盐胁迫对葡萄砧木‘3309C’叶片光抑制的影响*
2020-05-16高玉录仝亚军张成超孙永江杜远鹏
高玉录,仝亚军,张成超,孙永江,杜远鹏,翟 衡
(山东农业大学园艺科学与工程学院,泰安 271018)
我国盐渍土资源量多且分布广泛,盐碱地的改良利用是涉及农业可持续发展的重大问题,目前耐盐‘海水稻’已经进入产业规模示范阶段。葡萄是一种相对较耐盐的果树树种,也是盐碱地区农民喜欢种植的高效经济作物之一,而葡萄砧木在抵御逆境、病虫等方面发挥重要作用,利用砧木进行嫁接栽培已成为葡萄产业发展的必然趋势。
植物光合作用对逆境胁迫非常敏感,可以用作反映逆境胁迫的有效探针。因此,研究盐胁迫对葡萄砧木光合作用的影响,对揭示砧木的抗盐机制具有重要的理论意义。植物吸收的光能超过光合作用所能利用的限度而导致光合效率下降的现象叫光抑制,它包括光系统II(PSII)光抑制和光系统I(PSI)光抑制。通常认为PSII 是逆境胁迫光抑制的主要位点[1],PSII 最大光化学效率(Fv/Fm)的下降是PSII 发生光抑制的重要指标[2]。相比PSII,一般情况下PSI 不易发生光抑制,但研究表明盐胁迫情况下PSI 却是容易伤害位点[3-4]。植物环式电子传递(CEF)由于具有产生跨膜质子梯度并只合成ATP且无还原力(NADPH)的形成的特点,在短期盐胁迫过程中起到了重要作用[5],被认为是抵御逆境胁迫的重要防御机制[6-7],但缺少对长期盐胁迫下葡萄CEF 活性变化觃律的报道。本研究以葡萄优良抗性砧木‘3309C’为试材,对盆栽苗进行浇盐处理,模拟不同持续时间的盐胁迫,研究了植株光合特性对盐胁迫的响应,以期为葡萄抗盐栽培提供理论基础。
1 材料与方法
1.1 材料与处理
试验于2017 年4—9 月在山东农业大学园艺实验站葡萄园基地内进行,选取葡萄砧木品种‘3309C’1 年生苗为试验材料,春季种植到觃格为外口直径23.5 cm、内口直径20.1 cm、高14.0 cm、底部直径12.9 cm 的塑料盆中,基质为中性壤土、沙及有机肥按照的比例配制而成,待生长到6~7 片叶片时,选取长势一致的植株,用浓度为100 mmolL 的NaCl 溶液浇灌植株,每株浇1 L,每隔2 d 浇灌1 次,对照浇等量清水,每3 株为1个处理,重复3 次。
分别在浇灌处理前、处理7 d(浇灌4 次)和处理14 d(浇灌8 次)选择中部节位长势一致的叶片进行叶绿素荧光参数测定。
1.2 叶绿素荧光参数的测定
