越橘品种‘北陆’各器官发育过程中内源激素含量变化研究*
2020-05-16刘海楠孙海悦李亚东
刘海楠,谷 雨,孙海悦,李亚东
(吉林省小浆果遗传育种与创新利用工程研究中心,吉林农业大学园艺学院,长春 130118)
越橘为杜鹃花科越橘属(Vaccinium)植物,是一种具有较高经济价值的小浆果,其果实中含有丰富的营养物质,是多年生的灌木小浆果类果树。越橘的树体差异较明显,果实大小、形状、色泽各异。其口感细腻,甜酸适度,浆汁丰富,果实可加工可鲜食[1-3]。随着人们生活水平的提高,市场上对越橘果实的需求量越来越大,其栽培面积也在逐年增加,同时人们对果实的品质要求也越来越高。植物内源激素是植物正常代谢的产物,是在植物体内合成的,并能从产生部位转移到作用部位,在低浓度下就能调节植物生长发育的小分子有机化合物质。植物内源激素可以通过很多水平来起作用,包括激素合成、代谢、转运、感受和传导[4],从影响细胞的分裂、伸长、分化到影响植物发芽、生根、开花、结实、性别的决定、休眠和脱落等[5-7]。
关于植物激素的研究一直是植物科学前沿和热点领域之一[8]。国内外关于植物内源激素的研究数量有很多,在其他果树上研究得比较广泛,在越橘上仅在果实和花芽上有所研究,如李艳芳[9]、方仲相[10]、宋阳[11]及Oh[12]等人研究表明,在越橘果实中,脱落酸(ABA)能提高越橘果实果糖和葡萄糖的含量,促进果实软化、成熟和花色苷的积累并促进果实的显色,低浓度的ABA 能促进越橘花芽萌发,并且ABA 含量低的品种更容易成花。而生长素(IAA)抑制糖类物质和花色苷的合成,并且低浓度的IAA 能够促进越橘的花芽分化。玉米素(ZT)的含量在越橘果实快速生长期达到最高,并发挥其促进细胞分裂的作用。赤霉素(GA3)对糖类物质及花色苷的合成积累调控作用不显著,但是低浓度的赤霉素能够促进越橘芽由营养生长向生殖生长转化。本试验是对越橘地上部和地下部的器官发育过程中激素含量的变化进行研究,更加全面地分析植物激素对整个越橘植株的影响。
1 材料与方法
1.1 试验材料与来源
试验材料选自吉林农业大学浆果园,10 年生的越橘品种‘北陆’不同时期的花、果实、花芽、新梢、叶片及须根。
1.2 材料的采集
从春季撤防寒土开始取样直至秋季落叶为止,选取15 株长势一致的健康树,每5 株树为1 个重复,取样时分东、西2 个方向进行,每次每种样品取10 g,样品用冰盒带回实验室,经预处理后用液氮处理放入-40 ℃冰箱保存。
(1)花。从越橘花芽芽鳞松动开始取样,每7 d 取1 次样,取到花冠筒完全膨大开放为止。
