键,水解木聚糖而生成分子量大小不同的寡糖[18]。β-1,4-内切木聚糖酶能使组成细胞壁的不溶性木聚糖变成可溶性木聚糖,从而迅速降解细胞壁[19]。在病原菌侵染植物的过程中,β-1,4-内切木聚糖酶参与了细胞壁的降解,表明它在致病过程中发挥着重要作用[20]。然而,目前大丽轮枝菌中木聚糖酶研究仍较少,本研究从大丽轮枝菌基因组中鉴定β-1,4-内切木聚糖酶基因并进行生物信息学分析,同时对这些基因在棉花不同抗感品种根系分泌物中的表达模式进行分析,旨在揭示β-1,4-内切木聚糖酶基因与大丽轮枝菌致病性的联系,为进一步研究β-1,4-内切木聚糖酶基因在大丽轮枝菌致病过程中的作用以及揭示致病的分子机制奠定基础。
1 材料与方法
1.1 材料与处理
大丽轮枝菌V991菌株由石河子大学绿洲生态农业重点实验室保存,2个陆地棉品种‘新陆早7号’(感黄萎病)和‘中植棉2号’(耐黄萎病)均由石河子大学棉花研究所提供。
1.1.1 棉花根系分泌物的收集 将‘新陆早7号’和‘中植棉2号’品种的种子浸泡在1%(1 g NaClO加入100 mL H2O)NaClO溶液中进行表面消毒,用无菌蒸馏水反复冲洗后种在经过高温高压(121 ℃,60 min)灭菌处理的沙土中。每个品种种植2盆,每盆9株苗,光周期为16 h/8 h,温度28 ℃,每3 d用霍格兰营养液浇灌1次。 45 d后,从沙土中取出棉苗,用2 L灭菌水浸泡砂土,使根系分泌物充分溶解在水中,用细菌过滤器 (0.22 μm)过滤水溶液,在冷冻干燥机中浓缩至0.5 L。
1.1.2 大丽轮枝菌的培养与取样 将贮存的大丽轮枝菌V991分生孢子培养于察氏培养基上,25 ℃黑暗条件下培养5 d。收集新鲜的分生孢子,分别称取0.5 g分生孢子接菌于5 mL不同抗感棉花品种的根系分泌物中。25 ℃ 220 r/min摇培0、6和12 h后,收集V991的分生孢子。根系分泌物培养前0 h的大丽轮枝菌样本,标记为CK;感病品种‘新陆早7号’根系分泌物培养6 h和12 h的样本,分别标记为VDX6和VDX12;耐病品种‘中植棉2号’根系分泌物培养6 h和12 h的样本,分别标记为VDZ6和VDZ12;同时用水培养大丽轮枝菌,培养6 h和12 h的样本分别标记为VDW6和VDW12。每处理2个重复用于转录组测序,共构建14个文库。
1.2 方 法
1.2.1 大丽轮枝菌β-1,4-内切木聚糖酶基因的鉴定 从http://fungi.ensembl.org/Verticillium_dahliae/Info/Index/ 下载大丽轮枝菌全基因组编码序列CDS(Coding sequence) GCF_000150675.1_ASM15067v2-rna序列文件、GCF_000150671_ASM15067v2-protein 蛋白质序列文件、GCF_000150675.1_ASM15067v2_ genomic全基因组序列文件,以及Verticillium_dahliaeASM15067v2.42 GFF3文件。利用TBtools v 0.66443软件对GFF3文件中所有β-1,4-内切木聚糖酶基因进行搜索,同时在CDS、蛋白质、全基因组序列文件中获取基因对应的cDNA序列、蛋白质序列以及基因组序列。