使用连续激发式荧光仪(Handy PEA,Hansatech,英国)测定暗适应下的叶片快速叶绿素荧光诱导曲线。Fo为最小荧光(20 μs);Fk为K 点(300 μs)的荧光;FJ为J 点(2 ms)的荧光;Fm为最大荧光即P 点的荧光。通过JIP-test 分析可得到如下参数:


使用Dual-PAM100 叶绿素荧光仪(Walz,Germany)进行处理的活体测定。首先对经过一晚上暗适应的叶片进行Fm的测定,然后将叶片置于471 μmol·m-2·s-1的光下进行充分光启动后测定光下荧光参数:
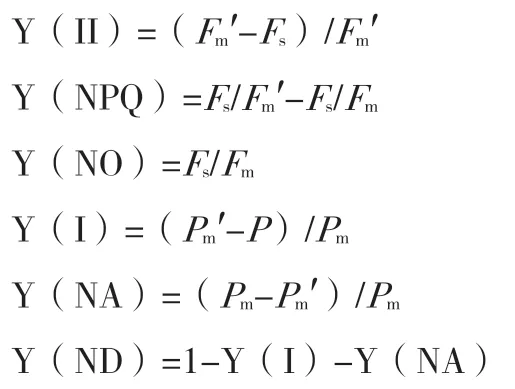
Y(II)表示PSII 的有效量子产率,Y(NPQ)和Y(NO)分别表示PSII 中调节和非调节能量耗散的量子产额,Y(NA)和Y(ND)分别表示由于PSI电子受体侧和供体侧限制引起的非光化学能量耗散的量子产额。Fm′表示光适应后的最大荧光,Fs是光适应后的稳态荧光,Pm为最大光氧化P700,Pm′表示光下PSI 最大荧光。

β/α-1 表示PSI 和PSII 间激发能分配的不平衡性[8]。
作用光关闭后叶绿素荧光瞬时上升测定:同样使用Dual-PAM 100 叶绿素荧光仪进行作用光关闭后叶绿素荧光瞬时上升的测定。选取长势一致的经过30 min 暗适应的叶片,测定开始后,先用小于0.1 μmol·m-2·s-1的调制测量光测定暗中最小荧光,然后再用大于7 000 μmol·m-2·s-1测定最大荧光,接着再用200 μmol·m-2·s-1活化光,此时荧光信号会随时间逐渐衰减,3 min 后关闭活化光,荧光信号降低后再次瞬时上升,同时继续记录此时荧光信号的变化。
P700+再还原速率的测定:利用Dual-PAM 100叶绿素荧光仪进行葡萄叶片P700+再还原速率的测定,经过30 min 暗适应的植物被远红光(>705 nm,5.2 μmol·m-2·s-1)照射使P700+充分氧化,经过30 s 照射后,荧光信号充分平稳。然后再关闭远红光使P700+再还原,并记录P700+再还原曲线的速率和时间,计算其半衰期(T1/2)。
1.3 数据分析
试验数据用SPSS 13.0 软件进行单因素方差分析,并进行LSD 多重比较。用Excel 进行数据分析以及作图,数据以平均值±标准误表示。
2 结果与分析
2.1 盐胁迫对PSII 及PSI 活性的影响