(2)果实。从越橘花后3 d 即幼果期开始采集,每3 d 取1 次样,取到果实成熟。
(3)花芽。从果实落果开始取样,每7 d 取1次样,到落叶期为止,取样部位是选取停长的枝条上花芽,从顶芽往下数1~5 个芽位。
(4)新梢。从新梢生长到5 cm 长开始取样,每7 d 取1 次样,到落叶期为止,取样部位是取上部5 cm 长的新梢,去掉叶片及芽。
(5)叶片。从展叶期开始取样,每7 d 采1 次样,到落叶期为止,取样部位从顶端往下数1~9片叶片,洗净擦干液氮速冻,-40 ℃保存。
(6)须根。从越橘展叶期开始取样,每14 d取1 次样,到落叶期为止,取样部位为距离树主干20 cm 的位置向下挖30 cm 深,取0~30 cm 处的须根,用水将须根上的泥土冲洗掉,滤纸吸干水,液氮速冻,-40 ℃保存。
金矿石常采用磨矿—浮选—浮选金精矿细磨—氰化浸出常规工艺获取成品金[1]。而大部分金矿中含有低品位的有色金属元素,如铅、铜等,其价值大约是金价值的10%或者更高,采用常规工艺无法有效回收这些共伴生有价元素[2]。调研走访得出企业不愿综合回收的主要原因有两方面:(1)在金价高位运行时期,企业重视金产量,忽视铅、铜等伴生元素的回收研究工作,不愿投入时间精力研发、改造;(2)在金价低迷时期,企业又无力进行流程优化、改进,因此造成了资源的极大浪费。
1.3 越橘各器官内源激素的提取与测定
1.3.1 激素的提取
(1)浸提。称取2 g 样品,在冰浴下研磨成浆,加入80%预冷甲醇20 mL,保鲜膜密封,4 ℃冰箱浸提16 h(注意避光)。
(2)去甲醇。浸提液10 000 r/min(4 ℃)离心10 min,取上清液,向残渣中加入20 mL 80%冷甲醇,10 000 r/min(4 ℃)离心10 min,合并上清液丢弃残渣。将上清液在40 ℃下减压蒸发至没有甲醇残余(约剩余5 mL)。将剩余水相转移到三角瓶中。
(3)萃取脱色。分别用10 mL 石油醚萃取脱色3 次弃去醚相,加入0.01 g PVPP,超声波振荡30 min,抽滤取上清(常温13 000 r/min离心10 min),再分别用10 mL 乙酸乙酯萃取3 次,合并酯相。将酯相在40 ℃条件下减压蒸发至干。
(4)定容。再用甲醇(色谱纯)溶液溶解并定容2 mL,经0.45 μm 微孔滤膜过滤得到待测液,保存在4 ℃冰箱中。
1.3.2 内源激素HPLC 检测
HPLC 检测所需的色谱条件与周荧等[13]在高效液相色谱测定蓝莓叶片内源激素的方法研究中的条件一致。
1.3.3 标准溶液的配制与测定