1.2.2 β-1,4-内切木聚糖酶基因的氨基酸序列分析 利用ExPAsy(http://web.expasy.org)工具在线分析β-1,4-内切木聚糖酶基因一级结构的基本理化性质,包括氨基酸序列大小、分子质量和等电点等。利用在线工具WOLFPSORT(https://wolfpsort.hgc.jp)和 TMHMM(http://www.cBMs.dtu.dk/services/TMHMM/)分别对编码蛋白质的亚细胞定位情况和跨膜结构域进行预测[21]。
1.2.3 多重序列比对和进化树分析 采用DNAMAN 6.0和ClustalX 2.0对β-1,4-内切木聚糖酶氨基酸序列进行多重序列比对。利用SMART工具在线预测11个β-1,4-内切木聚糖酶蛋白序列的信号肽和结构域,并用DOG 1.0软件进行绘制。以11个β-1,4-内切木聚糖酶蛋白序列的主结构域为探针序列,在CaZy数据库中(http://www.cazy.org/)查找其他物种中的相应蛋白[22],去除重复以及不完整序列后,挑选9条其他物种中的β-1,4-内切木聚糖酶蛋白序列用于进化树分析。利用MEGA 6.0提供的ClustalW程序对大丽轮枝菌及其他物种中的β-1,4-内切木聚糖酶基因进行多重序列比对,然后通过邻位相连法构建基因的复合进化树。
1.2.4 β-1,4-内切木聚糖酶基因的染色体分布 从大丽轮枝菌全基因组数据库中获得β-1,4-内切木聚糖酶基因染色体分布以及长度信息,利用TBtools v0.66443软件绘制β-1,4-内切木聚糖酶基因的染色体分布图。
1.2.5 β-1,4-内切木聚糖酶基因结构分析 从全基因组数据库下载该基因的外显子和内含子信息,利用在线软件Gene Structure Display Server (GSDS)(http://gsds.cBMi.pku .edu.cn/)分析大丽轮枝菌β-1,4-内切木聚糖酶基因外显子-内含子的数目及分布。
1.2.6 大丽轮枝菌样本RNA的提取及转录组测序 采用RNA simple total kit试剂盒(Tiagen,中国北京)提取大丽轮枝菌总RNA,利用Nanodrop○R2000分光光度计(Thermo Scientific,Wilmington,de,USA)和Agilent 2100生物分析仪(Agilent TeGHnologies,Santa Clara,CA,USA)分别检测RNA的纯度和完整性,利用Qubit○R2.0荧光计(Thermo Scientific,Wilmington,de,USA)对RNA的浓度进行精确测量,最终获得高质量的RNA样本用于RNA测序。采用Illumina HiSeqTM4000测序平台进行转录组测序,由北京诺禾致源科技股份有限公司完成,转录组数据已上传至NCBI SRA数据库中,登录号为PRJNA545805。
1.2.7 β-1,4-内切木聚糖酶基因对不同棉花抗感品种根系分泌物的响应分析 对转录组测序获得的raw reads进行质量控制后,利用HISAT软件将得到的clean reads比对到大丽轮枝菌的参考基因组数据(http//www.broadinstitute.org/annotation/genome/Verti- cillium dahlia /Blast.html)。采用HTSeq软件对各样品进行基因表达水平分析,使用的模型为union,分别统计单个基因的表达水平。