PSII 最大光化学效率可以反映PSII 受胁迫程度。从图1 可以看出,与对照相比,100 mmol/L 的NaCl 溶液处理到第7 d 时Fv/Fm开始出现降低,比对照下降了4.3%,第14 d 时出现显著下降,相比对照下降了13.6%。最大光氧化P700 可以反映PSI 活性,由图2 可知,胁迫处理第7 d 便导致了Pm值出现了显著降低,相比对照下降了14.7%,第14 d 下降了41.3%。上述结果表明,盐胁迫抑制了葡萄叶片PSII 和PSI 活性,而相比PSII,‘3309C’叶片PSI的盐胁迫耐受能力较弱。
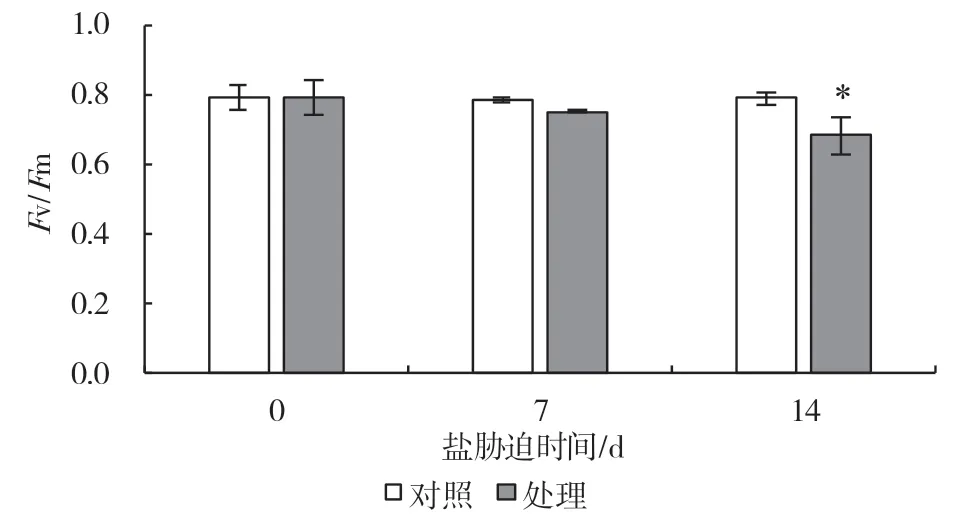
图1 盐胁迫对葡萄砧木叶片PSII 最大光化学效率的影响

图2 盐胁迫对葡萄砧木叶片最大光氧化P700 的影响
2.2 盐胁迫对叶绿素荧光曲线及参数的影响
通过分析叶绿素荧光多项上升动力学曲线可以获得与光合原初反应有关的丰富信息。从图3 可以看出,随着盐胁迫处理时间的延长,葡萄砧木叶片O-J-I-P 曲线发生变化。长期盐胁迫导致叶片荧光曲线发生明显变化,Fm值发生明显降低,说明盐胁迫下叶片单位面积吸收的光能明显降低。
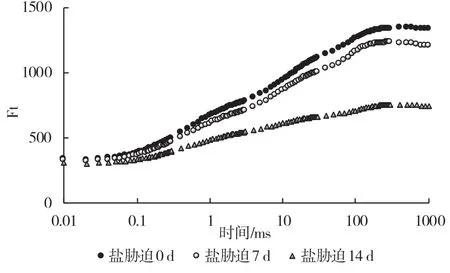
图3 盐胁迫下葡萄砧木叶片不同时间段稳态荧光变化
对植物快速叶绿素荧光动力学曲线参数进行进一步的分析,可以获得大量关于PSII 原初光化学反应的信息。K 点相对可变荧光(Wk)可以表示PSII供体侧放氧复合体的受伤害程度。由图4 可以看出,随着盐处理时间的增加,Wk值也在逐渐升高,到第14 d 有显著升高,上升幅度为14.8%,表明葡萄叶片PSII 供体侧受到了盐胁迫的伤害。而单位面积有活性反应中心数量(RC/CSm)及捕获的电子传递至QA以后的概率(ΨEo)在处理至第7 d 时较对照都有所降低,下降的幅度分别为7.5%、14.3%,第14 d出现显著下降,下降幅度分别为45.4%、30.3%(图5、6)。这都表明盐胁迫伤害了PSII 光合电子传递链。
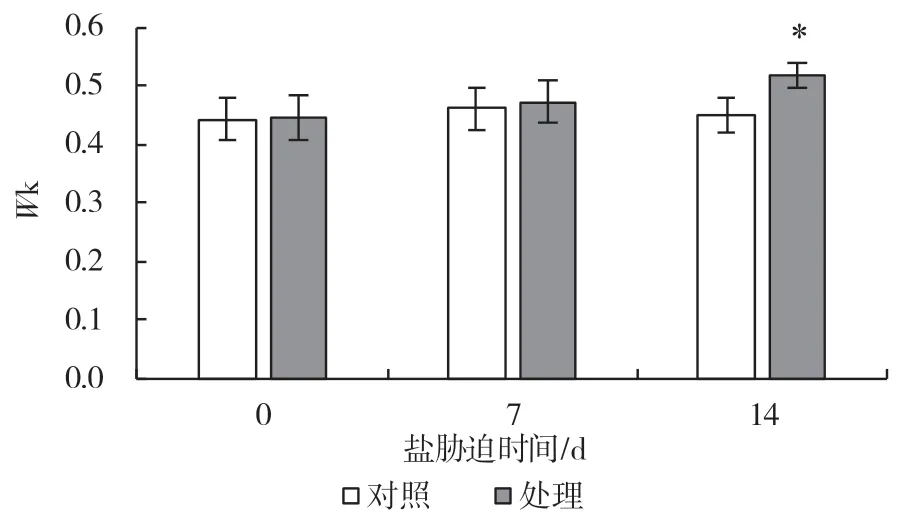
图4 盐胁迫对葡萄砧木叶片K 点相对可变荧光的影响

图5 盐胁迫对单位面积活性反应中心数量的影响

图6 盐胁迫对捕获的电子传递至QA以后的概率的影响
2.3 盐胁迫对光系统能量分配的影响