分别称取0.005 g 的ABA、IAA、GA3和ZT 的标准品,分别用甲醇溶解配制成50 mg/L 的母液,过0.45 μm 的有机相滤膜。将ABA、IAA、GA3和ZT 在1.3.2 的色谱条件下进样,记录保留时间,以此来定性;将标准母液稀释并混合进样,通过标准曲线法定量。
1.4 数据处理与分析
试验数据采用SPSS 23.0 分析软件和Microsoft Excel 2010 应用程序处理分析。
2 结果与分析
2.1 越橘花生长发育过程中内源激素含量变化
从图1 中可以看出,越橘花中ABA 呈先上升后下降的变化趋势,在5 月4 日即芽鳞分离这一时期出现峰值,然后先下降后逐渐趋于平稳。IAA 的含量变化趋势与ABA 的含量变化趋势完全相反,在芽鳞分离这一时期即5 月4 日出现最小值,随后又开始上升,在单花完全分离时出现峰值后又下降。GA3呈先上升后下降再上升的变化趋势,在花冠筒完全膨大时期即5 月22 日出现最大值。ZT 含量的变化呈先上升后缓慢下降的趋势,但ZT 的含量在花的整个发育过程中均处于较高水平,在5 月4 日出现峰值。
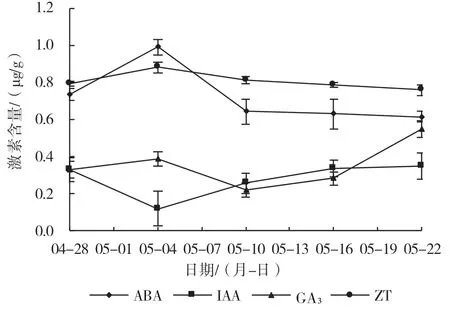
图1 ‘北陆’花中内源激素含量的变化
2.2 越橘果实生长发育过程中内源激素含量变化
从图2 可以看出,在果实的整个发育过程中4种内源激素含量均呈现先下降后上升再下降的趋势。在坐果期,ZT 和IAA 含量较高,此时果实处在第1 个快速生长期,在坐果后果实迅速膨大,随着果实的增大,ZT、IAA 和GA3的含量都逐渐降低,在6 月9 日降到最低值,果实进入缓慢生长期。在第2 次快速生长前期,果实内的ZT、IAA 和GA3含量迅速上升,在6 月27 日达第2 个峰值,随着果实的逐渐成熟,3 种内源激素的含量总体呈下降趋势。从图中可以看出,ABA 一直维持在较高的水平,在果实发育过程中,ABA 含量出现2 次峰值,在花后第6 d 即5 月28 日出现1 次峰值,第2 次峰值出现在花后第42 d 即7 月3 日,此时果实进入粉果期向果实的成熟期过渡。
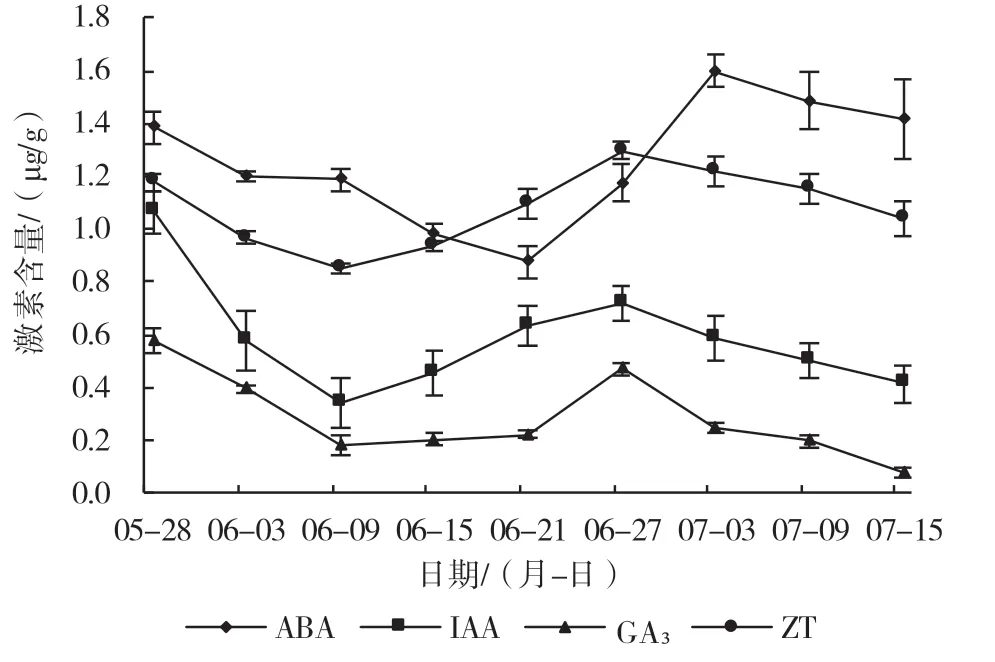
图2 ‘北陆’果实中内源激素含量的变化
2.3 越橘花芽中内源激素含量变化
在花芽分化过程中,4 种内源激素的含量变化如图3 所示,IAA 和ABA 的含量整体高于ZT 和GA3含量,IAA 的含量变化趋势是先下降后上升,ABA的含量呈先下降然后迅速上升后趋于平稳状态,ZT的含量在8 月9 日出现1 个高峰,在其他时期都处于较平稳的一个趋势,GA3的含量变化是先上升(在8 月9 日达到最大值),然后开始下降,后又开始上升,然后保持平稳的趋势。