FPKM(fragments per kilobase of exon per millon fragments mapped)为每百万fragments中来自某一基因每千碱基长度的fragments数目,同时考虑测序深度和基因长度对fragments计数的影响,是目前最为常用的基因表达水平估算方法[22]。根据转录组测序结果获得β-1,4-内切木聚糖酶基因的FPKM值,并采用HemI软件绘制基因在不同抗感棉花品种根系分泌物中的表达热图,使用Excel 2010制作基因表达折线图。
2 结果与分析
2.1 大丽轮枝菌β-1,4-内切木聚糖酶基因的鉴定
通过TBtools软件查找GFF3文件中所有 β-1,4-内切木聚糖酶基因,共获得11条关于β-1,4-内切木聚糖酶基因的记录,分别为VDAG_03385~VDAG_08273(表1)。这些基因编码序列(coding sequence, CDS)长度为666~1 314 bp,编码221~437个氨基酸,蛋白质分子质量为23.90~47.64 ku,理论等电点为5.09~10.00。对各成员序列的亚细胞定位预测表明,这些基因大多(7个)为细胞膜外蛋白,部分定位于细胞膜、线粒体、细胞质基质和细胞核中(表1)。其结果与角毛壳菌生物信息学分析一致,说明该酶蛋白主要在胞外发挥水解木聚糖的生物学作用[23]。
2.2 大丽轮枝菌β-1,4-内切木聚糖酶的保守域分析
利用蛋白跨膜结构域在线分析工具TMHMM(http://www.cBMs.dtu.dk/services/)发现,β-1,4-内切木聚糖酶蛋白不含跨膜结构域(transmembrane domain, TM)。利用在线工具SMART(http://smart.embl-heidelberg.de/smart/batGH.pl)对11个β-1,4-内切木聚糖酶蛋白序列进行信号肽和结构域预测,分析发现11个酶中 VDAG_03790、 VDAG_05042、 VDAG_05043、 VDAG_05294和 VDAG_06165均含有糖苷水解酶Glyco_hydro11结构域,其中 VDAG_06165还含有一个fCBD(cellulse biding domain;真菌纤维素结合域)结构域; VDAG_08273、 VDAG_03385、 VDAG_03809和 VDAG_04744均含有糖苷水解酶Glyco_hydro10结构域; VDAG_05509和 VDAG_03808均含有糖苷水解酶 Glyco_Hydro 43结构域,其中 VDAG_03808还含有一个CBM(carbohydrate binding modules;碳水化合物结合域)结构域(图1)。VDAG_03385、 VDAG_08273、 VDAG_04744、 VDAG_06165V、DAG_03809和 VDAG_05509含有不同长度的信号肽(图1)。
将 VDAG_03790、 VDAG_05042、 VDAG_05043、 VDAG_05294和 VDAG_06165蛋白的Glyco_hydro11结构域进行多重序列比对,发现GGKGW、YGW、EYY和STR区域具有较强的保守性(图2-A)。将 VDAG_08273、 VDAG_03385、 VDAG_03809和 VDAG_04744蛋白的Glyco_hydro10的结构域进行多重序列比对,保守区域为WDVVNE、ADP、NDY和TELD(图2-B)。对VDAG_03808和VDAG_05509蛋白的Glyco_Hydro 43结构域进行多重序列比对,KGVRD和IATDL区域具有较强的保守性(图2-C)。这些保守性区域可能与木聚糖酶的催化相关[24-25]。