盐胁迫改变了PSII 和PSI 的能量分配。由图7可知,随着盐处理时间的延长,PSII 可调节的热耗散Y(NPQ)逐渐升高,第14 d 与对照相比差异显著,上升了12.3%;而不可调节的热耗散Y(NO)虽有所上升但差异不显著,表明PSII 活性受到了抑制。与此同时,PSI 氧化态P700 即Y(ND)逐渐降低,第14 d 与对照相比差异显著,下降了12.3%,PSI 供体侧受抑制程度明显增加,而还原态P700+即Y(NA)逐渐升高,第7 d 即出现显著升高,升高了23.5%,第14 d 相比对照升高了127.3%,说明PSI 受体侧受抑制程度明显增加,进一步表明盐胁迫导致了PSI 光抑制的发生。
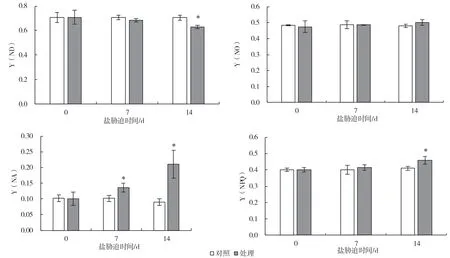
图7 盐胁迫对葡萄砧木叶片PSII 和PSI 光能利用效率的影响
利用PSII 荧光产量进一步分析2 个光系统之间的激发能分配,从图8 可以看到,随着盐胁迫处理时间的延长(β/α-1)的值逐渐增大,与对照处理相比,第7、14 d 处理后分别增加了8.4%和75.8%,说明盐胁迫导致了葡萄叶片PSII 和PSI 间激发能分配不平衡性的增大。
2.4 盐胁迫对环式电子传递的影响
通过检测作用光关闭后叶绿素荧光瞬时上升可以检测NDH 途径的环式电子传递能力的大小,从图9 可以看出,随着盐处理时间的延长,作用光关闭后的荧光上升幅度及斜率均逐渐降低,处理第7 d 就出现明显下降,表明长时间的盐处理抑制了依赖NDH 途径的CEF 活性。

图8 盐胁迫对葡萄砧木叶片PSII 和PSI 间激发能分配不平衡性的影响
P700+再还原动力曲线的半衰期能反映环式电子传递的速率[9],如图10 所示,随着盐处理的进行,第7 dT1/2值出现了显著升高,比对照升高了11.2%,第14 d 比对照升高了30.7%,结果进一步表明长时间盐处理抑制了葡萄CEF 的活性。
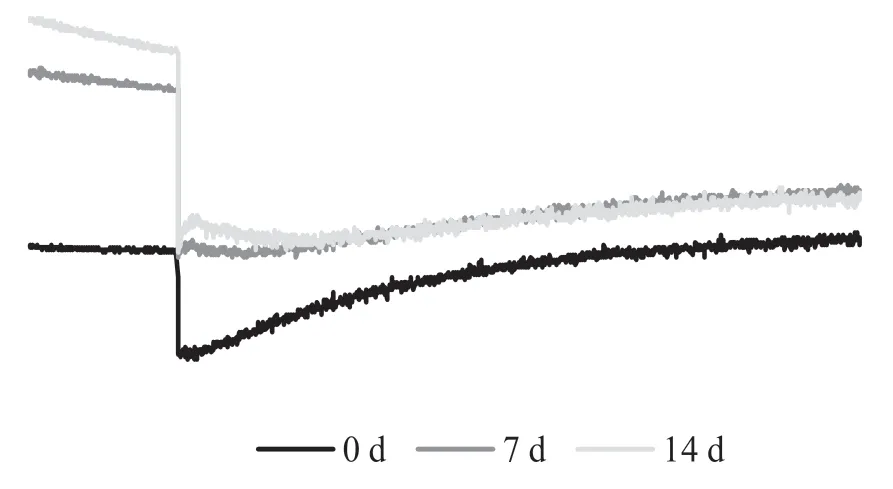
图9 盐胁迫作用光关闭后叶绿素荧光瞬时曲线
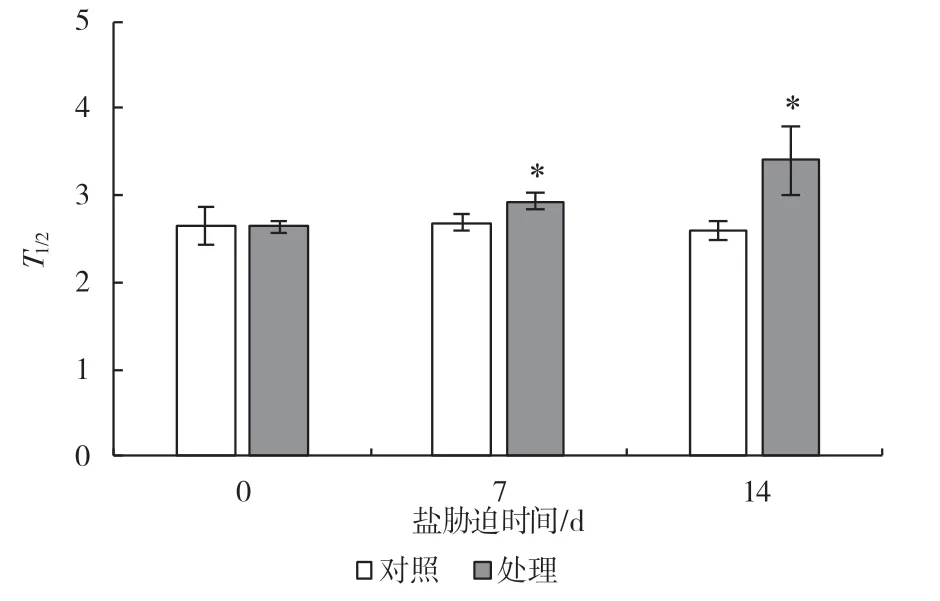
图10 盐胁迫对P700+再还原动力曲线半衰期的影响
3 讨论与结论