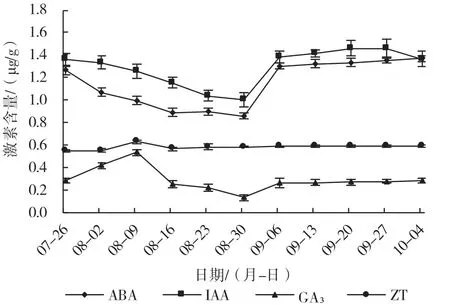
图3 ‘北陆’花芽中内源激素含量的变化
2.4 越橘新梢中内源激素含量变化
从图4 可以看出,在新梢生长过程中,IAA、GA3、ZT 含量变化趋势均呈双S 变化曲线,而ABA的含量则呈先下降后上升之后又逐渐下降的平缓变化趋势。5 月中旬IAA、GA3、ZT 含量较低,之后随着新梢逐渐生长,IAA、GA3、ZT 含量逐渐上升,在5 月底达到较大值,之后又经过迅速下降的过程后缓慢上升,在8 月底IAA、GA3、ZT 含量达到第2 个峰值,而ABA 的含量一直呈下降趋势,此时新梢处于旺盛生长期。之后IAA、GA3、ZT 含量迅速下降后维持在一个较低水平,而ABA 逐渐上升后又下降,此时新梢生长速度相对较慢甚至停止生长。
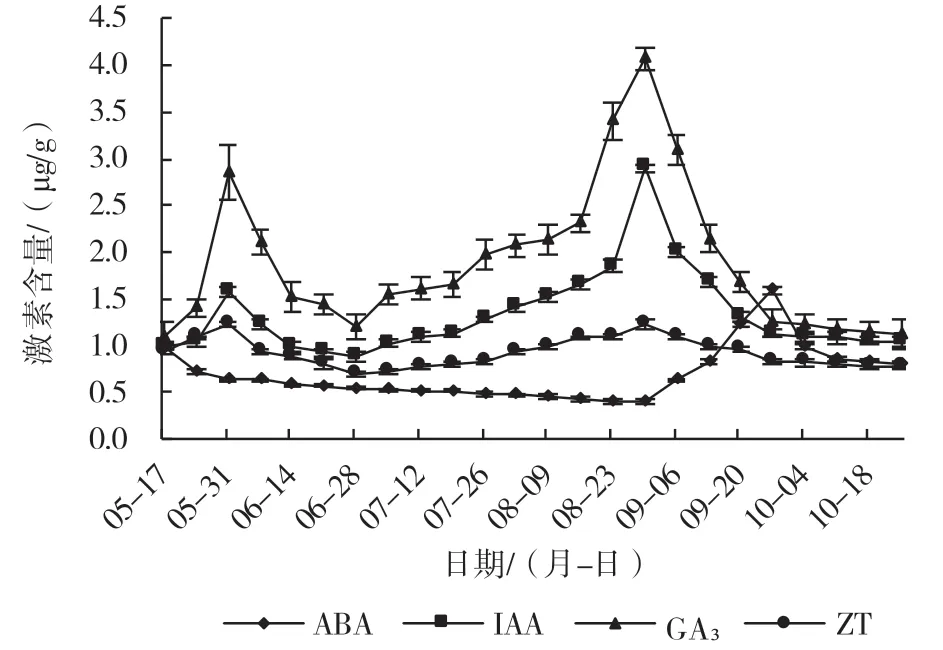
图4 ‘北陆’新梢中内源激素含量的变化
2.5 越橘叶片中内源激素含量变化
从图5 可以看出,4 月下旬展叶时IAA、GA3、ZT 含量相对较低,之后随着叶片面积的逐渐增长,IAA、GA3、ZT 含量逐渐上升,在6 月初达到峰值,此后IAA 含量逐渐下降,而GA3和ZT 含量迅速下降,到6 月底下降速度减慢,7 月底GA3、ZT 又开始迅速回升,到8 月底9 月初达到第2 个高峰,后缓慢下降至趋于平稳。ABA 含量在展叶初期相对较高,之后随着叶片的不断增长,其含量持续下降,到6 月初降至较低点,后逐渐上升,7 月初达到峰值后下降,7 月底降至较低后逐渐上升,此时恰为果实采收期。
2.6 须根中内源激素含量变化
须根的生长发育直接影响地上部器官的生长发育,由图6 可以看出,须根生长发育过程中IAA、GA3、ZT 含量均呈双S 形变化曲线,ABA 含量变化与其相反。5 月10 日地上部展叶开始随着地上部分逐渐增长,须根中IAA、GA3、ZT 的含量也逐渐上升,在6 月7 日出现第1 个峰值,随后又开始下降,在7 月19 日出现较低值,而ABA 的含量出现较高的峰值,此时地上部分的果实已进入成熟期,之后IAA、GA3、ZT 3 种激素含量又逐渐上升后开始缓慢下降,在8 月底出现第2 次峰值,此时地上部分生长缓慢甚至停止生长。

图5 ‘北陆’叶片中内源激素含量的变化
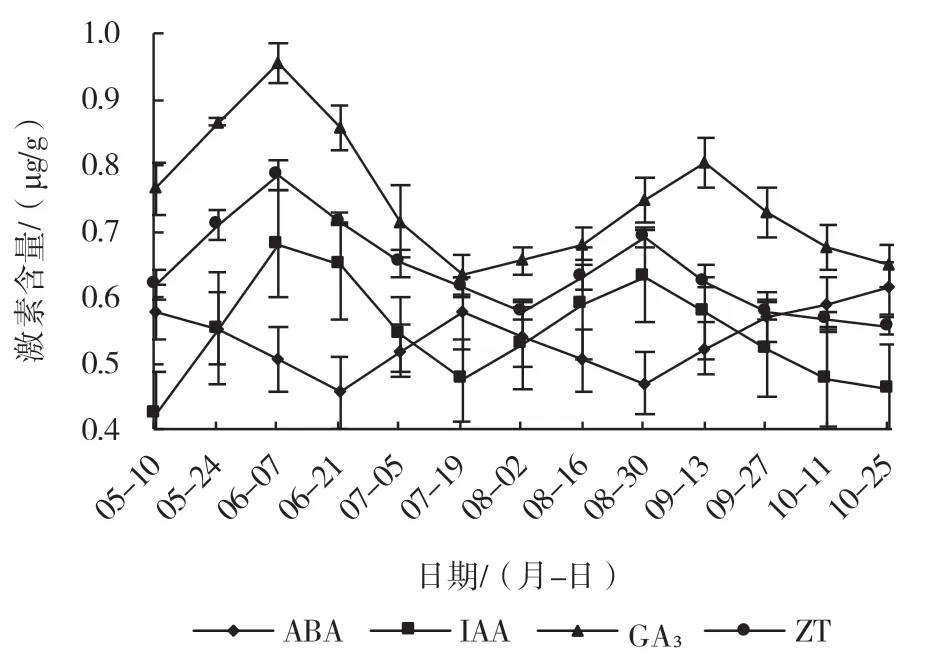