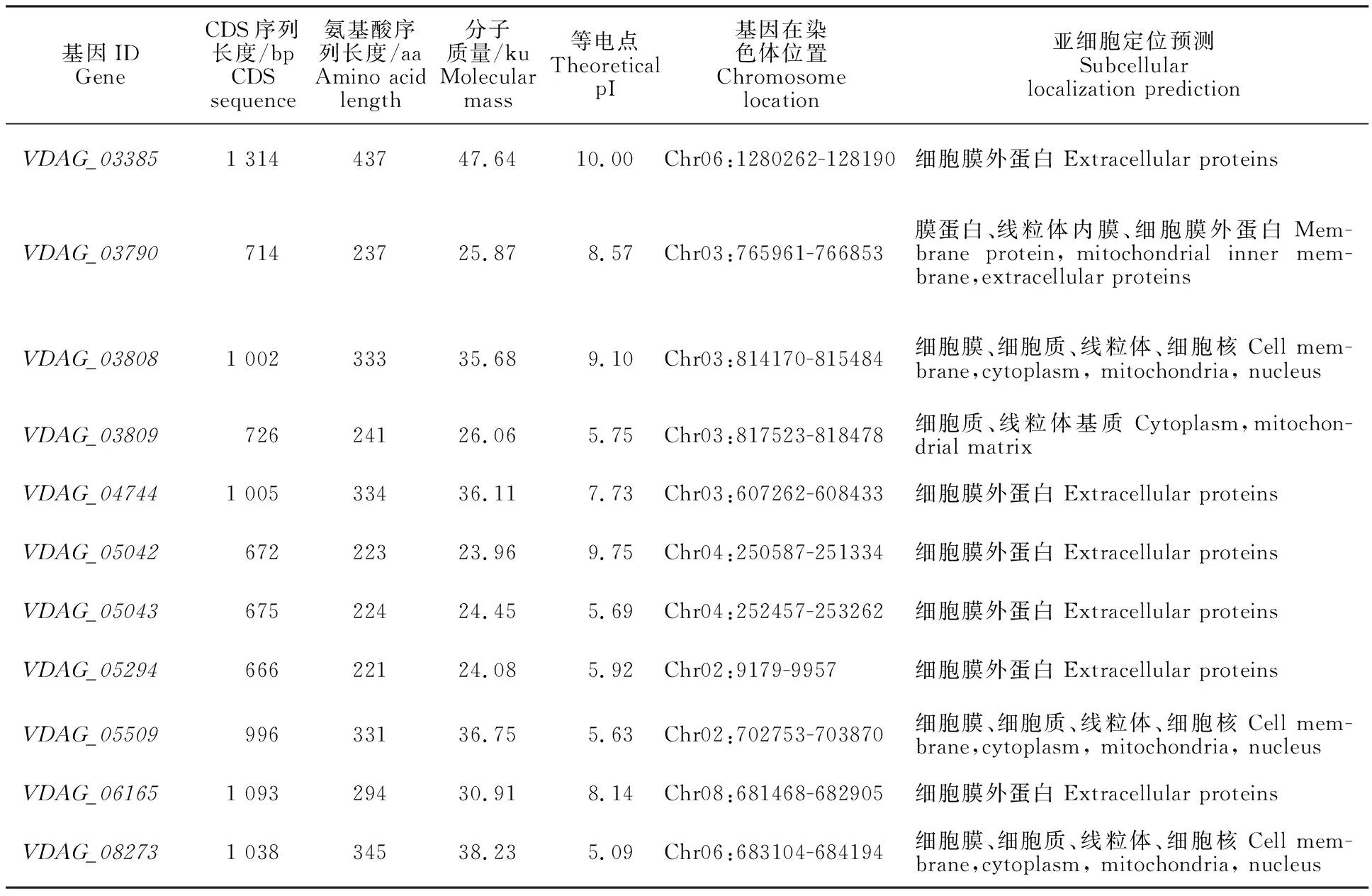
表1 大丽轮枝菌β-1,4-内切木聚糖酶基因的特征Table 1 Characterization of Endo-β-1,4-xylanase genes in V.dahliae
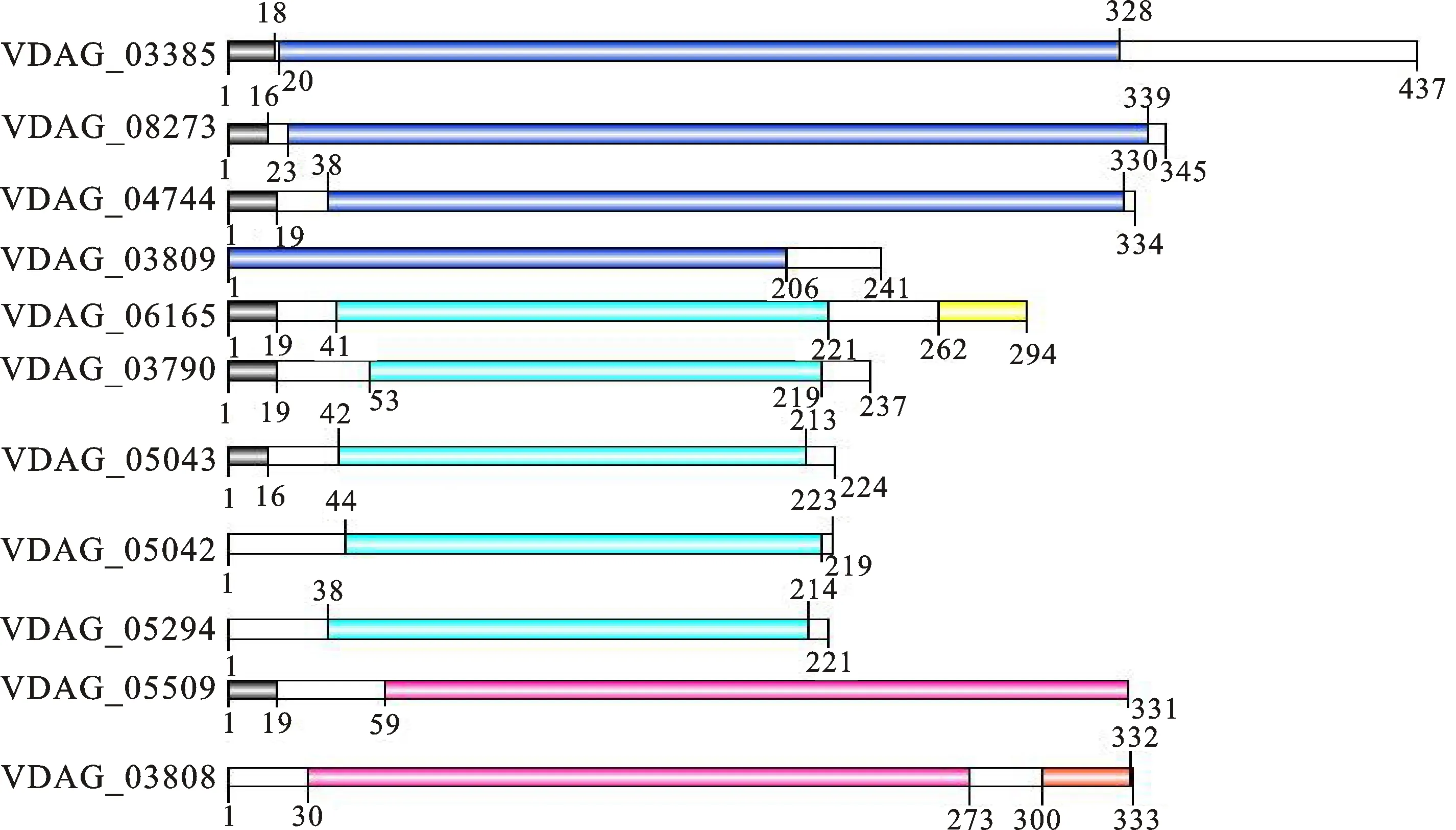
黑色表示信号肽;深蓝色表示Glyco_hydro10结构域;浅蓝色表示Glyco_hydro11结构域;粉色表示Glyco_Hydro 43结构域;黄色表示fCBD结构域;橙红色表示CBM结构域
The black indicates signal peptide;Dark blue indicates the Glyco_hydro10 domain; Light blue indicates the Glyco_hydro11 domain; Pink indicates the Glyco_Hydro 43 domain; Yellow indicates the fCBD domain;Orange red indicates the CBM domain
图1 大丽轮枝菌β-1,4-内切木聚糖酶蛋白结构域分析
Fig.1 Domains analysis of Endo-β-1,4-xylanase proteins inV.dahliae

A为VDAG_03790、VDAG_05042、VDAG_05043、VDAG_05294和VDAG_06165蛋白Glyco_hydro11结构域多重序列比对;B为VDAG_08273、VDAG_03385、VDAG_03809和VDAG_04744蛋白Glyco_hydro10结构域多重序列比对;C为VDAG_03808和VDAG_05509蛋白Glyco_Hydro 43结构域多重序列比对。灰色和黑色区域分别代表 75%以上和 100%的保守区。黑色方框表示氨基酸保守区域
A indicates a multiple sequence alignment of Glyco_hydro11 domain in VDAG_03790, VDAG_05042, VDAG_05043, VDAG_05294, and VDAG_06165 proteins; B indicates a multiple sequence alignment of Glyco_hydro10 domain in VDAG_08273, VDAG_03385, VDAG_03809 and VDAG_04744 proteins; C indicates a multiple sequence alignment of Glyco_Hydro 43 domain in VDAG_03808 and VDAG_05509 proteins.The gray and black areas represent more than 75% and 100% of conservative regions, respectively.Black boxes indicate regions of amino acid conservation