叶绿体是植物进行光合作用的细胞器,对叶片的叶绿素荧光参数进行分析能够反映胁迫条件下植物光合器官的伤害程度。光合机构由PSI 和PSII组成,当植物吸收的光能超过所能利用的光能时,过剩光能便会导致光合机构的光抑制,甚至造成光破坏[10]。研究表明氯化钠盐胁迫导致叶绿素发生降解[11],引起植物光系统活性发生抑制[4,10,12]。本研究中,随着氯化钠盐处理时间的增加,叶片最大光化学效率及最大光氧化P700 均逐渐降低,且最大光化学效率发生降低时间早于最大光氧化P700,表明盐胁迫抑制了葡萄叶片光系统活性,其中PSI 相比PSII 更容易受到盐胁迫的抑制,这与颜坤等[13]在金银花上的研究结果一致。
光合作用的高效率进行依赖于PSII 和PSI 的协调运转,从而确保光合色素吸收的光能在2 个光系统间的均衡分配[10]。盐胁迫下叶片2 个光系统间激发能分配失衡,叶片不能通过状态转换有效地将吸收的光能从PSII 传递给PSI,导致了葡萄叶片K 点相对可变荧光逐渐升高,而用于电子传递量子产额、单位面积有活性反应中心的数量均表现出下降趋势,表明盐胁迫抑制了葡萄叶片PSII 供体侧和受体侧导致PSII 电子传递受到抑制。随着处理时间的延长,叶片可调节的能量耗散Y(NPQ)逐渐升高,一方面表明其接受的光强过剩,另一方面说明叶片启动了与高能态淬灭有关的叶黄素循环过程并不能耗散掉所有的光能,随着盐胁迫时间的延长,过剩光能的比例越来越高,非调节性能量耗散Y(NO)逐渐升高,叶片发生严重光抑制[14]。
研究表明围绕PSI 的环式电子传递在逆境胁迫下对稳定PSI 活性起到了重要的作用,进而有助于PSII 的保护和修复[15]。植物体内CEF 主要起到了2方面作用:通过NPQ 进行热耗散[16-17]以及提供ATP进行CO2固定[18]。作用光关闭后荧光瞬时上升能够反映依赖NDH途径的CEF的活性[19],研究发现NDH途径在光保护过程中起重要作用[20],能够减轻逆境胁迫下叶绿体ROS 的积累及基质的过度还原[21]。随着盐胁迫处理时间的延长,葡萄叶片叶绿素荧光瞬时上升幅度和斜率均降低,表明长时间的盐处理抑制了NDH 途径。P700+再还原速率可以用来反映CEF 活性[22]。本研究中,盐胁迫下伴随光抑制的加剧,P700+再还原动力曲线的半衰期逐渐增大,这些结果说明长期盐胁迫下,CEF 活性的抑制可能是导致能够通过对叶绿体基质的过度还原,增加ROS 的累积,加剧光抑制程度。长期盐胁迫导致葡萄砧木2 个光系统激发能分配严重偏离平衡状态,抑制了光系统活性,更多的能量被用来进行热耗散,而围绕PSI 的环式电子传递活性的抑制则进一步加剧了葡萄叶片光系统光抑制程度。