图6 ‘北陆’须根中内源激素含量的变化
3 结论与讨论
3.1 越橘各器官内4 大内源激素含量的变化趋势
有关植物内源激素的研究很早就有很多报道,在越橘上也有一定的研究,但是对整株越橘各个器官中的激素变化尚未有人研究。前人研究表明,植物的生长发育离不开植物内源激素的调节[14],本试验通过运用HPLC 法对越橘品种‘北陆’的花、果实、花芽、新梢、叶片及须根中ABA、IAA、GA3、ZT 等植物激素进行检测。
本试验研究的花中4 种激素的含量变化趋势与郭辰等[15]在东兴金花茶花果期内源激素动态变化研究结果的相同之处是:IAA 和GA3整体都处于上升趋势,ABA 呈先上升后下降的趋势,而ZT 的含量变化趋势完全不同,本试验中ZT 的含量整体处在一个平稳的较高水平,而郭的研究中ZR 含量整体呈下降趋势,可能是树种不同所导致,虽然ZT和ZR 都属于细胞分裂素,但是属于2 种化合物,这可能是导致结果不同的根本原因。在越橘果实中的4种激素含量变化趋势是ABA呈先下降再上升然后缓慢下降后趋于平稳,GA3和IAA 在果实发育整个过程中均呈下降趋势,并且GA3一直处于较低水平,ZT 含量变化是呈先下降再上升再下降的趋势,GA3和ZT 的含量在果实发育中期较高,IAA 含量仅在早期较高,这一试验结果与李艳芳[16]在研究越橘品种‘日出’和‘喜来’果实中的内源激素含量变化研究结果一致。与赵利娟等[17]研究的蓝靛果忍冬果实中GA3、IAA、ZT 和ABA 的含量变化趋势也一致。本试验研究的花芽中的GA3、IAA 和ABA 含量变化趋势与宋杨等[11]在蓝莓不同品种花芽形成过程中内源激素变化的研究中5 种不同品种的蓝莓果实生长发育过程中的内源激素含量变化趋势基本一致,GA3的含量变化呈上升—下降—上升的趋势,IAA 的含量变化是呈先下降后又上升的趋势,ABA的含量变化是呈先下降再上升的趋势,而检测的细胞分裂素虽然与宋杨等研究结果不同,但是变化趋势基本相同。
本试验研究的越橘新梢中的GA3、IAA 和ABA的含量变化趋势与马海燕[18]在葡萄生长过程中内源激素含量变化研究中的趋势相同,二者研究的GA3和IAA的含量变化趋势均是上升—下降—上升—下降,ABA 含量变化是呈先下降再上升的趋势。在叶片中的GA3、IAA 和ABA 的含量变化趋势与高庆玉等[14]在树莓叶片生长过程中内源激素含量变化初步研究中相一致,二者GA3含量变化呈先上升再下降然后上升最后再下降的趋势,IAA 含量变化呈先上升后又下降的趋势,ABA 含量变化呈下降—上升—缓慢下降—缓慢上升后趋于一个平稳的趋势。本试验所研究的新梢和叶片中的细胞分裂素在其他树种上很少有研究,并且前人很少有对植物根系中的GA3、IAA、ZT 和ABA 含量的动态变化进行研究,在越橘上更是未见报道。本试验的这些研究可为未来在这方面的深入研究奠定理论基础。