图2 大丽轮枝菌β-1,4-内切木聚糖酶蛋白结构域多重序列比对
Fig.2 Multiple sequence alignment of domains in Endo-β-1,4-xylanase proteins fromVerticilliumdahliae
2.3 大丽轮枝菌β-1,4-内切木聚糖酶蛋白的进化关系分析
在CaZy数据库中查找并随机选取9个来自其他物种的β-1,4-内切木聚糖酶蛋白记录,其中XP_018153852.1(十字花科炭疽病菌)、EPS36346.1(捕食线虫真菌)、ABR14629.1(宇佐美曲霉)和RGP60782.1(拟枝孢镰刀菌)属于糖苷水解酶(glycoside hydrolase,GH)10家族成员,BAE71410.1(黑色素短梗霉)、ACS96449.1(嗜酸真菌)和ACR83565.1(黑曲霉)属于11家族成员,XP_013423968.1(出芽短梗霉)和THY53895.1(出芽短梗霉)属于43家族成员。利用MEGA 6.0软件将上述9个蛋白与11个大丽轮枝菌β-1,4-内切木聚糖酶蛋白进行进化关系分析,结果如图3所示,20个糖苷水解酶基因被清晰地分为3组。 VDAG_06165、 VDAG_03790、 VDAG_05294、VDAG_0 5042和 VDAG_05043与其他物种中11家族成员聚为一类; VDAG_08273、 VDAG_03385、 VDAG_03809和 VDAG_04744与其他物种中10家族成员聚为一类; VDAG_05509和 VDAG_03808与其他物种中43家族成员聚为一类。该结果与结构域预测结果相一致,表明大丽轮枝菌11个β-1,4-内切木聚糖酶蛋白中,含有5个糖苷水解酶11家族成员,4个10家族成员和2个43家族成员。
2.4 大丽轮枝菌β-1,4-内切木聚糖酶基因结构分析
利用在线软件GSDS对获得的11个β-1, 4-内切木聚糖基因的结构进行分析,结果如图4所示:VDAG_06165基因外显子和内含子最多,含有7个外显子和6个内含子;VDAG_05042和VDAG_08273基因外显子和内含子较少,分别含有2个外显子和1个内含子。其余8个基因中,4个基因(VDAG_03385、VDAG_03809、VDAG_04744和VDAG_03808)含有4个外显子和3个内含子,4个基因(VDAG_05294、VDAG_05043、VDAG_05042和VDAG_05509)含有3个外显子和2个内含子。

10、11和43分别指糖苷水解酶10、11和43家族
10、11 and 43 refer to glycoside hydrolase families of 10、 11 and 43, respectively
图3 大丽轮枝菌和其他物种中β-1,4-内切木聚糖酶蛋白进化树分析
Fig.3 Phylogenetic tree analysis of Endo-β-1,4-xylanase proteins inV.dahliaeand other species