3.2 越橘各器官内4 大内源激素的变化与品种发育的关系
本试验中花分为5 个时期分别是芽鳞松动期(4月28 日)、芽鳞分开期(5 月4 日)、花分开芽鳞脱落期(5 月10 日)、单花完全分离期(5 月16 日)和花冠筒完全膨大开放期(5 月22 日)。从芽鳞松动起开始到芽鳞分开期这段时期,ABA、ZT 和GA3的含量变化都是上升的趋势;IAA 则与其相反,从芽鳞分开到花分开芽鳞脱落这段时期,ABA、ZT 和GA3的含量呈下降状态,相反IAA 含量开始升高直至花冠筒完全膨大开放为止。从花分开芽鳞脱落开始,ZT 和ABA 的含量缓慢下降至花冠筒完全开放,而GA3含量从花分开芽鳞脱落时期开始升高至花冠筒完全膨大开放时达到最大值。IAA 和GA3对越橘花的开放可能有较大的促进作用。
根据前人研究[19],越橘果实在生长发育阶段大致可分为3 个阶段,分别为幼果期(第1 次快速生长期)、缓慢生长期、转色期和成熟期(第2 次快速生长期)。本试验的结果表明,在幼果期4 大内源激素的含量逐渐减少,而ABA 含量持续下降到果实缓慢生长期,在缓慢生长期之后,ABA 含量迅速上升,其他3 种激素含量在缓慢生长期呈上升趋势,在转色期,4 种激素含量又开始缓慢下降,在7 月15 日之后进入完全成熟期。在果实发育初期,4 种激素含量都较高,GA3和ZT 的含量较高有利于果实的细胞分裂和生长,IAA 的含量较高有利于果实坐果,ABA 含量出现1 个小高峰可能与花的脱落有关,在第2 次快速生长阶段前期,GA3、IAA 和ZT 含量处于较高水平,有利于果实的快速生长,在进入成熟期之前,高含量的ABA 有利于果实的成熟,这一研究结果与高庆玉等[20]在树莓果实激素研究和赵利娟等[17]在蓝靛果忍冬果实中激素研究的结果相一致。
根据周强[21]的试验结果,可以把越橘品种‘北陆’的花芽分化分为7 个时期,分别为未分化期(7月24 日)、分化初期(8 月1 日)、花序原基分化期(8 月9 日)、萼片原基分化期(8 月17 日)、花瓣原基分化期(8 月25 日)、雄蕊原基分化期(9 月2日)以及雌蕊原基分化期(9 月18—26 日)。