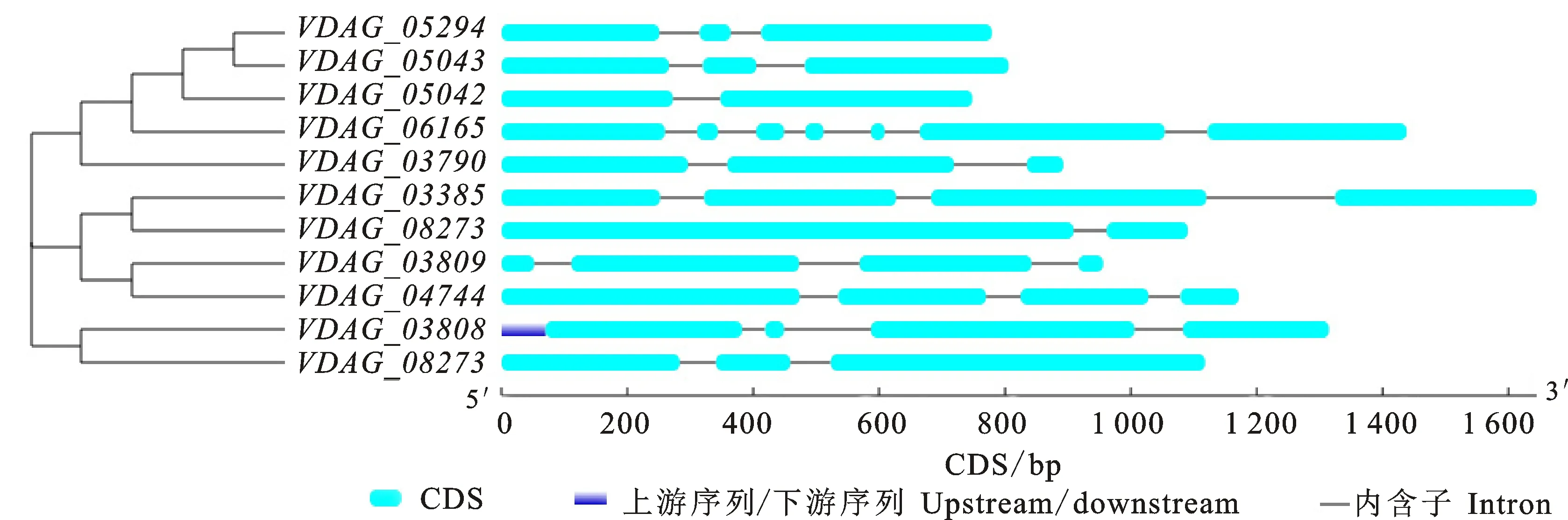
蓝色表示UTR(上下游序列),绿色表示编码区,灰线表示内含子区
The blueindicates UTR, the green indicates the exon, and the gray indicates the intron
图4 大丽轮枝菌β-1,4-内切木聚糖酶基因结构分析
Fig.4 Gene structures analysis of Endo-β-1,4-xylanase genes inV.dahliae
2.5 大丽轮枝菌β-1,4-内切木聚糖酶基因的染色体分布
大丽轮枝菌有8条染色体,Chr01长度为 5 243 713 bp,Chr02为5 264 584 bp,Chr03为 5 534 790 bp,Chr04为3 754 323 bp,Chr05为 3 249 347 bp,Chr06为2 537 490 bp,Chr07为 3 147 721bp,Chr08为2 984 914 bp。11个β-1,4-内切木聚糖酶基因分布在其中5条染色体上(图5)。3号染色体上分布的β-1,4-内切木聚糖酶基因最多,包括VDAG_03790、VDAG_03808、VDAG_03809和VDAG_04744四个基因,它们的基因组序列长度分别为893 bp、1 315 bp、956 bp和1 172 bp。8号染色体上分布的基因最少,仅含VDAG_06165基因,其基因组长度为1 438 bp。2号、4号和6号染色体上分别分布2个基因,VDAG_05294与VDAG_05509分布于2号染色体上,它们的基因组序列大小分别为779 bp和1 118 bp;VDAG_05042和VDAG_05043分布于4号染色体上,它们的基因组序列大小分别为748 bp和806 bp;VDAG_03385和VDAG_08273分布于6号染色体上,它们的基因组序列大小分别为1 645 bp和1 091 bp。根据200 kb的核苷酸中含有3个以上基因即为1个基因簇[26],3号染色体上的4个基因形成1个基因簇,其他染色体上未形成基因簇。
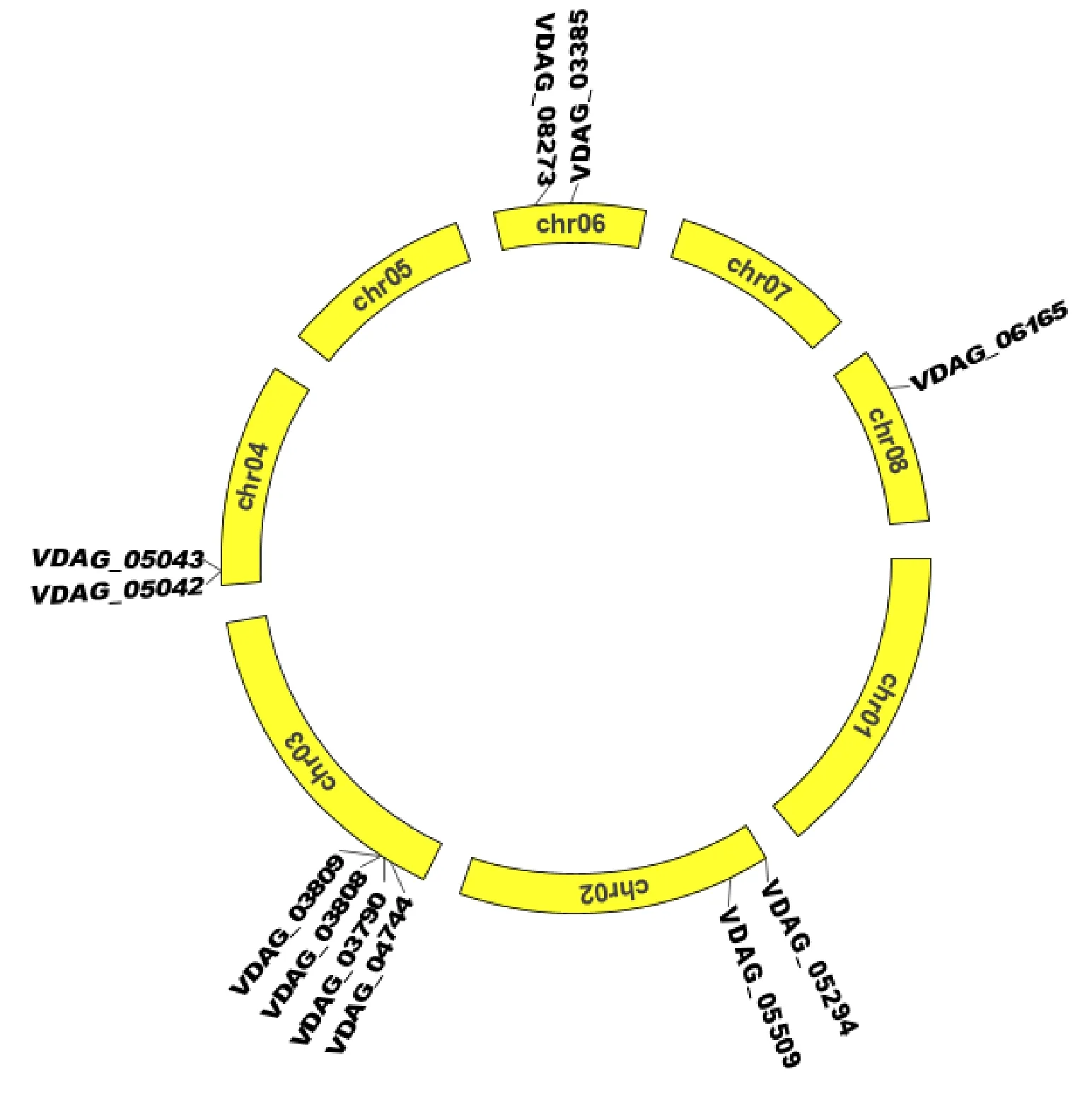
图5 大丽轮枝菌β-1,4-内切木聚糖酶基因在染色体上的分布Fig.5 Chromosomal distribution of Endo-β-1,4-xylanase genes in V.dahliae
2.6 β-1,4-内切木聚糖酶基因对不同抗感棉花品种根系分泌物的响应分析
抗病品种根系分泌物对病菌的孢子萌发和菌丝生长有一定的抑制作用,而感病品种根系分泌物则能刺激病菌生长[27-28]。根据不同抗感棉花品种根系分泌物培养6 h和12 h的大丽轮枝菌的转录组测序结果,获得β-1,4-内切木聚糖酶基因培养后每个时间点的FPKM值,用HemI软件绘制基因在不同抗感棉花根系分泌物中的表达热图。根据β-1,4-内切木聚糖酶基因在不同抗感根系分泌物中表达模式的差异(图6),11个基因被划分为3组,第3组基因(VDAG_03790、VDAG_03808、VDAG_03809、VDAG_05043和VDAG_06165)的表达量在感病品种根系分泌物培养的大丽轮枝菌(VDX6和VDX12)中明显较高,而在耐病品种根系分泌物培养的大丽轮枝菌(VDZ6和VDZ12)和水培养的大丽轮枝菌(VDW6和VDW12)中表达量较低。结合5个基因的FPKM折线图(图7)可见,VDAG_03790在感病品种根系分泌物培养6 h的样本中表达量明显升高;VDAG_03808在感病品种根系分泌物培养6 h和12 h的样本中表达量均较高,VDAG_03809、VDAG_05043和VDAG_06165在感病品种根系分泌物培养12 h的样本中表达量明显升高。而5个基因在耐病品种根系分泌物和水培养的样本表达量未见明显升高。上述分析表明5个基因均对感病品种根系分泌物产生响应,暗示它们与大丽轮枝菌的致病性相关。

红色和蓝色分别代表基因表达水平的高低
Red and blue bands represent high and low gene expression levels, respectively
图6 β-1,4-内切木聚糖酶基因在不同抗感棉花
根系分泌物培养的大丽轮枝菌中的表达热图
Fig.6 Heat map of Endo-β-1,4-xylanase genes
inV.dahliaecultured root exudates
from different resistant cottons
3 结论与讨论
糖苷水解酶(glycoside hydrolase,GH)亦称糖苷酶,是作用于各种糖苷或寡糖使糖苷键水解的酶的总称。CAZy(Carbohydrate-Active enzymes Database)数据库以糖苷水解酶与唯一性底物特异性结合原理,将糖苷水解酶家族分为130个家族[29-30]。内切木聚糖酶主要存在于两个离散的序列家族第10和11家族[31]。本研究通过对大丽轮枝菌中11个β-1,4-内切木聚糖酶基因进行生物信息学分析发现,5个基因含有Glyco_hydro11结构域,与其他物种中11家族成员聚为一类,属于糖苷水解酶11家族;4个基因含有Glyco_hydro10结构域,与其他物种中10家族成员聚为一类,属于10家族;2个基因含有Glyco_hydro43结构域,与其他物种中43家族成员聚为一类,属于43家族。
蛋白结构域分析还发现VDAG_06165中含有一个fCBD结构域,该结构域被认为存在于大多数真菌纤维素酶和半纤维素酶中[32]。CBD结构域在木聚糖酶降解天然植物细胞壁木聚糖中起着重要的作用[32-33],它是由33~36个氨基酸组成的具有高度保守性的区域。由于VDAG_06165含有fCBD结构域,推测该基因可能与降解木聚糖有关。VDAG_03808中含有一个CBM结构域,该结构域是碳水化合物结合模块(CBMS),通常存在于较长的蛋白质序列中,与碳水化合物活性酶的催化模块(例如糖苷水解酶和多糖裂合酶)共存,在那里它们可增强酶模块的催化活性[34]。CBM通过使酶接近纤维素表面来影响酶的活性[30,34]。

VDX表示β-1,4-内切木聚糖酶基因在感病棉花品种根系分泌物中表达量;VDZ表示β-1,4-内切木聚糖酶基因在抗病棉花品种根系分泌物中表达量;VDW表示β-1,4-内切木聚糖酶基因在水培养中表达量