在花芽不同的分化时期,激素含量的变化也有所不同,花芽中的IAA 和ABA 的含量变化趋势一致,均呈先下降后上升的趋势,从7 月26 日花芽未分化期开始到8 月30 日花瓣原基分化期,IAA 和ABA 的含量持续下降,之后迅速上升,从9 月6 日雄蕊原基分化期到花芽分化完成时,IAA 和ABA 的含量趋于平缓。ZT 含量的变化是从未分化期开始上升,在8 月9 日花序原基分化期出现峰值后再下降到8 月16 日萼片原基分化期,随后上升到8 月30 日花瓣原基分化期后再继续缓慢上升并保持在一个相对较低水平上。GA3含量从未分化期开始升高,在8月9 日花原基分化期达到峰值后下降到8 月30 日花瓣原基分化期,随后略有上升,在雄蕊原基分化期后一直保持在较低水平,推测较低含量的GA3有利于花芽的发育,这一试验结果与吴曼等[22]在‘红丽’海棠芽的研究中的结果相似。
据前人研究结果可知,越橘新梢在整个生长周期有2 次生长高峰[23-24],即春季生长高峰期和秋季生长高峰期。本试验所用的试材第1 次快速生长期是5 月中旬到6 月上旬,6 月末出现黑点期新梢停止生长,约2 周之后新梢开始第2 次快速生长,时间是7 月中旬到8 月末,9 月之后新梢开始缓慢生长直至10 月停止生长。新梢中的IAA、GA3和ZT的含量在2 次快速生长阶段都呈上升趋势,而ABA的含量在新梢生长阶段一直呈下降趋势,直到9 月初开始升高并一直处在较高水平直至停止生长。可推测高含量的ABA 加速了新梢的衰老。而叶片的生长趋势和新梢的生长是分不开的,在新梢旺长期叶片也迅速生长,在生长季达到高峰,夏末至秋季逐渐下降,叶片内的激素含量随着叶片的生长而有所变化,从叶片展叶期开始,整个旺长期GA3、IAA和ZT 的含量不断升高。在第2 个新梢旺长期,GA3和ZT 含量呈上升趋势,IAA 的含量变化在第1 次旺长期后一直处于下降趋势,而ABA 含量变化与IAA 完全相反。由此可见,高含量的GA3、IAA 和ZT 有利于叶片面积的扩大,促进叶片的生长,延缓叶片的衰老,高浓度的ABA 能够促进叶片的脱落。
前人研究结果[25]表明,越橘根系在生长周期内出现2 次生长高峰,分别在6 月和9 月,在第1 次根系快速生长阶段,ABA 含量变化呈先下降后又升高趋势;在根系第2 次快速生长阶段,GA3的含量是先升高到一个峰值后又开始下降,IAA 和ZT 的含量变化呈下降趋势,ABA 则与其相反,呈上升趋势。前人研究表明,植物根系中合成的激素向地上部运输,供地上部的生长发育,在根系的2 次旺盛生长发育阶段,地上的新梢也是旺长期,推测根系高含量的GA3、IAA 和ZT 对地上部有一定的影响,而且在地上部果实进入成熟期时须根中的ABA 含量也是升高的。