VDX represents the expression level of Endo-β-1,4-xylanase genes inV.dahliaecultured with root exudates from susceptible cultivars;VDZ represents the expression level of Endo-β-1,4-xylanase genes inV.dahliaecultured with root exudates from tolerance cultivars;VDW represents the expression level of Endo-β-1,4-xylanase genes inV.dahliaecultured with water
图7 β-1,4-内切木聚糖酶基因在不同抗感棉花品种根系分泌物培养的
大丽轮枝菌中FPKM值的变化趋势图
Fig.7 Change trend of FPKM value of Endo-β-1,4-xylanase genes inV.dahliae
cultured with root exudates from different resistant cottons
糖苷水解酶43家族成员均含有Glyco_hydro43结构域,催化果胶降解的果胶酶一般含有该结构域[25]。本研究中VDAG_03808和VDAG_05509均含有糖苷水解酶43家族结构域,推测这两个基因可能参与果胶的降解。在所有含有糖苷水解酶43主结构域的蛋白质中,69%的蛋白质还含有一个信号肽,将翻译后的蛋白质导向细胞质外。VDAG_03808中无信号肽;VDAG_05509含有1~19 bp长度分泌信号肽,推测该基因编码的分泌蛋白在细胞外发挥作用。
根系分泌物是植物根系在生长过程中释放到介质中的全部有机和无机物质。大丽轮枝菌通过根系入侵棉花,因此根系分泌物的生物学效应对大丽轮枝菌的成功侵染至关重要[28,35-36]。棉花根系分泌物富含氨基酸和糖类,抗病品种和感病品种的根系分泌物成分有明显差异。与抗病棉花品种根系分泌物相比,感病品种根系分泌物中富含天冬氨酸、苏氨酸、赖氨酸和脯氨酸等大量氨基酸,葡萄糖、果糖和蔗糖含量明显较高[37]。抗病品种根系分泌物抑制大丽轮枝菌的生长,而感病品种根系分泌物促进其生长[22,28]。目前棉花根系分泌物对基因表达的影响研究仍较少,VdRGS1在根系分泌物处理的大丽轮枝菌中的表达量明显升高,基因敲除和回补试验发现该基因与大丽轮枝菌孢子生产、菌丝发育、微菌核的形成和致病性相关[13]。本研究分析不同抗感棉花品种根系分泌物对11个β-1,4-内切木聚糖酶基因的表达的影响。
植物病原真菌可以产生一系列细胞壁降解酶,以促进感染和定植,包括纤维素酶、半纤维素酶、果胶酶等。近年来,研究人员已利用基因敲除手段对大丽轮枝菌中细胞壁降解相关基因的功能进行研究,但由于这些基因都是以基因家族的形式存在,导致基因的功能研究未获得确定性的结果。因此,确定基因家族中对致病性起关键作用的基因对于大丽轮枝菌致病分子机制的解析是十分必要的。本研究通过制作11个基因在不同抗感棉花根系分泌物中的表达热图和部分基因的表达折线图,发现5个β-1,4-内切木聚糖酶基因在感病品种根系分泌物培养的大丽轮枝菌中明显上调,而在耐病品种根系分泌物和水培养的样本中未明显上调,表明它们对感病品种的根系分泌物有明显的响应,暗示它们可能与大丽轮枝菌的致病性相关,这些基因可用作今后利用基因敲除技术研究基因功能与大丽轮枝菌致病分子机制的目标基因。
